
ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets

Abstract
:
1. Introduction
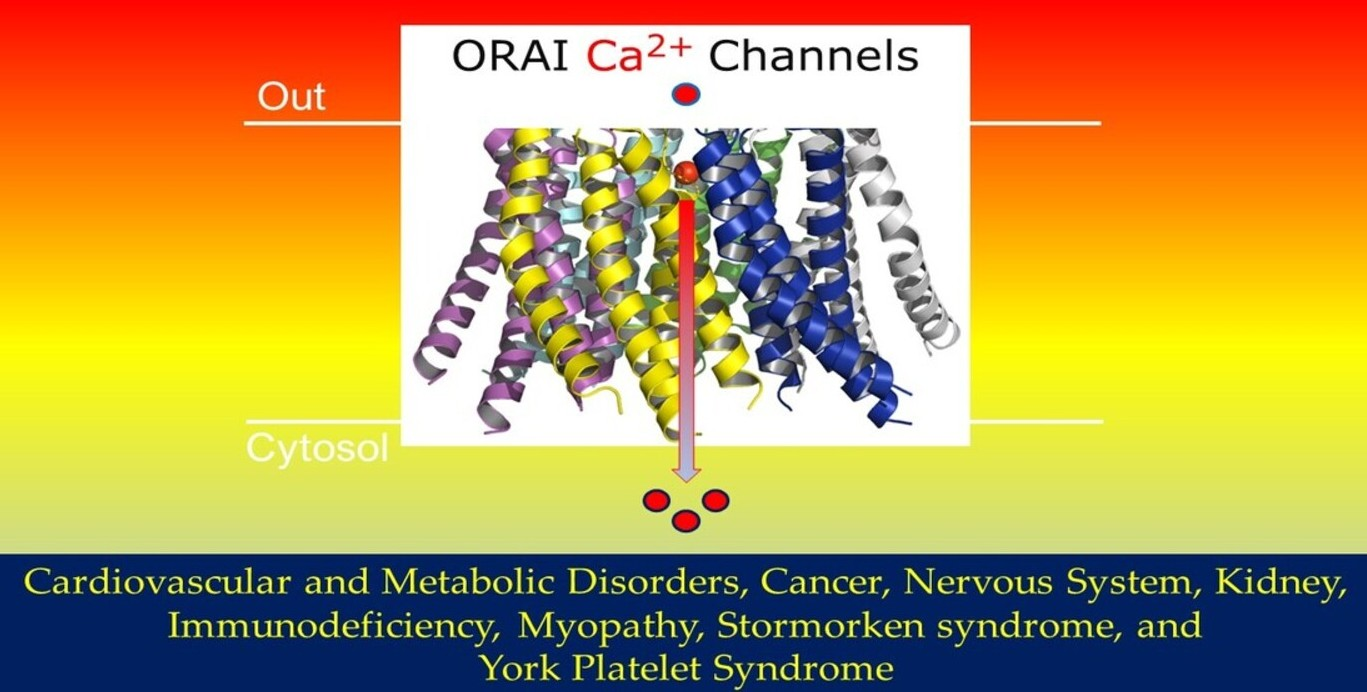
1.1. Membrane Topology Structure of the ORAI/STIM Channel Complexes
1.2. Activation and Regulation of ORAI/STIM Channel Complexes
2. ORAI/STIM Channel Complexes in Human Diseases
2.1. Immunodeficiency
2.2. Myopathy
2.3. Tubular Aggregate Myopathy
2.4. Stormorken Syndrome
2.5. York Platelet Syndrome
2.6. Cardiovascular and Metabolic Disorders
2.7. Cancers
2.8. Nervous System
2.9. Kidney
3. Pharmacology of Store-Operated Calcium Entry Channels
3.1. Activators of ORAI/STIM Channel Complexes
3.2. Thapsigargin
3.3. 2T and 3G
3.4. IA65
3.5. Inhibitors of ORAI/STIM Channel Complexes
3.6. Pyrazole Compounds: GSK-7975A and GSK-5503A
3.7. Compound 5J-4
3.8. 2-APB and Its Analogues DPB162-AE and DPB163-AE
3.9. Aspirin Metabolite Salicylate and Other NSAIDs
3.10. SKF-96365
3.11. Mibefradil
3.12. 4-Choloro-3-ethylphenol (4-CEP) and Its Analogues 4-Choloro-m-cresol (4-CmC) and 4-Chlorophenol (4-CIP)
3.13. Lanthanides
3.14. Bistrifluoromethyl-Pyrazole Derivative (BTP-2) or Pyrazole Derivatives
3.15. Synta-66
3.16. RO2959
3.17. CM4620, CM2489, CM3457, CM128 (CalciMedica Series)
3.18. AnCoA4
3.19. ML-9
3.20. JPIII
3.21. Pyrtriazole Compound 39
3.22. RP3128 and RP4010
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Putney, J.W. A Model for Receptor-Regulated Calcium Entry. Cell Calcium 1986, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Chen, G.-L.; Garcia-Vaz, E.; Bhandari, S.; Daskoulidou, N.; Berglund, L.M.; Jiang, H.; Hallett, T.; Zhou, L.-P.; Huang, L.; et al. ORAI Channels Are Critical for Receptor-Mediated Endocytosis of Albumin. Nat. Commun. 2017, 8, 1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Merritt, R.; Fu, L.; Pan, Z. Targeting Calcium Signaling in Cancer Therapy. Acta Pharm. Sin. B 2017, 7, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Rubaiy, H.N. A Short Guide to Electrophysiology and Ion Channels. J. Pharm. Pharm. Sci. 2017, 20, 48–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mignen, O.; Thompson, J.L.; Shuttleworth, T.J. Orai1 Subunit Stoichiometry of the Mammalian CRAC Channel Pore. J. Physiol. 2008, 586, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Balasuriya, D.; Srivats, S.; Murrell-Lagnado, R.D.; Edwardson, J.M. Atomic Force Microscopy (AFM) Imaging Suggests That Stromal Interaction Molecule 1 (STIM1) Binds to Orai1 with Sixfold Symmetry. FEBS Lett. 2014, 588, 2874–2880. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Pedi, L.; Diver, M.M.; Long, S.B. Crystal Structure of the Calcium Release-Activated Calcium Channel Orai. Science 2012, 338, 1308–1313. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.L.; Shuttleworth, T.J. How Many Orai’s Does It Take to Make a CRAC Channel? Sci. Rep. 2013, 3, 1961. [Google Scholar] [CrossRef] [Green Version]
- Yen, M.; Lewis, R.S. Numbers Count: How STIM and Orai Stoichiometry Affect Store-Operated Calcium Entry. Cell Calcium 2019, 79, 35–43. [Google Scholar] [CrossRef]
- Cai, X.; Zhou, Y.; Nwokonko, R.M.; Loktionova, N.A.; Wang, X.; Xin, P.; Trebak, M.; Wang, Y.; Gill, D.L. The Orai1 Store-Operated Calcium Channel Functions as a Hexamer * ♦. J. Biol. Chem. 2016, 291, 25764–25775. [Google Scholar] [CrossRef] [Green Version]
- Yen, M.; Lokteva, L.A.; Lewis, R.S. Functional Analysis of Orai1 Concatemers Supports a Hexameric Stoichiometry for the CRAC Channel. Biophys. J. 2016, 111, 1897–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Outhwaite, I.R.; Pedi, L.; Long, S.B. Cryo-EM Structure of the Calcium Release-Activated Calcium Channel Orai in an Open Conformation. eLife 2020, 9, e62772. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Yeung, P.S.-W.; Ing, C.E.; McNally, B.A.; Pomès, R.; Prakriya, M. STIM1 Activates CRAC Channels through Rotation of the Pore Helix to Open a Hydrophobic Gate. Nat. Commun. 2017, 8, 14512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiffner, A.; Maltan, L.; Fahrner, M.; Sallinger, M.; Weiß, S.; Grabmayr, H.; Höglinger, C.; Derler, I. Transmembrane Domain 3 (TM3) Governs Orai1 and Orai3 Pore Opening in an Isoform-Specific Manner. Front. Cell Dev. Biol. 2021, 9, 635705. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubaiy, H.N. The Therapeutic Agents That Target ATP-Sensitive Potassium Channels. Acta Pharm. Zagreb Croat. 2016, 66, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J. Inositol Trisphosphate and Calcium Signalling Mechanisms. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2009, 1793, 933–940. [Google Scholar] [CrossRef] [Green Version]
