
ACW-02 an Acridine Triazolidine Derivative Presents Antileishmanial Activity Mediated by DNA Interaction and Immunomodulation

 , ,
, ,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
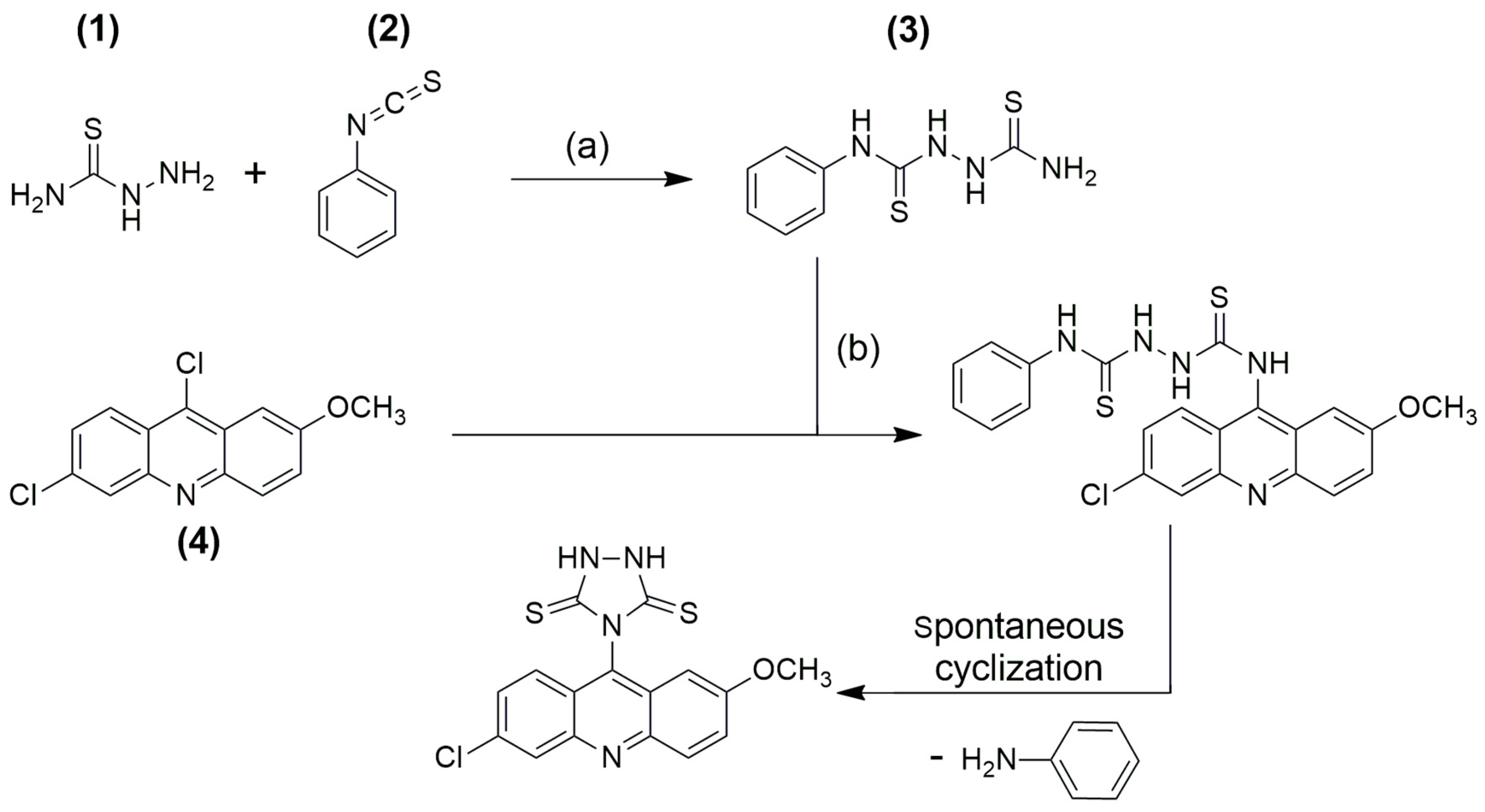
2.1. Synthesis and Characterization
2.2. Antileishmanial and Cytotoxic Activity Evaluation
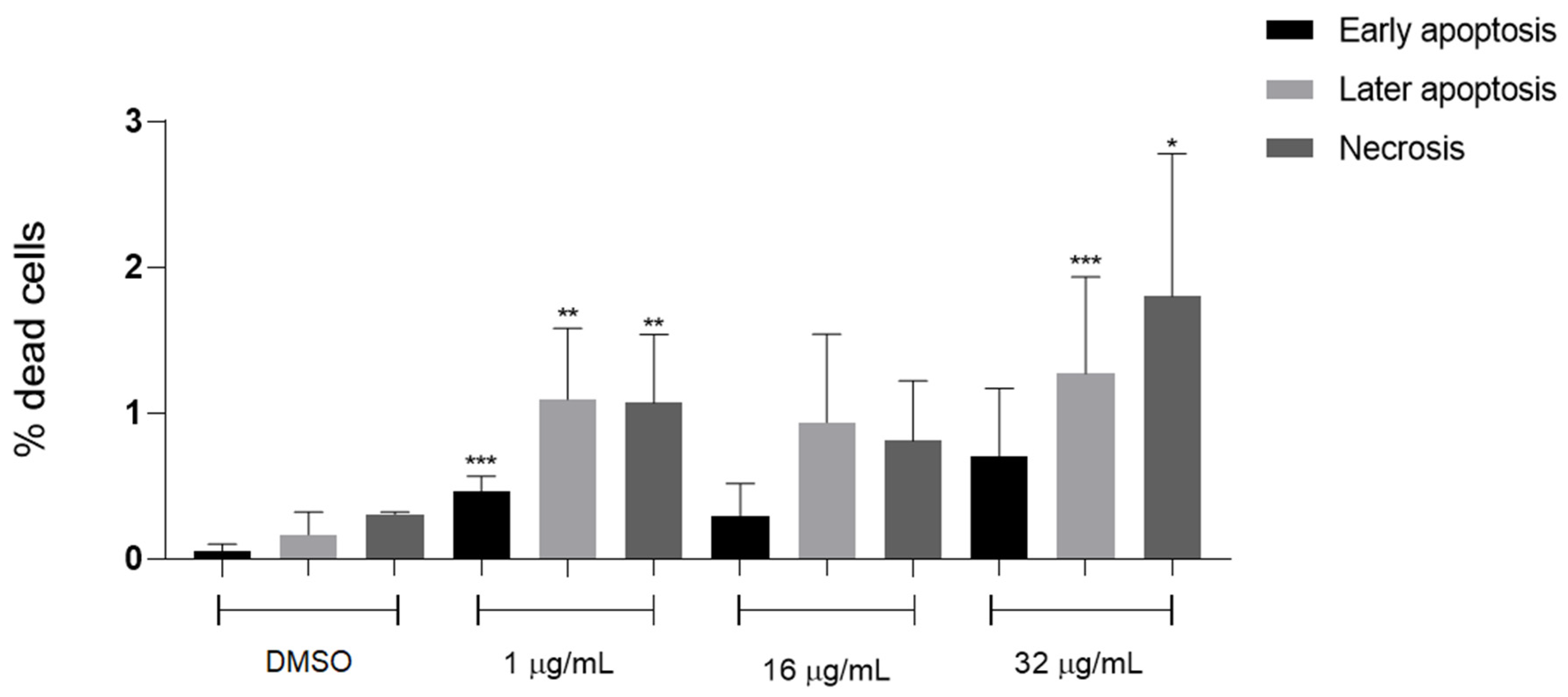
2.3. Cytotoxicity Evaluation in Macrophages by Annexin V-FITC/PI Double Staining Assay
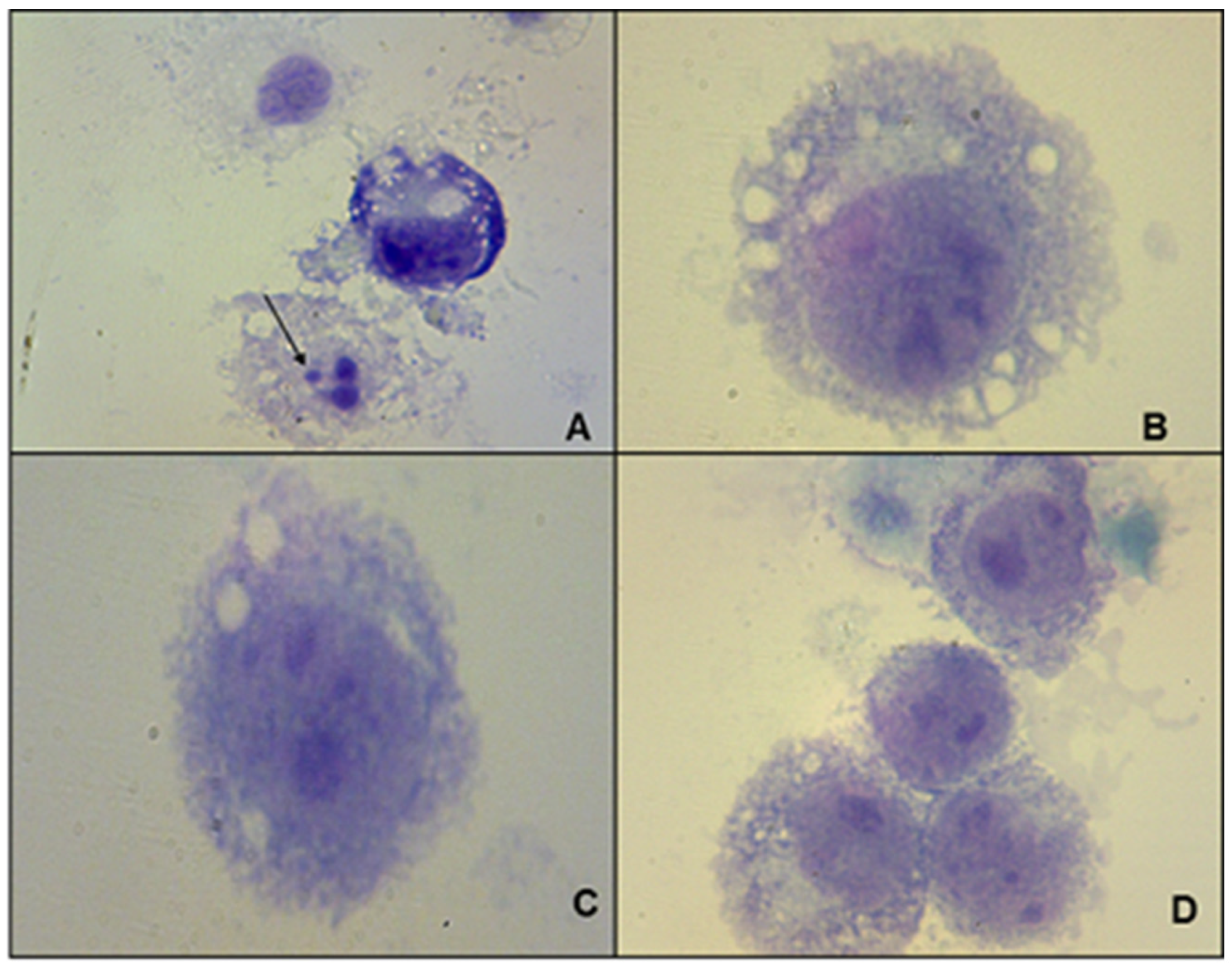
2.4. Determination of the Microbicidal Effect in Cultures of J774 Macrophages Infected with L. amazonensis
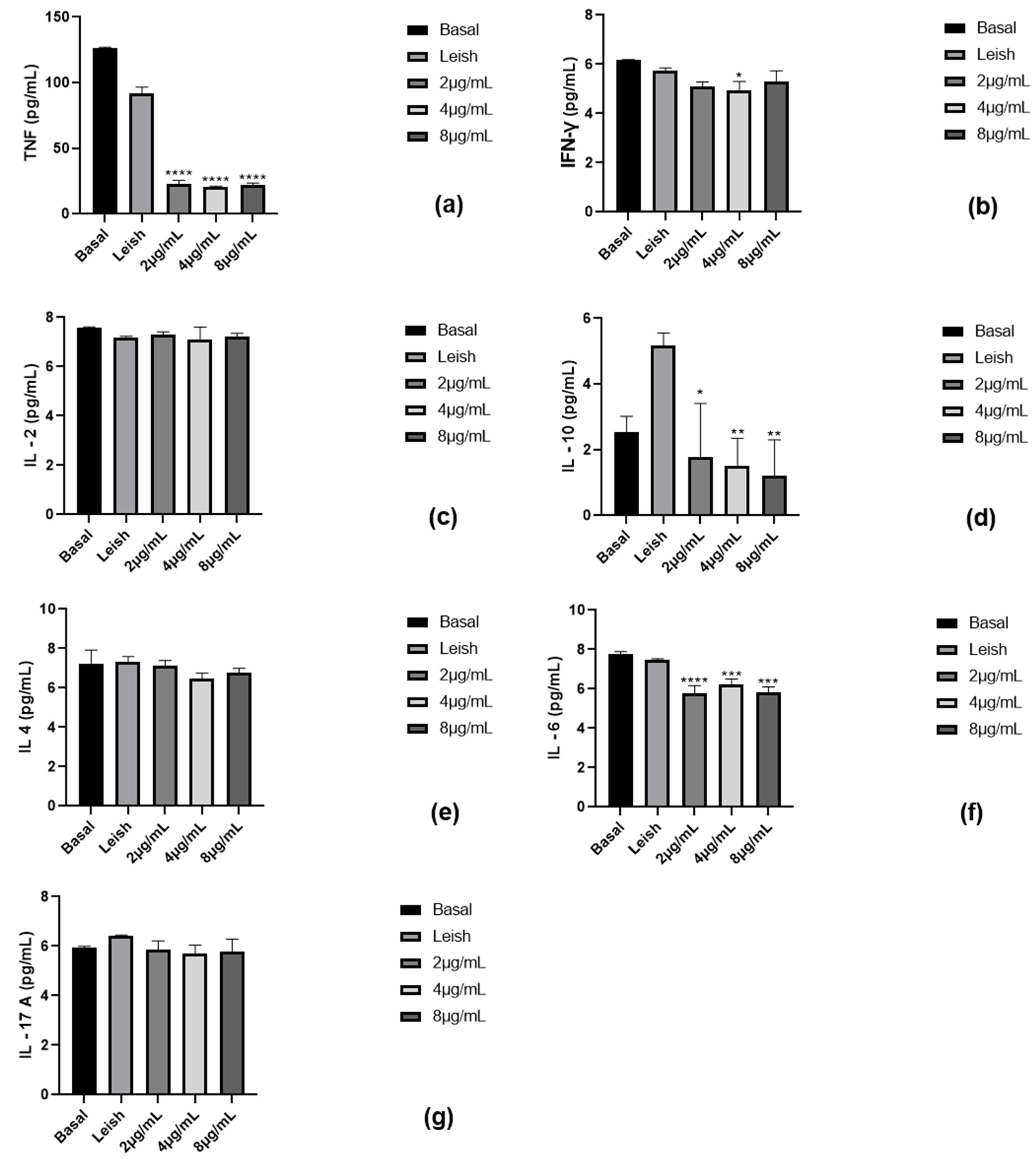
2.5. Assessment of Cytokine Production by Macrophages
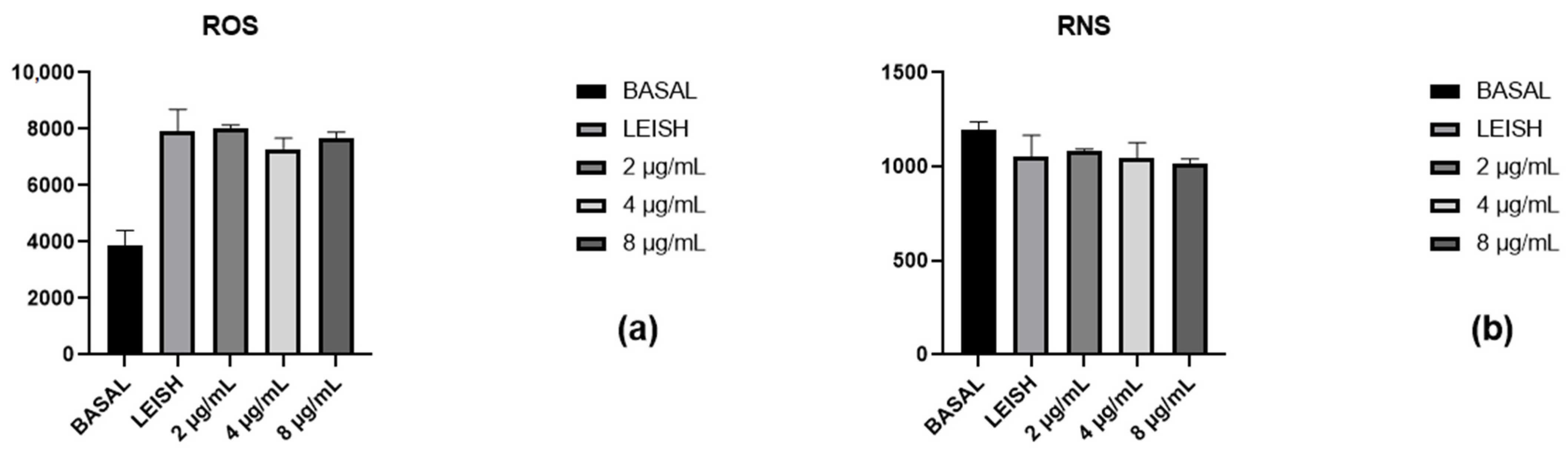
2.6. Reactive Oxygen and Nitrogen Species Evaluation
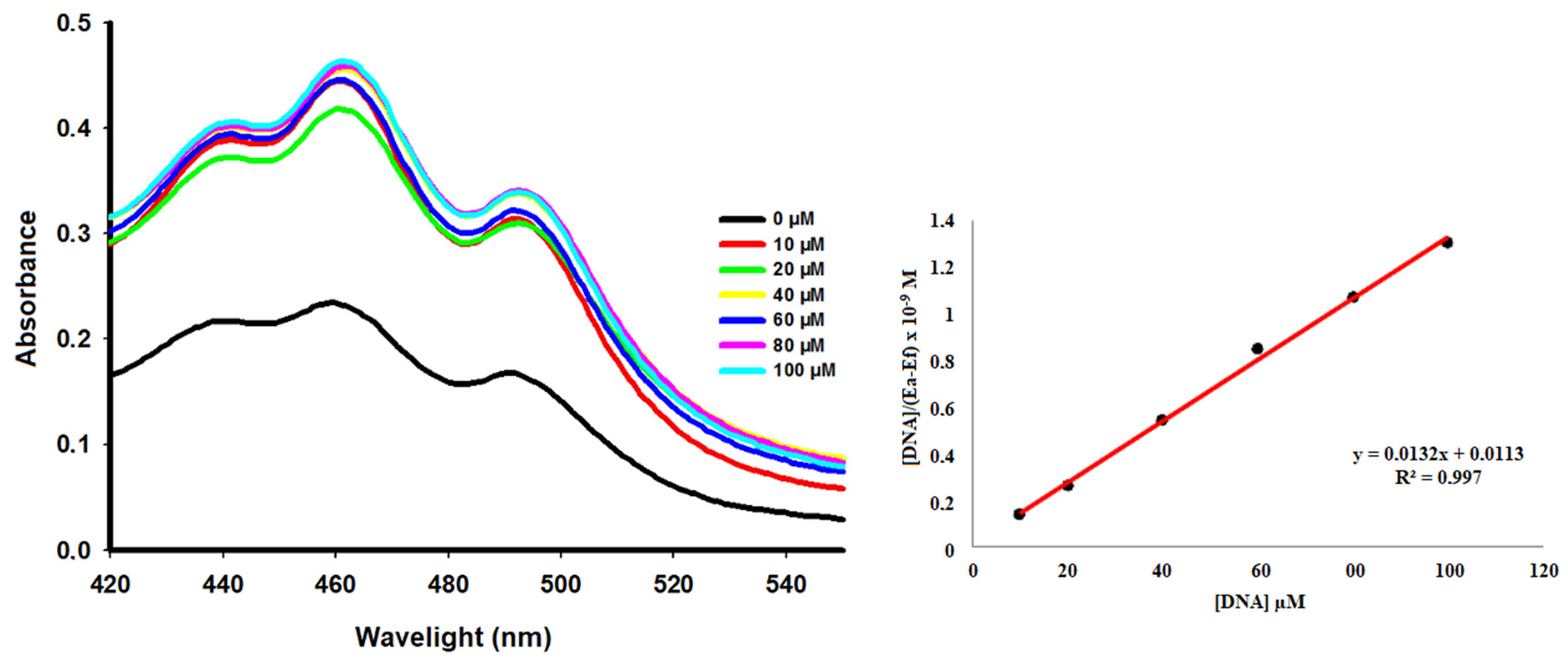
2.7. Interaction with DNA
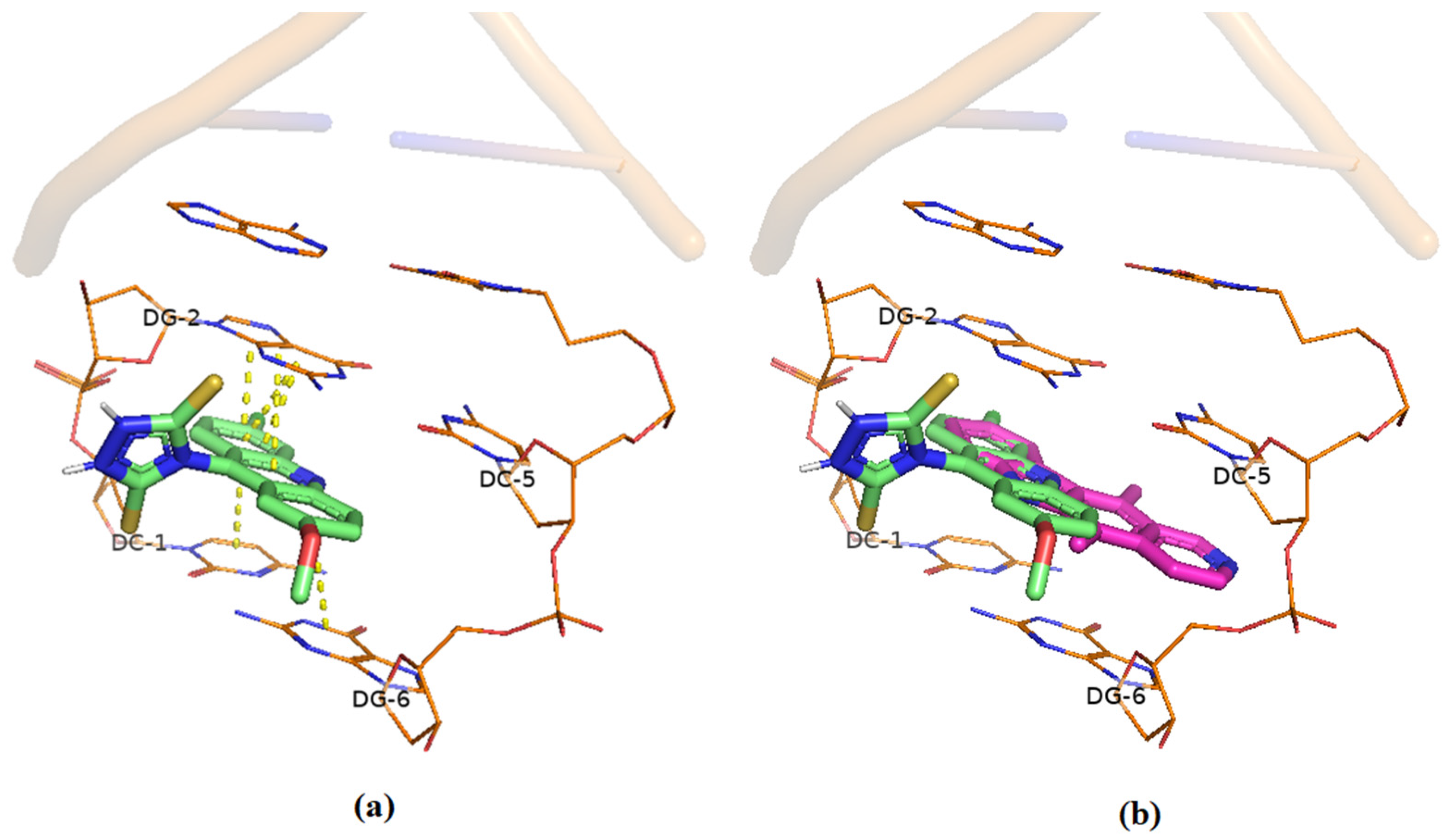
Docking Studies of Interaction with DNA
2.8. Theoretical Study of Possible Mechanisms of Action
2.8.1. Molecular Docking

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 4APN (TryR) | 3L4D (CYP51) | 2B9S (Topo I) | CPBLa | |
|---|---|---|---|---|
| ACW-02 | 54.22 | 58.74 | 60.34 | 50.97 |
| Standard * | 67.58 | 78.77 | 63.98 | 47.77 |
| RMSD | 0.76 | 1.02 | - | - |
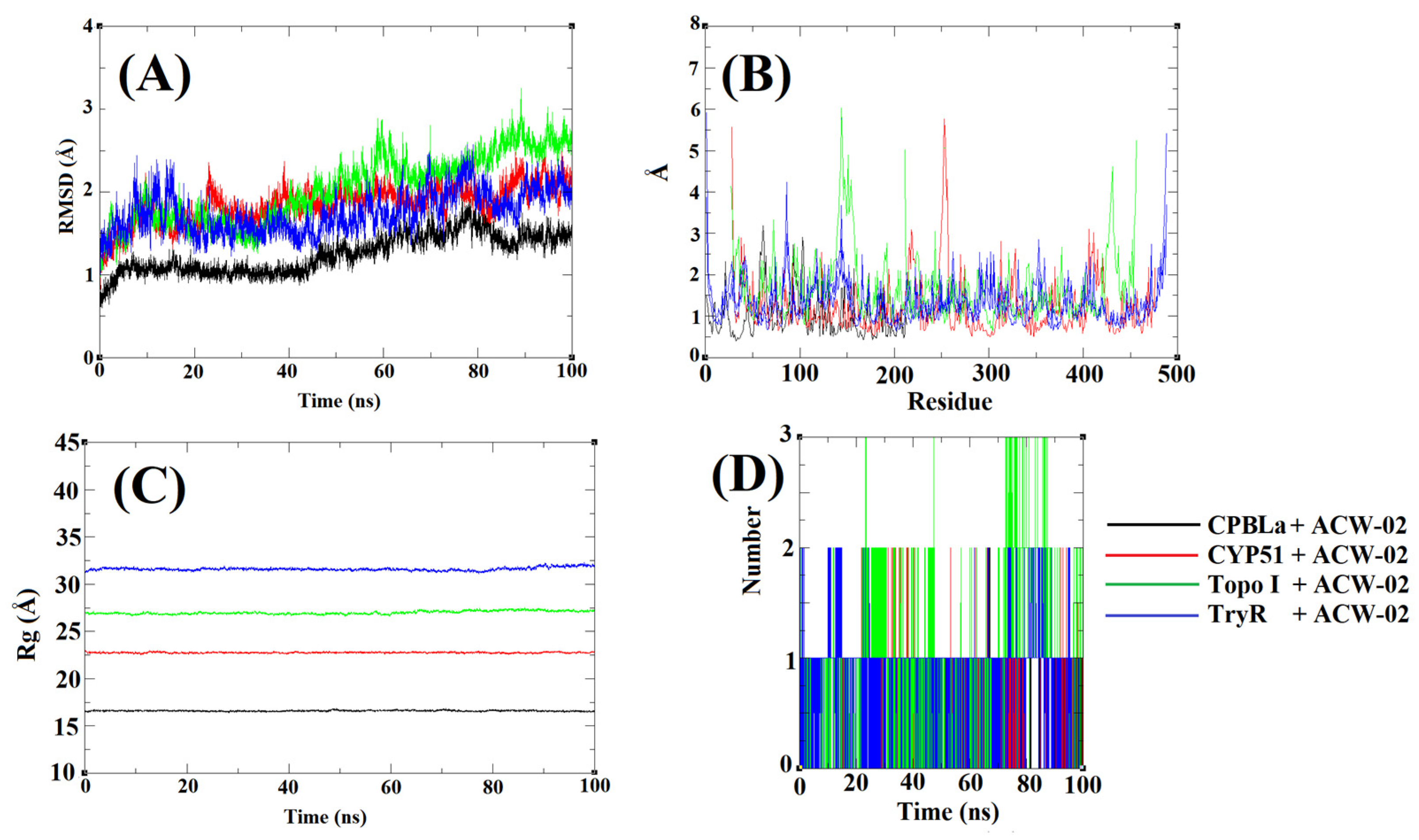
2.8.2. Molecular Dynamics (MD) Simulations
2.8.3. MM-PBSA Calculations
3. Discussion
4. Materials and Methods
4.1. General Procedure for the Synthesis of Triazolidine Acridine Derivative
4-(6-Chloro-2-methoxyacridin-9-yl)-1,2,4-triazolidine-3,5-dithione (ACW-02)
4.2. Antileishmanial and Cytotoxic Activity
4.2.1. Cultures of L. amazonensis
4.2.2. Macrophage Cultive
4.2.3. Determination of the Inhibitory and Cytotoxic Effects in L. amazonensis and JJ74 Macrophage Cultures
4.2.4. Cytotoxicity Evaluation in Macrophages by Annexin V-FITC/PI Double Staining Assay
4.2.5. Microbicidal Assay
4.2.6. Assessment of Cytokine Production by Macrophages
4.2.7. Evaluation of the Production of Reactive Oxygen and Nitrogen Species
4.3. ctDNA Interaction Evaluation by UV–vis Absorption
4.4. Statistical Analysis
4.5. Homology Modeling
4.6. Molecular Docking Studies
4.7. Molecular Dynamics Simulations
4.8. MM-PBSA Calculations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ready, P.D. Biology of Phlebotomine Sand Flies as Vectors of Disease Agents. Annu. Rev. Entomol. 2013, 58, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Valdivieso, E.; Mejías, F.; Torrealba, C.; Benaim, G.; Kouznetsov, V.V.; Sojo, F.; Rojas-Ruiz, F.A.; Arvelo, F.; Dagger, F. In Vitro 4-Aryloxy-7-Chloroquinoline Derivatives Are Effective in Mono- and Combined Therapy against Leishmania Donovani and Induce Mitocondrial Membrane Potential Disruption. Acta Trop. 2018, 183, 36–42. [Google Scholar] [CrossRef]
