
Pioglitazone Ameliorates Hippocampal Neurodegeneration, Disturbances in Glucose Metabolism and AKT/mTOR Signaling Pathways in Pentyelenetetrazole-Kindled Mice

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
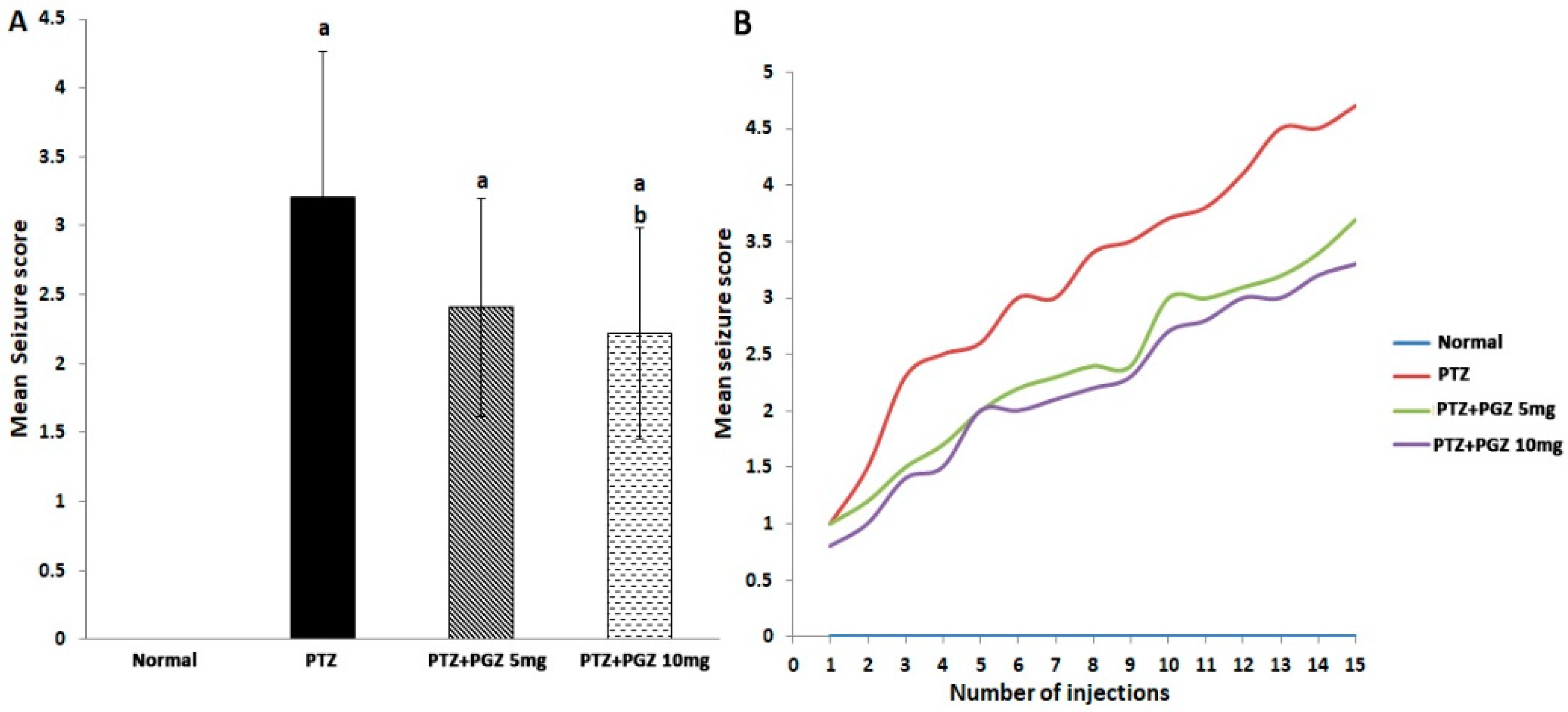
2.1. Pioglitazone Reduced Mean Seizure Score
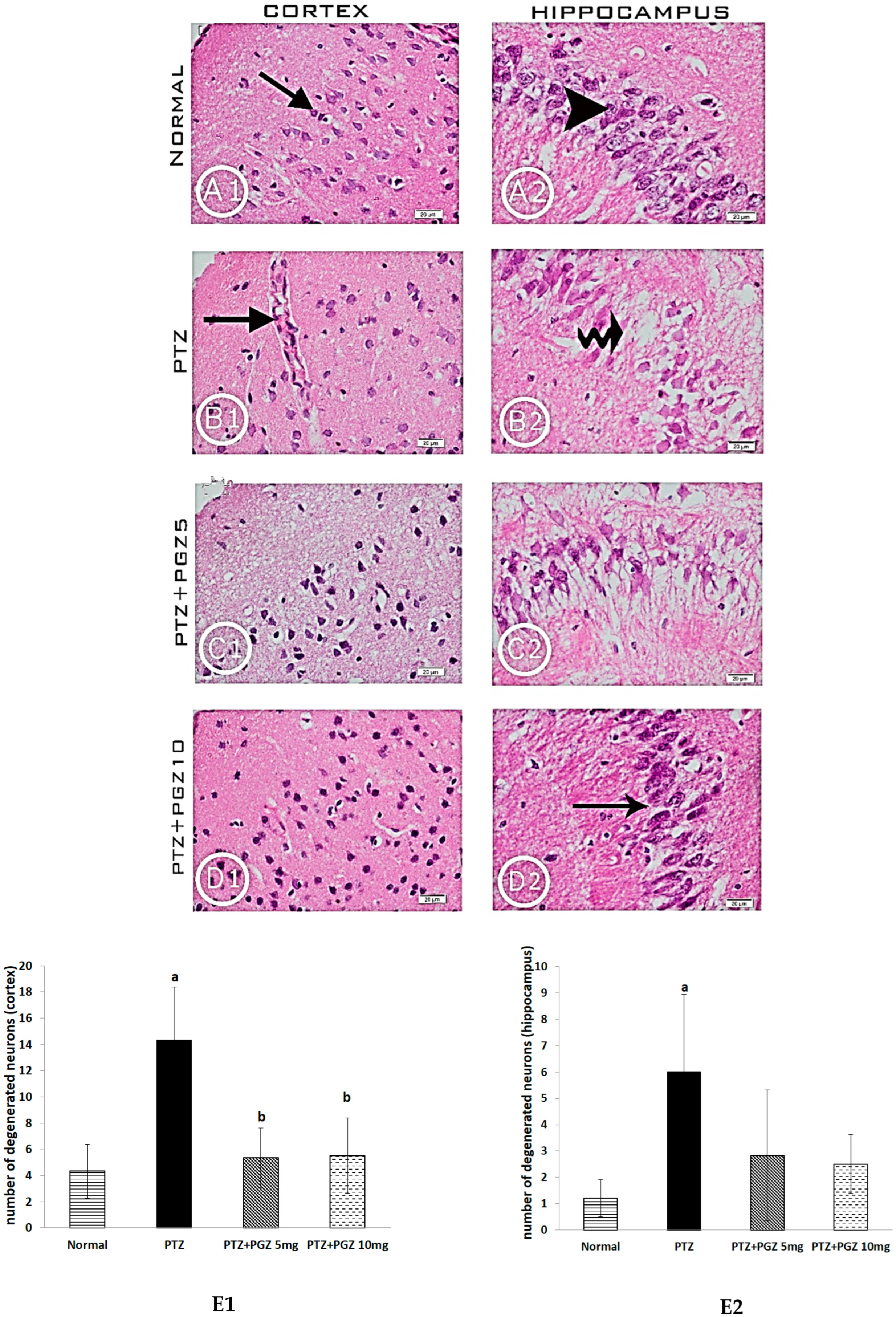
2.2. Pioglitazone Improved the Histological Features in the Hippocampus and Cortex of PTZ Epileptic Mice
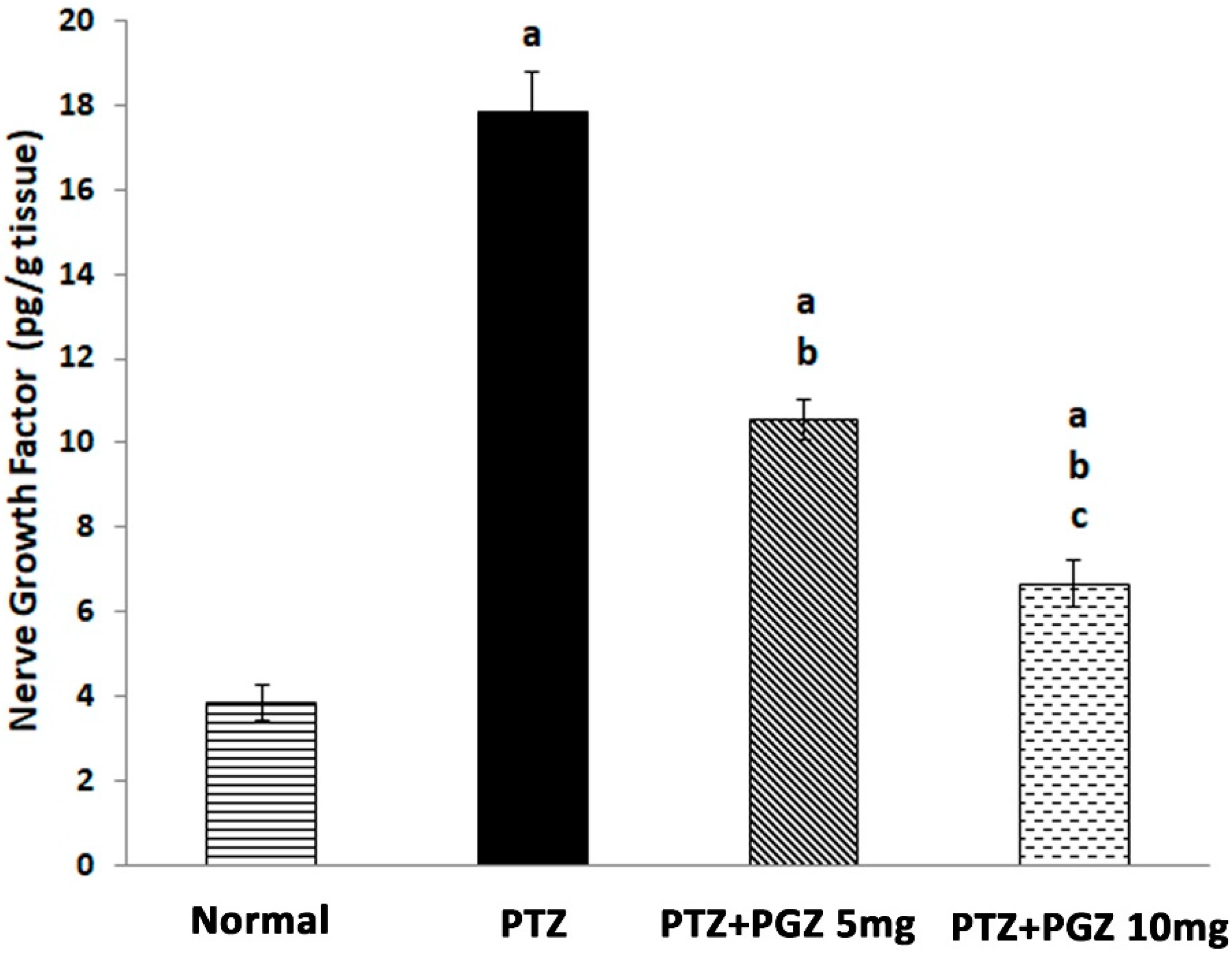
2.3. Pioglitazone Reduced NGF Levels
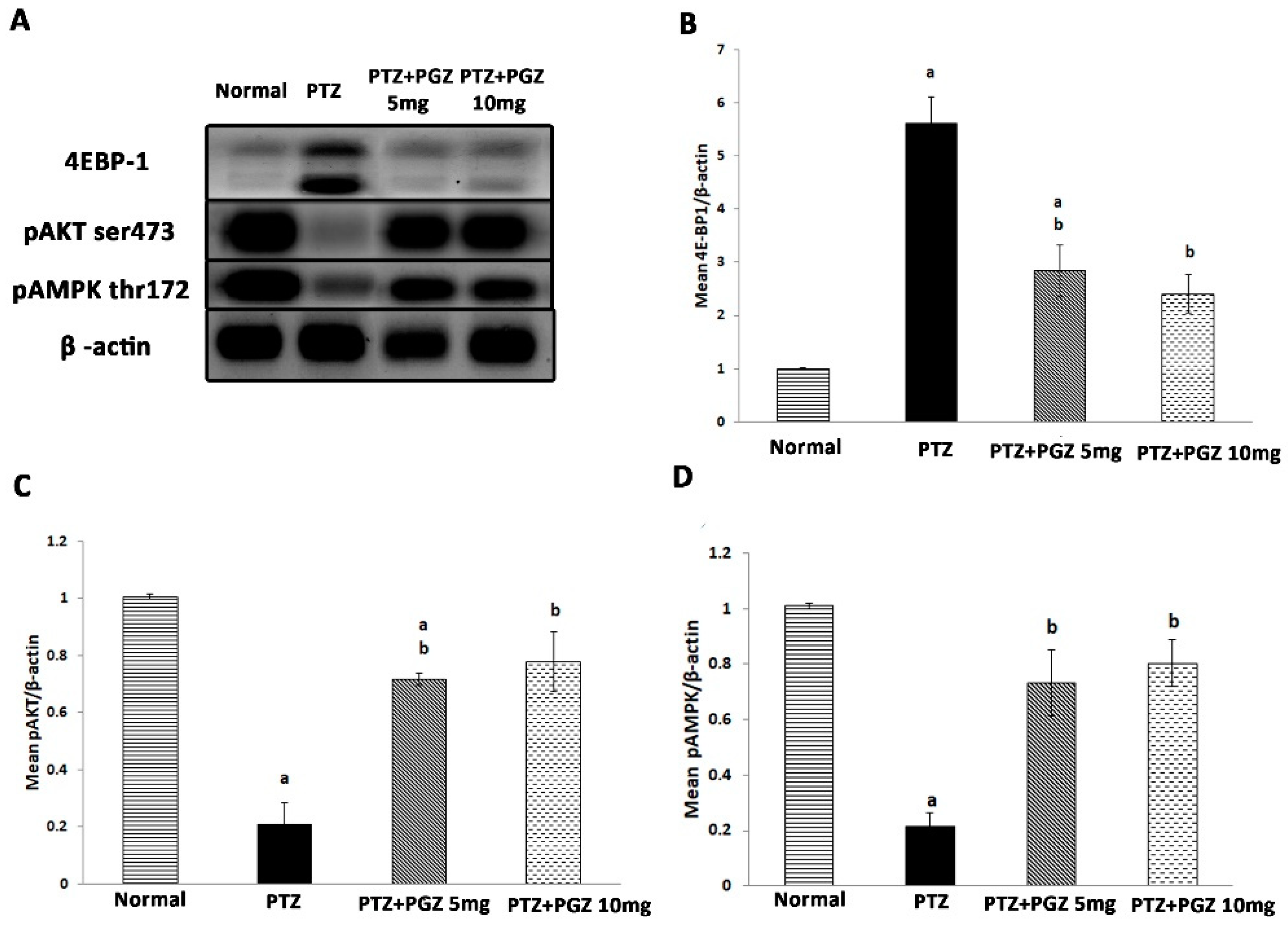
2.4. Pioglitazone Upregulated AKT/AMPK and Downregulated mTOR Signaling Pathways
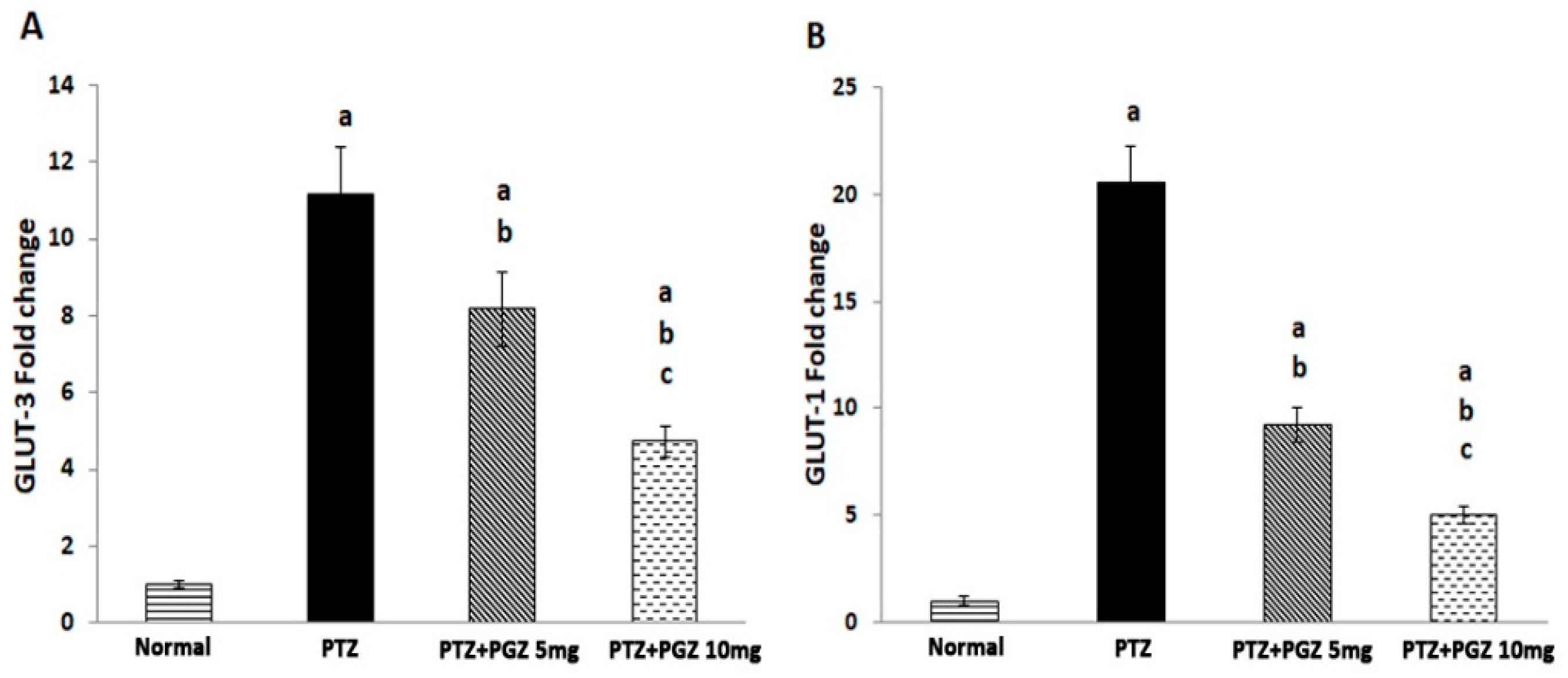
2.5. Pioglitazone Reduced GLUT 1 and GLUT 3 Genes Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals and Drugs
4.3. Induction of Epilepsy in Mice
4.4. Experimental Design
4.5. Assessment of Seizure Activity in PTZ-Kindled Mice
4.6. Tissue Preparation
4.7. Histopathological Examination
4.8. ELISA Analysis
4.9. Evaluation of GLUT-1 and GLUT-3 Genes Expression in Hippocampus by Real-Time PCR
4.10. Western Blotting Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, C.-X.; Bian, M.; Gong, G.-H. Current Research on Antiepileptic Compounds. Molecules 2015, 20, 20741–20776. [Google Scholar] [CrossRef] [PubMed]
- Kwan, P.; Brodie, M.J. Early Identification of Refractory Epilepsy. N. Engl. J. Med. 2000, 342, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Ostendorf, A.P.; Wong, M. MTOR Inhibition in Epilepsy: Rationale and Clinical Perspectives. CNS Drugs 2015, 29, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, K.; Kotulska, K.; Jóźwiak, S. Role of MTOR Inhibitors in Epilepsy Treatment. Pharmacol. Rep. 2015, 67, 636–646. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, H.; Yang, J.; Wu, J.; McMahon, J.; Lin, Y.; Cao, Z.; Gruenthal, M.; Huang, Y. Pharmacological Inhibition of the Mammalian Target of Rapamycin Pathway Suppresses Acquired Epilepsy. Neurobiol. Dis. 2010, 40, 193–199. [Google Scholar] [CrossRef]
- Leo, A.; Constanti, A.; Coppola, A.; Citraro, R.; De Sarro, G.; Russo, E. MTOR Signaling in Epilepsy and Epileptogenesis: Preclinical and Clinical Studies. In Molecules to Medicine with mTOR; Maiese, K., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 123–142. [Google Scholar]
- Stafstrom, C.E.; Ockuly, J.C.; Murphree, L.; Valley, M.T.; Roopra, A.; Sutula, T.P. Anticonvulsant And Antiepileptic Actions of 2-Deoxy-Dglucose In Epilepsy Models. Ann. Neurol. 2009, 65, 435–447. [Google Scholar] [CrossRef]
- Lei, J.; Feng, F.; Duan, Y.; Xu, F.; Liu, Z.; Lian, L.; Liang, Q.; Zhang, N.; Wang, F. Intranasal Nerve Growth Factor Attenuating the Seizure Onset via P75R/Caspase Pathway in the Experimental Epilepsy. Brain Res. Bull. 2017, 134, 79–84. [Google Scholar] [CrossRef]
- Adams, B.; Sazgar, M.; Osehobo, P.; Van der Zee, C.E.E.M.; Diamond, J.; Fahnestock, M.; Racine, R.J. Nerve Growth Factor Accelerates Seizure Development, Enhances Mossy Fiber Sprouting, and Attenuates Seizure-Induced Decreases in Neuronal Density in the Kindling Model of Epilepsy. J. Neurosci. 1997, 17, 5288–5296. [Google Scholar] [CrossRef]
- Okada, K.; Yamashita, U.; Tsuji, S. Ameliorative Effect of Pioglitazone on Seizure Responses in Genetically Epilepsy-Susceptible EL Mice. Brain Res. 2006, 1102, 175–178. [Google Scholar] [CrossRef]