- Liou, J.; Kim, M.L.; Heo, W.D.; Jones, J.T.; Myers, J.W.; Ferrell, J.E.; Meyer, T. STIM Is a Ca2+ Sensor Essential for Ca2+-Store-Depletion-Triggered Ca2+ Influx. Curr. Biol. CB 2005, 15, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Amcheslavsky, A.; Wood, M.L.; Yeromin, A.V.; Parker, I.; Freites, J.A.; Tobias, D.J.; Cahalan, M.D. Molecular Biophysics of Orai Store-Operated Ca2+ Channels. Biophys. J. 2015, 108, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Fahrner, M.; Grabmayr, H.; Romanin, C. Mechanism of STIM Activation. Curr. Opin. Physiol. 2020, 17, 74–79. [Google Scholar] [CrossRef]
- Park, C.Y.; Hoover, P.J.; Mullins, F.M.; Bachhawat, P.; Covington, E.D.; Raunser, S.; Walz, T.; Garcia, K.C.; Dolmetsch, R.E.; Lewis, R.S. STIM1 Clusters and Activates CRAC Channels via Direct Binding of a Cytosolic Domain to Orai1. Cell 2009, 136, 876–890. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.P.; Zeng, W.; Dorwart, M.R.; Choi, Y.-J.; Worley, P.F.; Muallem, S. SOAR and the Polybasic STIM1 Domains Gate and Regulate the Orai Channels. Nat. Cell Biol. 2009, 11, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shawer, H.; Norman, K.; Cheng, C.W.; Foster, R.; Beech, D.J.; Bailey, M.A. ORAI1 Ca2+ Channel as a Therapeutic Target in Pathological Vascular Remodelling. Front. Cell Dev. Biol. 2021, 9, 653812. [Google Scholar] [CrossRef] [PubMed]
- Prakriya, M.; Lewis, R.S. Store-Operated Calcium Channels. Physiol. Rev. 2015, 95, 1383–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubaiy, H.N. Treasure Troves of Pharmacological Tools to Study Transient Receptor Potential Canonical 1/4/5 Channels. Br. J. Pharmacol. 2019, 176, 832–846. [Google Scholar] [CrossRef] [PubMed]
- Rubaiy, H.N.; Ludlow, M.J.; Bon, R.S.; Beech, D.J. Pico145 - Powerful New Tool for TRPC1/4/5 Channels. Channels 2017, 11, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Rubaiy, H.N.; Ludlow, M.J.; Siems, K.; Norman, K.; Foster, R.; Wolf, D.; Beutler, J.A.; Beech, D.J. Tonantzitlolone Is a Nanomolar Potency Activator of Transient Receptor Potential Canonical 1/4/5 Channels. Br. J. Pharmacol. 2018, 175, 3361–3368. [Google Scholar] [CrossRef] [Green Version]
- Rubaiy, H.N.; Ludlow, M.J.; Henrot, M.; Gaunt, H.J.; Miteva, K.; Cheung, S.Y.; Tanahashi, Y.; Hamzah, N.; Musialowski, K.E.; Blythe, N.M.; et al. Picomolar, Selective, and Subtype-Specific Small-Molecule Inhibition of TRPC1/4/5 Channels. J. Biol. Chem. 2017, 292, 8158–8173. [Google Scholar] [CrossRef] [Green Version]
- Rubaiy, H.N.; Seitz, T.; Hahn, S.; Choidas, A.; Habenberger, P.; Klebl, B.; Dinkel, K.; Nussbaumer, P.; Waldmann, H.; Christmann, M.; et al. Identification of an (−)-englerin A Analogue, Which Antagonizes (−)-englerin A at TRPC1/4/5 Channels. Br. J. Pharmacol. 2018, 175, 830–839. [Google Scholar] [CrossRef]
- Zeng, H.; Sanes, J.R. Neuronal Cell-Type Classification: Challenges, Opportunities and the Path Forward. Nat. Rev. Neurosci. 2017, 18, 530–546. [Google Scholar] [CrossRef]
- Navarro-Borelly, L.; Somasundaram, A.; Yamashita, M.; Ren, D.; Miller, R.J.; Prakriya, M. STIM1–Orai1 Interactions and Orai1 Conformational Changes Revealed by Live-Cell FRET Microscopy. J. Physiol. 2008, 586, 5383–5401. [Google Scholar] [CrossRef] [PubMed]
- Feske, S.; Gwack, Y.; Prakriya, M.; Srikanth, S.; Puppel, S.-H.; Tanasa, B.; Hogan, P.G.; Lewis, R.S.; Daly, M.; Rao, A. A Mutation in Orai1 Causes Immune Deficiency by Abrogating CRAC Channel Function. Nature 2006, 441, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Feske, S. CRAC Channelopathies. Pflugers Arch. 2010, 460, 417–435. [Google Scholar] [CrossRef] [PubMed]
- McCarl, C.-A.; Picard, C.; Khalil, S.; Kawasaki, T.; Röther, J.; Papolos, A.; Kutok, J.; Hivroz, C.; LeDeist, F.; Plogmann, K.; et al. ORAI1 Deficiency and Lack of Store-Operated Ca2+ Entry Cause Immunodeficiency, Myopathy and Ectodermal Dysplasia. J. Allergy Clin. Immunol. 2009, 124, 1311–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partiseti, M.; Le Deist, F.; Hivroz, C.; Fischer, A.; Korn, H.; Choquet, D. The Calcium Current Activated by T Cell Receptor and Store Depletion in Human Lymphocytes Is Absent in a Primary Immunodeficiency. J. Biol. Chem. 1994, 269, 32327–32335. [Google Scholar] [CrossRef] [PubMed]
- Böhm, J.; Laporte, J. Gain-of-Function Mutations in STIM1 and ORAI1 Causing Tubular Aggregate Myopathy and Stormorken Syndrome. Cell Calcium 2018, 76, 1–9. [Google Scholar] [CrossRef]