- World Health Organization. Leishmaniasis. Available online: https://www.who.int/health-topics/leishmaniasis (accessed on 29 July 2022).
- Anversa, L.s.; Tiburcio, M.G.S.; Richini-Pereira, V.n.B.; Ramirez, L.E. Human Leishmaniasis in Brazil: A General Review. Rev. Assoc. Med. Bras. 2018, 64, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, A.; Kushwaha, P.; Gupta, S.; Dodda, R.P.; Ramalingam, K.; Kant, R.; Goyal, N.; Sashidhara, K.V. Synthesis and Evaluation of Novel Triazolyl Quinoline Derivatives as Potential Antileishmanial Agents. Eur. J. Med. Chem. 2018, 154, 172–181. [Google Scholar] [CrossRef]
- Soulat, D.; Bogdan, C. Function of Macrophage and Parasite Phosphatases in Leishmaniasis. Front. Immunol. 2017, 8, 1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podinovskaia, M.; Descoteaux, A. Leishmania and the Macrophage: A Multifaceted Interaction. Future Microbiol. 2015, 10, 111–129. [Google Scholar] [CrossRef]
- Mcgwire, B.S.; Satoskar, A.R. Leishmaniasis: Clinical Syndromes and Treatment. QJM Int. J. Med. 2014, 107, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, A.; Chandrakar, P.; Gupta, S.; Parmar, N.; Singh, S.K.; Rashid, M.; Kushwaha, P.; Wahajuddin, M.; Sashidhara, K.V.; Kar, S. Synthesis, Biological Evaluation, Structure-Activity Relationship, and Mechanism of Action Studies of Quinoline-Metronidazole Derivatives Against Experimental Visceral Leishmaniasis. J. Med. Chem. 2019, 62, 5655–5671. [Google Scholar] [CrossRef] [PubMed]
- Almeida, F.S.; Sousa, G.L.S.; Rocha, J.C.; Ribeiro, F.F.; de Oliveira, M.R.; de Lima Grisi, T.C.S.; Araújo, D.A.M.; Michelangela, M.S.; Castro, R.N.; Amaral, I.P.G.; et al. In Vitro Anti-Leishmania Activity and Molecular Docking of Spiro-Acridine Compounds as Potential Multitarget Agents against Leishmania Infantum. Bioorg. Med. Chem. Lett. 2021, 49, 128289. [Google Scholar] [CrossRef]
- Albino, S.L.; da Silva, J.M.; de Caldas Nobre, M.S.; de Medeiros E Silva, Y.M.S.; Santos, M.B.; de Araújo, R.S.A.; do Carmo Alves de Lima, M.; Schmitt, M.; de Moura, R.O. Bioprospecting of Nitrogenous Heterocyclic Scaffolds with Potential Action for Neglected Parasitosis: A Review. Curr. Pharm. Des. 2020, 26, 4112–4150. [Google Scholar] [CrossRef]
- De Lima Serafim, V.; Félix, M.B.; Frade Silva, D.K.; Rodrigues, K.A.d.F.; Andrade, P.N.; de Almeida, S.M.V.; de Albuquerque dos Santos, S.; de Oliveira, J.F.; de Lima, M.d.C.A.; Mendonça-Junior, F.J.B.; et al. New Thiophene–Acridine Compounds: Synthesis, Antileishmanial Activity, DNA Binding, Chemometric, and Molecular Docking Studies. Chem. Biol. Drug Des. 2018, 91, 1141–1155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, B.; Zhang, W.; Yang, T.; Wang, N.; Gao, C.; Tan, C.; Liu, H.; Jiang, Y. Synthesis