- Lee, C.H.; Yi, M.-H.; Chae, D.J.; Zhang, E.; Oh, S.-H.; Kim, D.W. Effect of Pioglitazone on Excitotoxic Neuronal Damage in the Mouse Hippocampus. Biomol. Ther. 2015, 23, 261–267. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, D.M. Anticonvulsant Potential of the Peroxisome Proliferator-Activated Receptor Gamma Agonist Pioglitazone in Pentylenetetrazole-Induced Acute Seizures and Kindling in Mice. Brain Res. 2010, 1351, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Shafaroodi, H.; Moezi, L.; Ghorbani, H.; Zaeri, M.; Hassanpour, S.; Hassanipour, M.; Dehpour, A.R. Sub-Chronic Treatment with Pioglitazone Exerts Anti-Convulsant Effects in Pentylenetetrazole-Induced Seizures of Mice: The Role of Nitric Oxide. Brain Res. Bull. 2012, 87, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Radu, B.M.; Epureanu, F.B.; Radu, M.; Fabene, P.F.; Bertini, G. Nonsteroidal Anti-Inflammatory Drugs in Clinical and Experimental Epilepsy. Epilepsy Res. 2017, 131, 15–27. [Google Scholar] [CrossRef] [PubMed]
- San, Y.-Z.; Liu, Y.; Zhang, Y.; Shi, P.-P.; Zhu, Y.-L. Peroxisome Proliferator-Activated Receptor-γ Agonist Inhibits the Mammalian Target of Rapamycin Signaling Pathway and Has a Protective Effect in a Rat Model of Status Epilepticus. Mol. Med. Rep. 2015, 12, 1877–1883. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Ahmad, F.J.; Pillai, K.K.; Vohora, D. Amiloride Protects against Pentylenetetrazole-Induced Kindling in Mice. Br. J. Pharmacol. 2005, 145, 880–884. [Google Scholar] [CrossRef]
- Singh, T.; Mishra, A.; Goel, R.K. PTZ Kindling Model for Epileptogenesis, Refractory Epilepsy, and Associated Comorbidities: Relevance and Reliability. Metab. Brain Dis. 2021, 36, 1573–1590. [Google Scholar] [CrossRef]