- Böhm, J.; Bulla, M.; Urquhart, J.E.; Malfatti, E.; Williams, S.G.; O’Sullivan, J.; Szlauer, A.; Koch, C.; Baranello, G.; Mora, M.; et al. ORAI1 Mutations with Distinct Channel Gating Defects in Tubular Aggregate Myopathy. Hum. Mutat. 2017, 38, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Okuma, H.; Saito, F.; Mitsui, J.; Hara, Y.; Hatanaka, Y.; Ikeda, M.; Shimizu, T.; Matsumura, K.; Shimizu, J.; Tsuji, S.; et al. Tubular Aggregate Myopathy Caused by a Novel Mutation in the Cytoplasmic Domain of STIM1. Neurol. Genet. 2016, 2, e50. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Carreras-Sureda, A.; Didier, M.; Henry, C.; Frieden, M.; Demaurex, N. The TAM-Associated STIM1I484R Mutation Increases ORAI1 Channel Function Due to a Reduced STIM1 Inactivation Break and an Absence of Microtubule Trapping. Cell Calcium 2022, 105, 102615. [Google Scholar] [CrossRef]
- Endo, Y.; Noguchi, S.; Hara, Y.; Hayashi, Y.K.; Motomura, K.; Miyatake, S.; Murakami, N.; Tanaka, S.; Yamashita, S.; Kizu, R.; et al. Dominant Mutations in ORAI1 Cause Tubular Aggregate Myopathy with Hypocalcemia via Constitutive Activation of Store-Operated Ca2+ Channels. Hum. Mol. Genet. 2015, 24, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Bulla, M.; Gyimesi, G.; Kim, J.H.; Bhardwaj, R.; Hediger, M.A.; Frieden, M.; Demaurex, N. ORAI1 Channel Gating and Selectivity Is Differentially Altered by Natural Mutations in the First or Third Transmembrane Domain. J. Physiol. 2019, 597, 561–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacruz, R.S.; Feske, S. Diseases Caused by Mutations in ORAI1 and STIM1. Ann. N. Y. Acad. Sci. 2015, 1356, 45–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, G.; Bruechle, N.O.; Singh, A.R.; Knopp, C.; Jedraszak, G.; Elbracht, M.; Brémond-Gignac, D.; Hartmann, K.; Sevestre, H.; Deutz, P.; et al. Gain-of-Function Mutation in STIM1 (P.R304W) Is Associated with Stormorken Syndrome. Hum. Mutat. 2014, 35, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Nesin, V.; Wiley, G.; Kousi, M.; Ong, E.-C.; Lehmann, T.; Nicholl, D.J.; Suri, M.; Shahrizaila, N.; Katsanis, N.; Gaffney, P.M.; et al. Activating Mutations in STIM1 and ORAI1 Cause Overlapping Syndromes of Tubular Myopathy and Congenital Miosis. Proc. Natl. Acad. Sci. USA 2014, 111, 4197–4202. [Google Scholar] [CrossRef] [Green Version]
- Markello, T.; Chen, D.; Kwan, J.Y.; Horkayne-Szakaly, I.; Morrison, A.; Simakova, O.; Maric, I.; Lozier, J.; Cullinane, A.R.; Kilo, T.; et al. York Platelet Syndrome Is a CRAC Channelopathy Due to Gain-of-Function Mutations in STIM1. Mol. Genet. Metab. 2015, 114, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Roman, J.; Palmer, M.I.; Palmer, C.A.; Johnson, N.E.; Butterfield, R.J. Myopathy in the York Platelet Syndrome: An Underrecognized Complication. Case Rep. Pathol. 2018, 2018, 5130143. [Google Scholar] [CrossRef]
- Pacheco, J.; Vaca, L. STIM-TRP Pathways and Microdomain Organization: Auxiliary Proteins of the STIM/Orai Complex. In Store-Operated Ca2+ Entry (SOCE) Pathways: Emerging Signaling Concepts in Human (Patho)physiology; Groschner, K., Graier, W.F., Romanin, C., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; pp. 189–210. ISBN 978-3-319-57732-6. [Google Scholar]
- Xia, W.; Li, Y.; Wang, B.; Chen, J.; Wang, X.; Sun, Q.; Sun, F.; Li, Z.; Zhao, Z. Enhanced Store-Operated Calcium Entry in Platelets Is Associated with Peripheral Artery Disease in Type 2 Diabetes. Cell. Physiol. Biochem. 2015, 37, 1945–1955. [Google Scholar] [CrossRef]
- Grosse, J.; Braun, A.; Varga-Szabo, D.; Beyersdorf, N.; Schneider, B.; Zeitlmann, L.; Hanke, P.; Schropp, P.; Mühlstedt, S.; Zorn, C.; et al. An EF Hand Mutation in Stim1 Causes Premature Platelet Activation and Bleeding in Mice. J. Clin. Invest. 2007, 117, 3540–3550. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Boulaftali, Y.; Greene, T.K.; Ouellette, T.D.; Poncz, M.; Feske, S.; Bergmeier, W. Relative Contribution of STIM1 and CalDAG-GEFI to Calcium-Dependent Platelet Activation and Thrombosis. J. Thromb. Haemost. JTH 2011, 9, 2077–2086. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Wang, L.; Fu, T.-M.; Wu, H. Mechanism of TRPM 2 Channel Gating Revealed by Cryo-EM. FEBS J. 2019, 286, 3333–3339. [Google Scholar] [CrossRef] [Green Version]
- Van Assche, T.; Fransen, P.; Guns, P.-J.; Herman, A.G.; Bult, H. Altered Ca2+ Handling of Smooth Muscle Cells in Aorta of Apolipoprotein E-Deficient Mice before Development of Atherosclerotic Lesions. Cell Calcium 2007, 41, 295–302. [Google Scholar] [CrossRef]