and Antiproliferative Activity of 9-Benzylamino-6-Chloro-2-Methoxy-Acridine Derivatives as Potent DNA-Binding Ligands and Topoisomerase II Inhibitors. Eur. J. Med. Chem. 2016, 116, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, J.; Saquib, M.; Singh, S.; Tufail, F.; Singh, J.; Singh, J. Catalyst-Free Glycerol-Mediated Green Synthesis of 5′-Thioxospiro[Indoline-3,3′-[1,2,4]Triazolidin]-2-Ones/Spiro[Indoline-3,3′-[1,2,4]Triazolidine]-2,5′-Diones. Synth. Commun. 2017, 47, 1999–2006. [Google Scholar] [CrossRef]
- Tiwari, N.; Gedda, M.R.; Tiwari, V.K.; Singh, S.P.; Singh, R.K. Limitations of Current Therapeutic Options, Possible Drug Targets and Scope of Natural Products in Control of Leishmaniasis. Mini-Rev. Med. Chem. 2017, 18, 26–41. [Google Scholar] [CrossRef]
- Gonçalves-de-Albuquerque, S.d.C.; Pessoa-e-Silva, R.; Trajano-Silva, L.A.M.; de Goes, T.C.; de Morais, R.C.S.; Oliveira, C.N.d.C.; de Lorena, V.M.B.; de Paiva-Cavalcanti, M. The Equivocal Role of Th17 Cells and Neutrophils on Immunopathogenesis of Leishmaniasis. Front. Immunol. 2017, 8, 1437. [Google Scholar] [CrossRef] [Green Version]
- Mirzaei, A.; Maleki, M.; Masoumi, E.; Maspi, N. A Historical Review of the Role of Cytokines Involved in Leishmaniasis. Cytokine 2021, 145, 155297. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, Á.; Valério-Bolas, A.; Palma-Marques, J.; Mourata-Gonçalves, P.; Ruas, P.; Dias-Guerreiro, T.; Santos-Gomes, G. Cutaneous Leishmaniasis: The Complexity of Host’s Effective Immune Response against a Polymorphic Parasitic Disease. J. Immunol. Res. 2019, 2019, 2603730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesmer, L.A.; Lundy, S.K.; Sarkar, S.; Fox, D.A. Th17 Cells in Human Disease. Immunol. Rev. 2008, 223, 87–113. [Google Scholar] [CrossRef]
- Tomiotto-Pellissier, F.; Bortoleti, B.T.d.S.; Assolini, J.P.; Gonçalves, M.D.; Carloto, A.C.M.; Miranda-Sapla, M.M.; Conchon-Costa, I.; Bordignon, J.; Pavanelli, W.R. Macrophage Polarization in Leishmaniasis: Broadening Horizons. Front. Immunol. 2018, 9, 2529. [Google Scholar] [CrossRef] [Green Version]
- Kima, P.E. Leishmania Molecules That Mediate Intracellular Pathogenesis. Microbes Infect. 2014, 16, 721–726. [Google Scholar] [CrossRef]
- McGhee, J.D.; von Hippel, P.H. Theoretical Aspects of DNA-Protein Interactions: Co-Operative and Non-Co-Operative Binding of Large Ligands to a One-Dimensional Homogeneous Lattice. J. Mol. Biol. 1974, 86, 469–489. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.; Joshi, R.; Ottoor, D. Spectroscopic and Molecular Docking Study to Understand the Binding Interaction of Rosiglitazone with Bovine Serum Albumin in Presence of Valsartan. J. Lumin. 2018, 197, 200–210. [Google Scholar] [CrossRef]