- Shimada, T.; Yamagata, K. Pentylenetetrazole-Induced Kindling Mouse Model. J. Vis. Exp. 2018, 136, 56573. [Google Scholar] [CrossRef]
- Blumcke, I.; Spreafico, R.; Haaker, G.; Coras, R.; Kobow, K.; Bien, C.G.; Pfäfflin, M.; Elger, C.; Widman, G.; Schramm, J.; et al. Histopathological Findings in Brain Tissue Obtained during Epilepsy Surgery. N. Engl. J. Med. 2017, 377, 1648–1656. [Google Scholar] [CrossRef]
- Takahashi, M.; Hayashi, S.; Kakita, A.; Wakabayashi, K.; Fukuda, M.; Kameyama, S.; Tanaka, R.; Takahashi, H.; Nawa, H. Patients with Temporal Lobe Epilepsy Show an Increase in Brain-Derived Neurotrophic Factor Protein and Its Correlation with Neuropeptide Y. Brain Res. 1999, 818, 579–582. [Google Scholar] [CrossRef]
- Wang, Z.-G.; Li, H.; Huang, Y.; Li, R.; Wang, X.-F.; Yu, L.-X.; Guang, X.-Q.; Li, L.; Zhang, H.-Y.; Zhao, Y.-Z.; et al. Nerve Growth Factor-Induced Akt/MTOR Activation Protects the Ischemic Heart via Restoring Autophagic Flux and Attenuating Ubiquitinated Protein Accumulation. Oncotarget 2016, 8, 5400–5413. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.H.; Bordey, A. Convergent and Divergent Mechanisms of Epileptogenesis in mTORopathies. Front. Neuroanat. 2021, 15, 664695. [Google Scholar] [CrossRef] [PubMed]
- Tekgul, H.; Simsek, E.; Erdoğan, M.A.; Yiğittürk, G.; Erbaş, O.; Taşkıran, D. The Potential Effects of Anticonvulsant Drugs on Neuropeptides and Neurotrophins in Pentylenetetrazol Kindled Seizures in the Rat. Int. J. Neurosci. 2020, 130, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yan, H.; Frost, P.; Gera, J.; Lichtenstein, A. Mammalian Target of Rapamycin Inhibitors Activate the AKT Kinase in Multiple Myeloma Cells by Up-Regulating the Insulin-like Growth Factor Receptor/Insulin