- Stolwijk, J.A.; Zhang, X.; Gueguinou, M.; Zhang, W.; Matrougui, K.; Renken, C.; Trebak, M. Calcium Signaling Is Dispensable for Receptor Regulation of Endothelial Barrier Function. J. Biol. Chem. 2016, 291, 22894–22912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.-Y.; Kelsen, B. Influence of Personality and Motivation on Oral Presentation Performance. J. Psycholinguist. Res. 2018, 47, 755–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinelli, A.M.; Trebak, M. Orai Channel-Mediated Ca2+ Signals in Vascular and Airway Smooth Muscle. Am. J. Physiol.-Cell Physiol. 2016, 310, C402–C413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswamee, P.; Pounardjian, T.; Giovannucci, D.R. Arachidonic Acid-Induced Ca2+ Entry and Migration in a Neuroendocrine Cancer Cell Line. Cancer Cell Int. 2018, 18, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.-K.; Lin, Y.-H.; Chang, H.-A.; Lai, Y.-S.; Chen, Y.-C.; Huang, S.-C.; Chou, C.-Y.; Chiu, W.-T. Chemoresistant Ovarian Cancer Enhances Its Migration Abilities by Increasing Store-Operated Ca2+ Entry-Mediated Turnover of Focal Adhesions. J. Biomed. Sci. 2020, 27, 36. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, N.; Pan, H.; Xie, J.; Han, W. Development of Store-Operated Calcium Entry-Targeted Compounds in Cancer. Front. Pharmacol. 2021, 12, 688244. [Google Scholar] [CrossRef] [PubMed]
- Schmid, E.; Stagno, M.J.; Yan, J.; Stournaras, C.; Lang, F.; Fuchs, J.; Seitz, G. Store-Operated Ca(2+) Entry in Rhabdomyosarcoma Cells. Biochem. Biophys. Res. Commun. 2016, 477, 129–136. [Google Scholar] [CrossRef]
- Wang, Y.; He, J.; Jiang, H.; Zhang, Q.; Yang, H.; Xu, X.; Zhang, C.; Xu, C.; Wang, J.; Lu, W. Nicotine Enhances Store-operated Calcium Entry by Upregulating HIF-1α and SOCC Components in Non-small Cell Lung Cancer Cells. Oncol. Rep. 2018, 40, 2097–2104. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wang, H.; Huang, H.; Sun, L.; Dong, S.; Huang, N.; Shi, M.; Bin, J.; Liao, Y.; Liao, W. Elevated Orai1 and STIM1 Expressions Upregulate MACC1 Expression to Promote Tumor Cell Proliferation, Metabolism, Migration, and Invasion in Human Gastric Cancer. Cancer Lett. 2016, 381, 31–40. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Chiu, W.-T.; Chen, Y.-T.; Lin, P.-Y.; Huang, H.-J.; Chou, C.-Y.; Chang, H.-C.; Tang, M.-J.; Shen, M.-R. Calcium Store Sensor Stromal-Interaction Molecule 1-Dependent Signaling Plays an Important Role in Cervical Cancer Growth, Migration, and Angiogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 15225–15230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Sanderson, M.J. Store-Operated Calcium Entry Is Required for Sustained Contraction and Ca2+ Oscillations of Airway Smooth Muscle. J. Physiol. 2017, 595, 3203–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Begum, G.; Pointer, K.; Clark, P.A.; Yang, S.-S.; Lin, S.-H.; Kahle, K.T.; Kuo, J.S.; Sun, D. WNK1-OSR1 Kinase-Mediated Phospho-Activation of Na+-K+-2Cl− Cotransporter Facilitates Glioma Migration. Mol. Cancer 2014, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Schild, A.; Bhardwaj, R.; Wenger, N.; Tscherrig, D.; Kandasamy, P.; Dernič, J.; Baur, R.; Peinelt, C.; Hediger, M.A.; Lochner, M. Synthesis and Pharmacological Characterization of 2-Aminoethyl Diphenylborinate (2-APB) Derivatives for Inhibition of Store-Operated Calcium Entry (SOCE) in MDA-MB-231 Breast Cancer Cells. Int. J. Mol. Sci. 2020, 21, 5604. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Collado, J.; Jardin, I.; López, J.J.; Ronco, V.; Salido, G.M.; Dubois, C.; Prevarskaya, N.; Rosado, J.A. Role of Orai3 in the Pathophysiology of Cancer. Int. J. Mol. Sci. 2021, 22, 11426. [Google Scholar] [CrossRef] [PubMed]
- Ay, B.; Prakash, Y.S.; Pabelick, C.M.; Sieck, G.C. Store-Operated Ca2+ Entry in Porcine Airway Smooth Muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2004, 286, L909–L917. [Google Scholar] [CrossRef] [PubMed]
- Benzerdjeb, N.; Sevestre, H.; Ahidouch, A.; Ouadid-Ahidouch, H. Orai3 Is a Predictive Marker of Metastasis and Survival in Resectable Lung Adenocarcinoma. Oncotarget 2016, 7, 81588–81597. [Google Scholar] [CrossRef] [Green Version]
- Dubois, C.; Vanden Abeele, F.; Lehen’kyi, V.; Gkika, D.; Guarmit, B.; Lepage, G.; Slomianny, C.; Borowiec, A.S.; Bidaux, G.; Benahmed, M.; et al. Remodeling of Channel-Forming ORAI Proteins Determines an Oncogenic Switch in Prostate Cancer. Cancer Cell 2014, 26, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Fiorio Pla, A.; Kondratska, K.; Prevarskaya, N. STIM and ORAI Proteins: Crucial Roles in Hallmarks of Cancer. Am. J. Physiol.-Cell Physiol. 2016, 310, C509–C519. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-F.; Lin, P.-C.; Yeh, Y.-M.; Chen, L.-H.; Shen, M.-R. Store-Operated Ca2+ Entry in Tumor Progression: From Molecular Mechanisms to Clinical Implications. Cancers 2019, 11, 899. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xin, P.; Yoast, R.E.; Emrich, S.M.; Johnson, M.T.; Pathak, T.; Benson, J.C.; Azimi, I.; Gill, D.L.; Monteith, G.R.; et al. Distinct Pharmacological Profiles of ORAI1, ORAI2, and ORAI3 Channels. Cell Calcium 2020, 91, 102281. [Google Scholar] [CrossRef] [PubMed]