- Tokala, R.; Thatikonda, S.; Sana, S.; Regur, P.; Godugu, C.; Shankaraiah, N. Synthesis and in Vitro Cytotoxicity Evaluation of β-Carboline-Linked 2,4-Thiazolidinedione Hybrids: Potential DNA Intercalation and Apoptosis-Inducing Studies. New J. Chem. 2018, 42, 16226–16236. [Google Scholar] [CrossRef]
- Wang, H.; Cai, Z.; Zheng, S.; Ma, H.; Lin, H.; Zheng, X. Design, Synthesis and Biological Evaluation of Some Novel Thiazole-2- Carboxamide Derivatives as Antitumor Agents. Lett. Drug Des. Discov. 2017, 15, 388–397. [Google Scholar] [CrossRef]
- Zhou, Y.; Song, T.; Cao, Y.; Gong, G.; Zhang, Y.; Zhao, H.; Zhao, G. Synthesis and Characterization of Planar Chiral Cyclopalladated Ferrocenylimines: DNA/HSA Interactions and in Vitro Cytotoxic Activity. J. Organomet. Chem. 2018, 871, 1–9. [Google Scholar] [CrossRef]
- Aneja, B.; Arif, R.; Perwez, A.; Napoleon, J.V.; Hasan, P.; Rizvi, M.M.A.; Azam, A.; Rahisuddin; Abid, M. N-Substituted 1,2,3-Triazolyl-Appended Indole-Chalcone Hybrids as Potential DNA Intercalators Endowed with Antioxidant and Anticancer Properties. ChemistrySelect 2018, 3, 2638–2645. [Google Scholar] [CrossRef]
- Sama, F.; Raizada, M.; Ashafaq, M.; Ahamad, M.N.; Mantasha, I.; Iman, K.; Shahid, M.; Rahisuddin; Arif, R.; Shah, N.A.; et al. Synthesis, Structure and DNA Binding Properties of a Homodinuclear Cu(II) Complex: An Experimental and Theoretical Approach. J. Mol. Struct. 2019, 1176, 283–289. [Google Scholar] [CrossRef]
- Ihmels, H.; Otto, D. Intercalation of Organic Dye Molecules into Double-Stranded DNA—General Principles and Recent Developments. Top. Curr. Chem. 2005, 258, 161–204. [Google Scholar] [CrossRef]
- Emami, S.; Tavangar, P.; Keighobadi, M. An Overview of Azoles Targeting Sterol 14α-Demethylase for Antileishmanial Therapy. Eur. J. Med. Chem. 2017, 135, 241–259. [Google Scholar] [CrossRef]
- Menezes, T.M.; de Almeida, S.M.V.; de Moura, R.O.; Seabra, G.; de Lima, M.d.C.A.; Neves, J.L. Spiro-Acridine Inhibiting Tyrosinase Enzyme: Kinetic, Protein-Ligand Interaction and Molecular Docking Studies. Int. J. Biol. Macromol. 2019, 122, 289–297. [Google Scholar] [CrossRef]
- Turcano, L.; Torrente, E.; Missineo, A.; Andreini, M.; Gramiccia, M.; Di Muccio, T.; Genovese, I.; Fiorillo, A.; Harper, S.; Bresciani, A.; et al. Identification and Binding Mode of a Novel Leishmania Trypanothione Reductase Inhibitor from High Throughput Screening. PLoS Negl. Trop. Dis. 2018, 12, e0006969. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos Nascimento, I.J.; de Aquino, T.M.; da Silva-Júnior, E.F. Cruzain and Rhodesain Inhibitors: Last Decade of Advances in Seeking for New Compounds Against American and African Trypanosomiases. Curr. Top. Med. Chem. 2021, 21, 1871–1899. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.W.; Zhang, Y. DockRMSD: An Open-Source Tool for Atom Mapping and RMSD Calculation of Symmetric Molecules through Graph Isomorphism. J. Cheminform. 2019, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Verma, R.K.; Prajapati, V.K.; Verma, G.K.; Chakraborty, D.; Sundar, S.; Rai, M.; Dubey, V.K.; Singh, M.S. Molecular Docking and In Vitro Antileishmanial Evaluation of Chromene-2-Thione Analogues. ACS Med. Chem. Lett. 2012, 3, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Prada, C.F.; Álvarez-Velilla, R.; Balaña-Fouce, R.; Prieto, C.; Calvo-Álvarez, E.; Escudero-Martínez, J.M.; Requena, J.M.; Ordóñez, C.; Desideri, A.; Pérez-Pertejo, Y.; et al. Gimatecan and Other Camptothecin Derivatives Poison Leishmania DNA-Topoisomerase IB Leading to a Strong Leishmanicidal Effect. Biochem. Pharmacol. 2013, 85, 1433–1440. [Google Scholar] [CrossRef] [Green Version]
- Tejería, A.; Pérez-Pertejo, Y.; Reguera, R.M.; Balaña-Fouce, R.; Alonso, C.; Fuertes, M.; González, M.; Rubiales, G.; Palacios, F. Antileishmanial Effect of New Indeno-1,5-Naphthyridines, Selective Inhibitors of Leishmania Infantum Type IB DNA Topoisomerase. Eur. J. Med. Chem. 2016, 124, 740–749. [Google Scholar] [CrossRef]
- Mamidala, R.; Majumdar, P.; Jha, K.K.; Bathula, C.; Agarwal, R.; Chary, M.T.; Mazumdar, H.K.; Munshi, P.; Sen, S. Identification of Leishmania Donovani Topoisomerase 1 Inhibitors via Intuitive Scaffold Hopping and Bioisosteric Modification of Known Top 1 Inhibitors. Sci. Rep. 2016, 6, 26603. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, L.; Alves, K.F.; Maciel-Rezende, C.M.; Jesus, L.d.O.P.; Pires, F.R.; Junior, C.V.; Izidoro, M.A.; Júdice, W.A.d.S.; dos Santos, M.H.; Marques, M.J. Benzophenone Derivatives as Cysteine Protease Inhibitors and Biological Activity against Leishmania (L.) Amazonensis Amastigotes. Biomed. Pharmacother. 2015, 75, 93–99. [Google Scholar] [CrossRef]
- Dos Santos Nascimento, I.J.; da Silva-Júnior, E.F. TNF-α Inhibitors from Natural Compounds: An Overview, CADD Approaches, and Their Exploration for Anti-Inflammatory Agents. Comb. Chem. High Throughput Screen. 2022, 25, 2317–2340. [Google Scholar] [CrossRef]
- Dos Santos Nascimento, I.J.; de Aquino, T.M.; da Silva-Júnior, E.F. Molecular Docking and Dynamics Simulations Studies of a Dataset of NLRP3 Inflammasome Inhibitors. Recent Adv. Inflamm. Allergy Drug Discov. 2022, 15, 80–86. [Google Scholar] [CrossRef]
- Dos Santos Nascimento, I.J.; de Aquino, T.M.; da Silva-Júnior, E.F. Repurposing FDA-Approved Drugs Targeting SARS-CoV2 3CLpro: A Study by Applying Virtual Screening, Molecular Dynamics, MM-PBSA Calculations and Covalent Docking. Lett. Drug Des. Discov. 2022, 19, 637–653. [Google Scholar] [CrossRef]