Receptor Substrate-1/Phosphatidylinositol 3-Kinase Cascade. Mol. Cancer Ther. 2005, 4, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Svejda, B.; Kidd, M.; Kazberouk, A.; Lawrence, B.; Pfragner, R.; Modlin, I.M. Limitations in Small Intestinal Neuroendocrine Tumor Therapy by MTor Kinase Inhibition Reflect Growth Factor-Mediated PI3K Feedback Loop Activation via ERK1/2 and AKT. Cancer 2011, 117, 4141–4154. [Google Scholar] [CrossRef]
- Hahn-Windgassen, A.; Nogueira, V.; Chen, C.-C.; Skeen, J.E.; Sonenberg, N.; Hay, N. Akt Activates the Mammalian Target of Rapamycin by Regulating Cellular ATP Level and AMPK Activity. J. Biol. Chem. 2005, 280, 32081–32089. [Google Scholar] [CrossRef]
- Atef, M.M.; El-Sayed, N.M.; Ahmed, A.A.M.; Mostafa, Y.M. Donepezil Improves Neuropathy through Activation of AMPK Signalling Pathway in Streptozotocin-Induced Diabetic Mice. Biochem. Pharmacol. 2019, 159, 1–10. [Google Scholar] [CrossRef]
- Li, B.B.; Qian, C.; Gameiro, P.A.; Liu, C.-C.; Jiang, T.; Roberts, T.M.; Struhl, K.; Zhao, J.J. Targeted Profiling of RNA Translation Reveals MTOR-4EBP1/2-Independent Translation Regulation of MRNAs Encoding Ribosomal Proteins. Proc. Natl. Acad. Sci. USA 2018, 115, E9325–E9332. [Google Scholar] [CrossRef]
- Woodcock, H.V.; Eley, J.D.; Guillotin, D.; Platé, M.; Nanthakumar, C.B.; Martufi, M.; Peace, S.; Joberty, G.; Poeckel, D.; Good, R.B.; et al. The mTORC1/4E-BP1 Axis Represents a Critical Signaling Node during Fibrogenesis. Nat. Commun. 2019, 10, 6. [Google Scholar] [CrossRef]
- Rozengurt, E.; Soares, H.P.; Sinnet-Smith, J. Suppression of Feedback Loops Mediated by PI3K/MTOR Induces Multiple over-Activation of Compensatory Pathways: An Unintended Consequence Leading to Drug Resistance. Mol. Cancer Ther. 2014, 13, 2477–2488. [Google Scholar] [CrossRef]