- Basile, D.P.; Collett, J.A. Orai1: A New Therapeutic Target for the Acute Kidney Injury-to-Chronic Kidney Disease Transition. Nephron 2022, 146, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Mai, X.; Shang, J.; Liang, S.; Yu, B.; Yuan, J.; Lin, Y.; Luo, R.; Zhang, F.; Liu, Y.; Lv, X.; et al. Blockade of Orai1 Store-Operated Calcium Entry Protects against Renal Fibrosis. J. Am. Soc. Nephrol. JASN 2016, 27, 3063–3078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanwar, J.; Motiani, R.K. Role of SOCE Architects STIM and Orai Proteins in Cell Death. Cell Calcium 2018, 69, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Luik, R.M.; Lewis, R.S. New Insights into the Molecular Mechanisms of Store-Operated Ca2+ Signaling in T Cells. Trends Mol. Med. 2007, 13, 103–107. [Google Scholar] [CrossRef]
- Bird, G.S.J.; Putney, J.W. Capacitative Calcium Entry Supports Calcium Oscillations in Human Embryonic Kidney Cells. J. Physiol. 2005, 562, 697–706. [Google Scholar] [CrossRef]
- Bird, G.S.; DeHaven, W.I.; Smyth, J.T.; Putney Jr, J.W. Methods for Studying Store-Operated Calcium Entry. Methods 2008, 46, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Jairaman, A.; Prakriya, M. Molecular Pharmacology of Store-Operated CRAC Channels. Channels 2013, 7, 402–414. [Google Scholar] [CrossRef] [Green Version]
- Thastrup, O.; Cullen, P.J.; Drøbak, B.K.; Hanley, M.R.; Dawson, A.P. Thapsigargin, a Tumor Promoter, Discharges Intracellular Ca2+ Stores by Specific Inhibition of the Endoplasmic Reticulum Ca2(+)-ATPase. Proc. Natl. Acad. Sci. USA 1990, 87, 2466–2470. [Google Scholar] [CrossRef] [Green Version]
- Sagara, Y.; Inesi, G. Inhibition of the Sarcoplasmic Reticulum Ca2+ Transport ATPase by Thapsigargin at Subnanomolar Concentrations. J. Biol. Chem. 1991, 266, 13503–13506. [Google Scholar] [CrossRef]
- Riva, B.; Griglio, A.; Serafini, M.; Cordero-Sanchez, C.; Aprile, S.; Di Paola, R.; Gugliandolo, E.; Alansary, D.; Biocotino, I.; Lim, D.; et al. Pyrtriazoles, a Novel Class of Store-Operated Calcium Entry Modulators: Discovery, Biological Profiling and In Vivo Proof-of-Concept Efficacy in Acute Pancreatitis. J. Med. Chem. 2018, 61, 9756–9783. [Google Scholar] [CrossRef] [PubMed]
- Azimi, I.; Stevenson, R.J.; Zhang, X.; Meizoso-Huesca, A.; Xin, P.; Johnson, M.; Flanagan, J.U.; Chalmers, S.B.; Yoast, R.E.; Kapure, J.S.; et al. A New Selective Pharmacological Enhancer of the Orai1 Ca2+ Channel Reveals Roles for Orai1 in Smooth and Skeletal Muscle Functions. ACS Pharmacol. Transl. Sci. 2020, 3, 135–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodeify, R.; Yu, F.; Courjaret, R.; Nader, N.; Dib, M.; Sun, L.; Adap, E.; Hubrack, S.; Machaca, K. Regulation and Role of Store-Operated Ca2+ Entry in Cellular Proliferation. In Calcium Entry Channels in Non-Excitable Cells; Kozak, J.A., Putney, J.W., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2018; ISBN 978-1-4987-5272-5. [Google Scholar]
- Derler, I.; Schindl, R.; Fritsch, R.; Heftberger, P.; Riedl, M.C.; Begg, M.; House, D.; Romanin, C. The Action of Selective CRAC Channel Blockers Is Affected by the Orai Pore Geometry. Cell Calcium 2013, 53, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Owsianik, G.; Talavera, K.; Voets, T.; Nilius, B. Permeation and Selectivity of TRP Channels. Annu. Rev. Physiol. 2006, 68, 685–717. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-D.; Srikanth, S.; Yee, M.-K.W.; Mock, D.C.; Lawson, G.W.; Gwack, Y. ORAI1 Deficiency Impairs Activated T Cell Death and Enhances T Cell Survival. J. Immunol. 2011, 187, 3620–3630. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.-T.; Venkatachalam, K.; Parys, J.B.; Gill, D.L. Modification of Store-Operated Channel Coupling and Inositol Trisphosphate Receptor Function by 2-Aminoethoxydiphenyl Borate in DT40 Lymphocytes. J. Biol. Chem. 2002, 277, 6915–6922. [Google Scholar] [CrossRef] [Green Version]
- DeHaven, W.I.; Smyth, J.T.; Boyles, R.R.; Putney, J.W. Calcium Inhibition and Calcium Potentiation of Orai1, Orai2, and Orai3 Calcium Release-Activated Calcium Channels *. J. Biol. Chem. 2007, 282, 17548–17556. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Du, L.; Zhou, Y.; Li, M. Store-Operated CRAC Channel Inhibitors: Opportunities and Challenges. Future Med. Chem. 2016, 8, 817–832. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Mei, Y.; Xia, J.; Hu, H. Orai1 and Orai3 Mediate Store-Operated Calcium Entry Contributing to Neuronal Excitability in Dorsal Root Ganglion Neurons. Front. Cell. Neurosci. 2017, 11, 400. [Google Scholar] [CrossRef]
- Kappel, S.; Kilch, T.; Baur, R.; Lochner, M.; Peinelt, C. The Number and Position of Orai3 Units within Heteromeric Store-Operated Ca2+ Channels Alter the Pharmacology of ICRAC. Int. J. Mol. Sci. 2020, 21, 2458. [Google Scholar] [CrossRef] [Green Version]