- Acharya, P.T.; Bhavsar, Z.A.; Jethava, D.J.; Patel, D.B.; Patel, H.D. A Review on Development of Bio-Active Thiosemicarbazide Derivatives: Recent Advances. J. Mol. Struct. 2021, 1226, 129268. [Google Scholar] [CrossRef]
- Gazieva, G.A.; Kravchenko, A.N. Thiosemicarbazides in the Synthesis of Five- and Six-Membered Heterocyclic Compounds. Russ. Chem. Rev. 2012, 81, 494–523. [Google Scholar] [CrossRef]
- Taha, M.; Ismail, N.H.; Ali, M.; Rashid, U.; Imran, S.; Uddin, N.; Khan, K.M. Molecular Hybridization Conceded Exceptionally Potent Quinolinyl-Oxadiazole Hybrids through Phenyl Linked Thiosemicarbazide Antileishmanial Scaffolds: In Silico Validation and SAR Studies. Bioorg. Chem. 2017, 71, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.P.; Carneiro, Z.A.; Prado, F.S.; Souza, J.R.; Furlan e Silva, L.H.; Oliveira, C.G.; Deflon, V.M.; de Albuquerque, S.; Leite, N.B.; Machado, A.E.H.; et al. Cu(I) Complexes with Thiosemicarbazides Derived from p-Toluenesulfohydrazide: Structural, Luminescence and Biological Studies. Polyhedron 2018, 155, 170–179. [Google Scholar] [CrossRef]
- Leite, A.C.L.; de Lima, R.S.; Moreira, D.R.d.M.; Cardoso, M.V.d.O.; Gouveia de Brito, A.C.; Farias dos Santos, L.M.; Hernandes, M.Z.; Kiperstok, A.C.; de Lima, R.S.; Soares, M.B.P. Synthesis, Docking, and In Vitro Activity of Thiosemicarbazones, Aminoacyl-Thiosemicarbazides and Acyl-Thiazolidones against Trypanosoma Cruzi. Bioorg. Med. Chem. 2006, 14, 3749–3757. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, S.M.V.; Lafayette, E.A.; Silva, W.L.; de Lima Serafim, V.; Menezes, T.M.; Neves, J.L.; Ruiz, A.L.T.G.; de Carvalho, J.E.; de Moura, R.O.; Beltrão, E.I.C.; et al. New Spiro-Acridines: DNA Interaction, Antiproliferative Activity and Inhibition of Human DNA Topoisomerases. Int. J. Biol. Macromol. 2016, 92, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Janočková, J.; Plšíková, J.; Kašpárková, J.; Brabec, V.; Jendželovský, R.; Mikeš, J.; Kova̘, J.; Hamu̘aková, S.; Fedoročko, P.; Kuča, K.; et al. Inhibition of DNA Topoisomerases i and II and Growth Inhibition of HL-60 Cells by Novel Acridine-Based Compounds. Eur. J. Pharm. Sci. 2015, 76, 192–202. [Google Scholar] [CrossRef]
- Kava, H.W.; Murray, V. CpG Methylation Increases the DNA Binding of 9-Aminoacridine Carboxamide Pt Analogues. Bioorg. Med. Chem. 2016, 24, 4701–4710. [Google Scholar] [CrossRef]
- Singh, P.; Kumar, A.; Sharma, A.; Kaur, G. Identification of Amino Acid Appended Acridines as Potential Leads to Anti-Cancer Drugs. Bioorg. Med. Chem. Lett. 2015, 25, 3854–3858. [Google Scholar] [CrossRef]