- Gronlund, K.M.; Gerhart, D.Z.; Leino, R.L.; McCall, A.L.; Drewes, L.R. Chronic Seizures Increase Glucose Transporter Abundance in Rat Brain. J. Neuropathol. Exp. Neurol. 1996, 55, 832–840. [Google Scholar] [CrossRef] [Green Version]
- Nehlig, A.; Rudolf, G.; Leroy, C.; Rigoulot, M.-A.; Simpson, I.A.; Vannucci, S.J. Pentylenetetrazol-Induced Status Epilepticus up-Regulates the Expression of Glucose Transporter MRNAs but Not Proteins in the Immature Rat Brain. Brain Res. 2006, 1082, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Ke, R.; Xu, Q.; Li, C.; Luo, L.; Huang, D. Mechanisms of AMPK in the Maintenance of ATP Balance during Energy Metabolism. Cell Biol. Int. 2018, 42, 384–392. [Google Scholar] [CrossRef]
- Assaf, N.; El-Shamarka, M.E.; Salem, N.A.; Khadrawy, Y.A.; El Sayed, N.S. Neuroprotective Effect of PPAR Alpha and Gamma Agonists in a Mouse Model of Amyloidogenesis through Modulation of the Wnt/Beta Catenin Pathway via Targeting Alpha- and Beta-Secretases. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 97, 109793. [Google Scholar] [CrossRef] [PubMed]
- Barbiero, J.K.; Santiago, R.M.; Persike, D.S.; da Silva Fernandes, M.J.; Tonin, F.S.; da Cunha, C.; Lucio Boschen, S.; Lima, M.M.S.; Vital, M.A.B.F. Neuroprotective Effects of Peroxisome Proliferator-Activated Receptor Alpha and Gamma Agonists in Model of Parkinsonism Induced by Intranigral 1-Methyl-4-Phenyl-1,2,3,6-Tetrahyropyridine. Behav. Brain Res. 2014, 274, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.A.; Moghimi, A.; Roohbakhsh, A. Anticonvulsant and Ameliorative Effects of Pioglitazone on Cognitive Deficits, Inflammation and Apoptosis in the Hippocampus of Rat Pups Exposed to Febrile Seizure. Iran. J. Basic Med. Sci. 2019, 22, 267–276. [Google Scholar]