- Bomben, V.C.; Sontheimer, H.W. Inhibition of Transient Receptor Potential Canonical Channels Impairs Cytokinesis in Human Malignant Gliomas. Cell Prolif. 2008, 41, 98–121. [Google Scholar] [CrossRef] [PubMed]
- Chokshi, R.; Fruasaha, P.; Kozak, J.A. 2-Aminoethyl Diphenyl Borinate (2-APB) Inhibits TRPM7 Channels through an Intracellular Acidification Mechanism. Channels 2012, 6, 362–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, G.; Montalbetti, N.; Simonin, A.; Danko, T.; Balazs, B.; Zsembery, A.; Hediger, M.A. Inhibition of the Human Epithelial Calcium Channel TRPV6 by 2-Aminoethoxydiphenyl Borate (2-APB). Cell Calcium 2012, 52, 468–480. [Google Scholar] [CrossRef]
- Li, M.; Jiang, J.; Yue, L. Functional Characterization of Homo- and Heteromeric Channel Kinases TRPM6 and TRPM7. J. Gen. Physiol. 2006, 127, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Saotome, K.; McGoldrick, L.L.; Sobolevsky, A.I. Structural Bases of TRP Channel TRPV6 Allosteric Modulation by 2-APB. Nat. Commun. 2018, 9, 2465. [Google Scholar] [CrossRef] [PubMed]
- Goto, J.-I.; Suzuki, A.Z.; Ozaki, S.; Matsumoto, N.; Nakamura, T.; Ebisui, E.; Fleig, A.; Penner, R.; Mikoshiba, K. Two Novel 2-Aminoethyl Diphenylborinate (2-APB) Analogues Differentially Activate and Inhibit Store-Operated Ca2+ Entry via STIM Proteins. Cell Calcium 2010, 47, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hendron, E.; Wang, X.; Zhou, Y.; Cai, X.; Goto, J.-I.; Mikoshiba, K.; Baba, Y.; Kurosaki, T.; Wang, Y.; Gill, D.L. Potent Functional Uncoupling between STIM1 and Orai1 by Dimeric 2-Aminodiphenyl Borinate Analogs. Cell Calcium 2014, 56, 482–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Y.; Zhou, Y.; Hendron, E.; Mancarella, S.; Andrake, M.D.; Rothberg, B.S.; Soboloff, J.; Gill, D.L. Distinct Orai-Coupling Domains in STIM1 and STIM2 Define the Orai-Activating Site. Nat. Commun. 2014, 5, 3183. [Google Scholar] [CrossRef] [Green Version]
- Núñez, L.; Valero, R.A.; Senovilla, L.; Sanz-Blasco, S.; García-Sancho, J.; Villalobos, C. Cell Proliferation Depends on Mitochondrial Ca2+ Uptake: Inhibition by Salicylate. J. Physiol. 2006, 571, 57–73. [Google Scholar] [CrossRef]
- Burn, J.; Mathers, J.C.; Bishop, D.T. Chemoprevention in Lynch Syndrome. Fam. Cancer 2013, 12, 707–718. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Sun, J.; Huang, M.-Y.; Wang, Y.-S.; Hou, M.-F.; Sun, Y.; He, H.; Krishna, N.; Chiu, S.-J.; Lin, S.; et al. STIM1 Overexpression Promotes Colorectal Cancer Progression, Cell Motility and COX-2 Expression. Oncogene 2015, 34, 4358–4367. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, E.; Valero, R.A.; Quintana, A.; Hoth, M.; Núñez, L.; Villalobos, C. Nonsteroidal Anti-Inflammatory Drugs Inhibit Vascular Smooth Muscle Cell Proliferation by Enabling the Ca2+-Dependent Inactivation of Calcium Release-Activated Calcium/Orai Channels Normally Prevented by Mitochondria. J. Biol. Chem. 2011, 286, 16186–16196. [Google Scholar] [CrossRef] [Green Version]
- Cabello, O.A.; Schilling, W.P. Vectorial Ca2+ Flux from the Extracellular Space to the Endoplasmic Reticulum via a Restricted Cytoplasmic Compartment Regulates Inositol 1,4,5-Trisphosphate-Stimulated Ca2+ Release from Internal Stores in Vascular Endothelial Cells. Biochem. J. 1993, 295, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, Z.; Sui, X.; Yao, J.; Xie, J.; Jiang, L.; Zhou, Y.; Pan, H.; Han, W. SKF-96365 Activates Cytoprotective Autophagy to Delay Apoptosis in Colorectal Cancer Cells through Inhibition of the Calcium/CaMKIIγ/AKT-Mediated Pathway. Cancer Lett. 2016, 372, 226–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, S.; Sun, Y.; Sukumaran, P.; Singh, B.B. Resveratrol Activates Autophagic Cell Death in Prostate Cancer Cells via Downregulation of STIM1 and the MTOR Pathway. Mol. Carcinog. 2016, 55, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ren, Y.; Wang, L.; Zhao, W.; Dong, X.; Pan, J.; Gao, H.; Tian, Y. Orai1 and Stim1 Mediate the Majority of Store-Operated Calcium Entry in Multiple Myeloma and Have Strong Implications for Adverse Prognosis. Cell. Physiol. Biochem. 2018, 48, 2273–2285. [Google Scholar] [CrossRef]
- Zeng, B.; Yuan, C.; Yang, X.; Atkin, S.; Xu, S.-Z. TRPC Channels and Their Splice Variants Are Essential for Promoting Human Ovarian Cancer Cell Proliferation and Tumorigenesis. Curr. Cancer Drug Targets 2012, 13. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J., J.; Huang, X.-Y. Orai1 and STIM1 Are Critical for Breast Tumor Cell Migration and Metastasis | Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S1535610808004388?token=AB3AA70BCD2C6D456CAC6DC1FAB05D8586491B411FAAA4921EEC653FF41FE3379709A8FF19E4A80ECCEBB79CB59359B3&originRegion=eu-west-1&originCreation=20220514185214 (accessed on 14 May 2022).