- Mitra, P.; Chakraborty, P.K.; Saha, P.; Ray, P.; Basu, S. Antibacterial Efficacy of Acridine Derivatives Conjugated with Gold Nanoparticles. Int. J. Pharm. 2014, 473, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Palaniraja, J.; Kumar, S.S.; Ramki, S.; Arunachalam, P.; Roopan, S.M. Conventional Spectroscopic Identification of Biologically Active Imidazo-Pyrimido Fused Acridines: In Vitro Anti-Bacterial and Anti-Feedant Activity. J. Mol. Liq. 2017, 230, 634–640. [Google Scholar] [CrossRef]
- De Silva, R.T.; Perera, R.P.; Nanayakkara, C.M. Design, Synthesis and Antibacterial Activity Evaluation of 9-Phenyl-10-(2-Phenylalkyl) Acridinium Bromide: A Novel Acridine Based Antibacterial Agent. Curr. Trends Biotechnol. Pharm. 2014, 8, 114–123. [Google Scholar]
- Chtita, S.; Ghamali, M.; Hmamouchi, R.; Elidrissi, B.; Bourass, M.; Larif, M.; Bouachrine, M.; Lakhlifi, T. Investigation of Antileishmanial Activities of Acridines Derivatives against Promastigotes and Amastigotes Form of Parasites Using Quantitative Structure Activity Relationship Analysis. Adv. Phys. Chem. 2016, 2016, 5137289. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Calienes Valdés, A. Acridine and Acridinones: Old and New Structures with Antimalarial Activity. Open Med. Chem. J. 2011, 5, 11–20. [Google Scholar] [CrossRef]
- Dostálová, A.; Volf, P. Leishmania Development in Sand Flies: Parasite-Vector Interactions Overview. Parasites Vectors 2012, 5, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, T.F.; Tomiotto-Pellissier, F.; Pasquali, A.K.S.; Pinto-Ferreira, F.; Pavanelli, W.R.; Conchon-Costa, I.; Navarro, I.T.; Caldart, E.T. Phenotypical and Genotypical Differences among Leishmania (Leishmania) Amazonensis Isolates That Caused Different Clinical Frames in Humans and Dogs: A Systematic Review. Acta Trop. 2021, 221, 106018. [Google Scholar] [CrossRef] [PubMed]
- De Menezes, J.P.; Saraiva, E.M.; Da Rocha-Azevedo, B. The Site of the Bite: Leishmania Interaction with Macrophages, Neutrophils and the Extracellular Matrix in the Dermis. Parasites Vectors 2016, 9, 264. [Google Scholar] [CrossRef] [Green Version]
- Santos-Mateus, D.; Passero, F.; Rodrigues, A.; Pereira, M.D.A. The Battle between Leishmania and the Host Immune System at a Glance. Int. Trends Immun. 2016, 4, 28–34. [Google Scholar] [CrossRef]
- Gao, C.; Liu, F.; Luan, X.; Tan, C.; Liu, H.; Xie, Y.; Jin, Y.; Jiang, Y. Novel Synthetic 2-Amino-10-(3,5-Dimethoxy)Benzyl-9(10H)-Acridinone Derivatives as Potent DNA-Binding Antiproliferative Agents. Bioorg. Med. Chem. 2010, 18, 7507–7514. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, J.; Wang, M.Q.; Zhang, D.W.; Lu, Q.-S.; Huang, Y.; Lin, H.H.; Yu, X.Q. Synthesis, DNA Binding and Cleavage Activity of Macrocyclic Polyamines Bearing Mono- or Bis-Acridine Moieties. Eur. J. Med. Chem. 2010, 45, 5302–5308. [Google Scholar] [CrossRef] [PubMed]