- Citraro, R.; Leo, A.; Constanti, A.; Russo, E.; De Sarro, G. MTOR Pathway Inhibition as a New Therapeutic Strategy in Epilepsy and Epileptogenesis. Pharmacol. Res. 2016, 107, 333–343. [Google Scholar] [CrossRef]
- Goldstein, H.E.; Hauptman, J.S. The Putative Role of MTOR Inhibitors in Non-Tuberous Sclerosis Complex-Related Epilepsy. Front. Neurol. 2021, 12, 639319. [Google Scholar] [CrossRef]
- Galea, E.; Feinstein, D.; Lacombe, P. Pioglitazone Does Not Increase Cerebral Glucose Utilisation in a Murine Model of Alzheimer’s Disease and Decreases It in Wild-Type Mice. Diabetologia 2006, 49, 2153–2161. [Google Scholar] [CrossRef]
- Sefil, F.; Bagirici, F.; Acar, M.D.; Marangoz, C. Influence of Carbenoxolone on the Anticonvulsant Efficacy of Phenytoin in Pentylenetetrazole Kindled Rats. Acta Neurobiol. Exp. 2012, 72, 177–184. [Google Scholar]
- Moezi, L.; Hassanipour, M.; Zaeri, M.; Ghorbani, H.; Shafaroodi, H. The Influence of Ovariectomy on Anti-Convulsant Effect of Pioglitazone in Mice. Pathophysiology 2015, 22, 159–163. [Google Scholar] [CrossRef]
- El-Azab, M.F.; Moustafa, Y.M. Influence of Calcium Channel Blockers on Anticonvulsant and Antinociceptive Activities of Valproic Acid in Pentylenetetrazole-Kindled Mice. Pharmacol. Rep. 2012, 64, 305–314. [Google Scholar] [CrossRef]
- Carson, F.L.; Hladik, C.; Cappellano, C.H. Histotechnology: A Self-Instructional Text; American Society for Clinical Pathology: Chicago, IL, USA, 2015. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Annealing Temperature |
|---|---|---|