- Dago, C.D.; Maux, P.L.; Roisnel, T.; Brigaudeau, C.; Bekro, Y.-A.; Mignen, O.; Bazureau, J.-P. Preliminary Structure-Activity Relationship (SAR) of a Novel Series of Pyrazole SKF-96365 Analogues as Potential Store-Operated Calcium Entry (SOCE) Inhibitors. Int. J. Mol. Sci. 2018, 19, 856. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Rubaiy, H.N.; Chen, G.; Hallett, T.; Zaibi, N.; Zeng, B.; Saurabh, R.; Xu, S. Mibefradil, a T-type Ca2+ Channel Blocker Also Blocks Orai Channels by Action at the Extracellular Surface. Br. J. Pharmacol. 2019, 176, 3845–3856. [Google Scholar] [CrossRef]
- Zeng, B.; Chen, G.-L.; Daskoulidou, N.; Xu, S.-Z. The Ryanodine Receptor Agonist 4-Chloro-3-Ethylphenol Blocks ORAI Store-Operated Channels. Br. J. Pharmacol. 2014, 171, 1250–1259. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.-Z.; Muraki, K.; Zeng, F.; Li, J.; Sukumar, P.; Shah, S.; Dedman, A.M.; Flemming, P.K.; McHugh, D.; Naylor, J.; et al. A Sphingosine-1–Phosphate-Activated Calcium Channel Controlling Vascular Smooth Muscle Cell Motility. Circ. Res. 2006, 98, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Zitt, C.; Strauss, B.; Schwarz, E.C.; Spaeth, N.; Rast, G.; Hatzelmann, A.; Hoth, M. Potent Inhibition of Ca2+ Release-Activated Ca2+ Channels and T-Lymphocyte Activation by the Pyrazole Derivative BTP2. J. Biol. Chem. 2004, 279, 12427–12437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putney, J.W. Pharmacology of Capacitative Calcium Entry. Mol. Interv. 2001, 1, 84–94. [Google Scholar]
- McNally, B.A.; Yamashita, M.; Engh, A.; Prakriya, M. Structural Determinants of Ion Permeation in CRAC Channels. Proc. Natl. Acad. Sci. USA 2009, 106, 22516–22521. [Google Scholar] [CrossRef] [Green Version]
- Ohga, K.; Takezawa, R.; Arakida, Y.; Shimizu, Y.; Ishikawa, J. Characterization of YM-58483/BTP2, a Novel Store-Operated Ca2+ Entry Blocker, on T Cell-Mediated Immune Responses in Vivo. Int. Immunopharmacol. 2008, 8, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- He, L.-P.; Hewavitharana, T.; Soboloff, J.; Spassova, M.A.; Gill, D.L. A Functional Link between Store-Operated and TRPC Channels Revealed by the 3,5-Bis(Trifluoromethyl)Pyrazole Derivative, BTP2 *. J. Biol. Chem. 2005, 280, 10997–11006. [Google Scholar] [CrossRef] [Green Version]
- Takezawa, R.; Cheng, H.; Beck, A.; Ishikawa, J.; Launay, P.; Kubota, H.; Kinet, J.-P.; Fleig, A.; Yamada, T.; Penner, R. A Pyrazole Derivative Potently Inhibits Lymphocyte Ca2+ Influx and Cytokine Production by Facilitating Transient Receptor Potential Melastatin 4 Channel Activity. Mol. Pharmacol. 2006, 69, 1413–1420. [Google Scholar] [CrossRef]
- Azimi, I.; Flanagan, J.U.; Stevenson, R.J.; Inserra, M.; Vetter, I.; Monteith, G.R.; Denny, W.A. Evaluation of Known and Novel Inhibitors of Orai1-Mediated Store Operated Ca 2+ Entry in MDA-MB-231 Breast Cancer Cells Using a Fluorescence Imaging Plate Reader Assay. Bioorg. Med. Chem. 2017, 25, 440–449. [Google Scholar] [CrossRef] [Green Version]
- Ng, L.C.; McCormack, M.D.; Airey, J.A.; Singer, C.A.; Keller, P.S.; Shen, X.-M.; Hume, J.R. TRPC1 and STIM1 Mediate Capacitative Ca2+ Entry in Mouse Pulmonary Arterial Smooth Muscle Cells. J. Physiol. 2009, 587, 2429–2442. [Google Scholar] [CrossRef]
- Li, J.; McKeown, L.; Ojelabi, O.; Stacey, M.; Foster, R.; O’Regan, D.; Porter, K.E.; Beech, D.J. Nanomolar Potency and Selectivity of a Ca2+ Release-Activated Ca2+ Channel Inhibitor against Store-Operated Ca2+ Entry and Migration of Vascular Smooth Muscle Cells. Br. J. Pharmacol. 2011, 164, 382–393. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Panicker, S.; Lau, K.-Y.; Apparsundaram, S.; Patel, V.A.; Chen, S.-L.; Soto, R.; Jung, J.K.C.; Ravindran, P.; Okuhara, D.; et al. Characterization of a Novel CRAC Inhibitor That Potently Blocks Human T Cell Activation and Effector Functions. Mol. Immunol. 2013, 54, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Velicelebi, G.; Stauderman, K.; Dunn, M.; Roos, J. Pancreatitis Treatment 2016. WO2016138472A1, 26 February 2016. [Google Scholar]
- Ramos, S.; Grigoryev, S.; Rogers, E.; Roos, J.; Whitten, J.; Stauderman, K.; Velicelebi, G. CM3457, a Potent and Selective Oral CRAC Channel Inhibitor, Suppresses T and Mast Cell Function and Is Efficacious in Rat Models of Arthritis and Asthma (72.3). J. Immunol. 2012, 188, 72–73. [Google Scholar] [CrossRef]
- Sadaghiani, A.M.; Lee, S.M.; Odegaard, J.I.; Leveson-Gower, D.B.; McPherson, O.M.; Novick, P.; Kim, M.R.; Koehler, A.N.; Negrin, R.; Dolmetsch, R.E.; et al. Identification of Orai1 Channel Inhibitors by Using Minimal Functional Domains to Screen Small Molecule Microarrays. Chem. Biol. 2014, 21, 1278–1292. [Google Scholar] [CrossRef] [PubMed]
- Smyth, J.T.; DeHaven, W.I.; Bird, G.S.; Putney, J.W. Calcium store-dependent and independent reversal of stim1 localization and function. J. Cell Sci. 2008, 121, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Kitani, A.; Murray, P.J.; Wakatsuki, Y.; Fuss, I.J.; Strober, W. Nucleotide Binding Oligomerization Domain 2 Deficiency Leads to Dysregulated TLR2 Signaling and Induction of Antigen-Specific Colitis. Immunity 2006, 25, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Bartoli, F.; Bailey, M.A.; Rode, B.; Mateo, P.; Antigny, F.; Bedouet, K.; Gerbaud, P.; Gosain, R.; Plante, J.; Norman, K.; et al. Orai1 Channel Inhibition Preserves Left Ventricular Systolic Function and Normal Ca2+ Handling After Pressure Overload. Circulation 2020, 141, 199–216. [Google Scholar] [CrossRef]
- Barde, P.J.; Viswanadha, S.; Veeraraghavan, S.; Vakkalanka, S.V.; Nair, A. A First-in-Human Study to Evaluate the Safety, Tolerability and Pharmacokinetics of RP3128, an Oral Calcium Release-Activated Calcium (CRAC) Channel Modulator in Healthy Volunteers. J. Clin. Pharm. Ther. 2021, 46, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Sutovska, M.; Kocmalova, M.; Franova, S.; Vakkalanka, S.; Viswanadha, S. Pharmacodynamic Evaluation of RP3128, a Novel and Potent CRAC Channel Inhibitor in Guinea Pig Models of Allergic Asthma. Eur. J. Pharmacol. 2016, 772, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Vakkalanka, S.; Merikapudi, G.; Babu, G.; Routhu, K.; Veeraraghavan, S.; Viswanadha, S. Pre-Clinical Characterization of RP3128, a Novel and Potent CRAC Channel Inhibitor for the Treatment of Respiratory Disorders . Available online: https://www.ers-education.org/lr/show-details/?idP=127076 (accessed on 16 May 2022).