| GLUT-1 | Forward: CCAGCTGGGAATCGTCGTT | 62 °C |
| Reverse: CAAGTCTGCATTGCCCATGAT | ||
| GLUT-3 | Forward: CTCTTCAGGTCACCCAACTACGT | 55 °C |
| Reverse: CCGCGTCCTTGAAGATTCC | ||
| β -actin | Forward: ACGGCCAGGTCATCACTATTG | 56 °C |
| Reverse: CAAGAAGGAAGGCTGGAAAAGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Megiri, N.; Mostafa, Y.M.; Ahmed, A.; Mehanna, E.T.; El-Azab, M.F.; Alshehri, F.; Alahdal, H.; El-Sayed, N.M. Pioglitazone Ameliorates Hippocampal Neurodegeneration, Disturbances in Glucose Metabolism and AKT/mTOR Signaling Pathways in Pentyelenetetrazole-Kindled Mice. Pharmaceuticals 2022, 15, 1113. https://doi.org/10.3390/ph15091113
El-Megiri N, Mostafa YM, Ahmed A, Mehanna ET, El-Azab MF, Alshehri F, Alahdal H, El-Sayed NM. Pioglitazone Ameliorates Hippocampal Neurodegeneration, Disturbances in Glucose Metabolism and AKT/mTOR Signaling Pathways in Pentyelenetetrazole-Kindled Mice. Pharmaceuticals. 2022; 15(9):1113. https://doi.org/10.3390/ph15091113
Chicago/Turabian StyleEl-Megiri, Nada, Yasser M. Mostafa, Amal Ahmed, Eman T. Mehanna, Mona F. El-Azab, Fatma Alshehri, Hadil Alahdal, and Norhan M. El-Sayed. 2022. "Pioglitazone Ameliorates Hippocampal Neurodegeneration, Disturbances in Glucose Metabolism and AKT/mTOR Signaling Pathways in Pentyelenetetrazole-Kindled Mice" Pharmaceuticals 15, no. 9: 1113. https://doi.org/10.3390/ph15091113
APA StyleEl-Megiri, N., Mostafa, Y. M., Ahmed, A., Mehanna, E. T., El-Azab, M. F., Alshehri, F., Alahdal, H., & El-Sayed, N. M. (2022). Pioglitazone Ameliorates Hippocampal Neurodegeneration, Disturbances in Glucose Metabolism and AKT/mTOR Signaling Pathways in Pentyelenetetrazole-Kindled Mice. Pharmaceuticals, 15(9), 1113. https://doi.org/10.3390/ph15091113