- Rhizen Pharmaceuticals SA. A Phase I/Ib, Study to Evaluate Safety and Efficacy of RP4010, in Patients With Relapsed or Refractory Lymphomas. 2019. Available online: https://cinicaltrials.gov (accessed on 14 May 2022).
- Pallagi, P.; Görög, M.; Papp, N.; Madácsy, T.; Varga, Á.; Crul, T.; Szabó, V.; Molnár, M.; Dudás, K.; Grassalkovich, A.; et al. Bile Acid- and Ethanol-Mediated Activation of Orai1 Damages Pancreatic Ductal Secretion in Acute Pancreatitis. J. Physiol. 2022, 600, 1631–1650. [Google Scholar] [CrossRef]
- Cordero-Sanchez, C.; Pessolano, E.; Riva, B.; Vismara, M.; Trivigno, S.M.G.; Clemente, N.; Aprile, S.; Ruffinatti, F.A.; Portararo, P.; Filigheddu, N.; et al. CIC-39Na Reverses the Thrombocytopenia That Characterizes Tubular Aggregate Myopathy. Blood Adv. 2022, 6, 4471–4484. [Google Scholar] [CrossRef]
- Aprile, S.; Riva, B.; Bhela, I.P.; Cordero-Sanchez, C.; Avino, G.; Genazzani, A.A.; Serafini, M.; Pirali, T. 1,2,4-Oxadiazole-Bearing Pyrazoles as Metabolically Stable Modulators of Store-Operated Calcium Entry. ACS Med. Chem. Lett. 2021, 12, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Stanisz, H.; Saul, S.; Müller, C.S.L.; Kappl, R.; Niemeyer, B.A.; Vogt, T.; Hoth, M.; Roesch, A.; Bogeski, I. Inverse Regulation of Melanoma Growth and Migration by Orai1/STIM2-Dependent Calcium Entry. Pigment Cell Melanoma Res. 2014, 27, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.C.; McDonald, T.V.; Gardner, P. Inhibition by SK&F 96365 of Ca2+ Current, IL-2 Production and Activation in T Lymphocytes. Br. J. Pharmacol. 1994, 113, 861–868. [Google Scholar] [PubMed]
- Rothberg, B.S.; Wang, Y.; Gill, D.L. Orai Channel Pore Properties and Gating by STIM: Implications from the Orai Crystal Structure. Sci. Signal. 2013, 6, pe9. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Diseases | Subunit Combination | Reference(s) |
|---|---|---|
| Immunodeficiency | Orai1/STIM1 | [33] |
| Myopathy | Orai1/STIM1 | [37] |
| Tubular aggregate myopathy | Orai1/STIM1 | [36] |
| Stormorken syndrome | Orai1/STIM1 | [44] |
| York platelet syndrome | Orai1/STIM1 | [46] |
| Cardiovascular and metabolic disorders | Oria1/STIM1 | [75] |
| Cancers | Oria1/STIM1 ORAI3 (lung cancer) STIM2 (tumour growth) | [60] [68] [63] |
| Kidney | Orai1 | [73] |
| Name | IC50 | Cell Types | Reference |
|---|---|---|---|
| GSK-5503A & GSK-7975A | 4 μM | HEK-293 | [85] |
| 5J-4 | 807 nM (peak) 195 nM (sustained) | Primary murine effector T TH17 HeLa-O + S cells | [87] |
| DPB162-AE | 200 nM | HEK-293 | [99] |
| 4-CEP | 203.6 µM | HEK-293 | [113] |
| 4-CmC | 830.9 1 µM | HEK-293 | [113] |
| 4-CIP | 1437.1 µM | HEK-293 | [113] |
| SKF96365 | 12 µM | T leukaemic | [139] |
| BTP2 (YM-58483) | 2.8 µM | MDA-MB-231 breast cancer cells | [121] |
| Synta-66 | 1.4 μM | Rat basophilic leukaemia (RBL) | [122] |
| ML-9 | 10 μM | HEK-293 | [128,129] |
| JPIII | 399 nM | HEK-293 | [130] |
| Pyr 39 | 4.4 μM | HEK-293 | [82] |
| RO2959 | 265 nM | CD4+T lymphocytes | [124] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubaiy, H.N. ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets. Pharmaceuticals 2023, 16, 162. https://doi.org/10.3390/ph16020162
Rubaiy HN. ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets. Pharmaceuticals. 2023; 16(2):162. https://doi.org/10.3390/ph16020162
Chicago/Turabian StyleRubaiy, Hussein N. 2023. "ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets" Pharmaceuticals 16, no. 2: 162. https://doi.org/10.3390/ph16020162
APA StyleRubaiy, H. N. (2023). ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets. Pharmaceuticals, 16(2), 162. https://doi.org/10.3390/ph16020162