- Janovec, L.; Kožurková, M.; Sabolová, D.; Ungvarský, J.; Paulíková, H.; Plšíková, J.; Vantová, Z.; Imrich, J. Cytotoxic 3,6-Bis((Imidazolidinone)Imino)Acridines: Synthesis, DNA Binding and Molecular Modeling. Bioorg. Med. Chem. 2011, 19, 1790–1801. [Google Scholar] [CrossRef] [PubMed]
- Aleksić, M.M.; Kapetanović, V. An Overview of the Optical and Electrochemical Methods for Detection of DNA—Drug Interactions. Acta Chim. Slov. 2014, 61, 555–573. [Google Scholar]
- Rescifina, A.; Zagni, C.; Varrica, M.G.; Pistarà, V.; Corsaro, A. Recent Advances in Small Organic Molecules as DNA Intercalating Agents: Synthesis, Activity, and Modeling. Eur. J. Med. Chem. 2014, 74, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Sirajuddin, M.; Ali, S.; Badshah, A. Drug-DNA Interactions and Their Study by UV-Visible, Fluorescence Spectroscopies and Cyclic Voltametry. J. Photochem. Photobiol. B Biol. 2013, 124, 1–19. [Google Scholar] [CrossRef]
- Lang, X.; Li, L.; Chen, Y.; Sun, Q.; Wu, Q.; Liu, F.; Tan, C.; Liu, H.; Gao, C.; Jiang, Y. Novel Synthetic Acridine Derivatives as Potent DNA-Binding and Apoptosis-Inducing Antitumor Agents. Bioorg. Med. Chem. 2013, 21, 4170–4177. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, W.; Ma, R.; Chen, S.; Ren, S.; Jiang, T. Design, Synthesis and DNA Interaction Study of New Potential DNA Bis-Intercalators Based on Glucuronic Acid. Int. J. Mol. Sci. 2013, 14, 16851–16865. [Google Scholar] [CrossRef] [PubMed]
- Nepali, K.; Sharma, S.; Sharma, M.; Bedi, P.M.S.; Dhar, K.L. Rational Approaches, Design Strategies, Structure Activity Relationship and Mechanistic Insights for Anticancer Hybrids. Eur. J. Med. Chem. 2014, 77, 422–487. [Google Scholar] [CrossRef]
- Shi, J.H.; Liu, T.T.; Jiang, M.; Chen, J.; Wang, Q. Characterization of Interaction of Calf Thymus DNA with Gefitinib: Spectroscopic Methods and Molecular Docking. J. Photochem. Photobiol. B Biol. 2015, 147, 47–55. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, L.; Dong, Y.; Li, J.; Liu, X. Spectroscopic and Molecular Modeling Methods to Investigate the Interaction between 5-Hydroxymethyl-2-Furfural and Calf Thymus DNA Using Ethidium Bromide as a Probe. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2014, 124, 78–83. [Google Scholar] [CrossRef]
- Li, M.; Liu, Y. Topoisomerase I in Human Disease Pathogenesis and Treatments. Genom. Proteom. Bioinform. 2016, 14, 166–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.R.; Guimarães, A.S.; do Nascimento, J.; do Santos Nascimento, I.J.; da Silva, E.B.; McKerrow, J.H.; Cardoso, S.H.; da Silva-Júnior, E.F. Computer-Aided Design of 1,4-Naphthoquinone-Based Inhibitors Targeting Cruzain and Rhodesain Cysteine Proteases. Bioorg. Med. Chem. 2021, 41, 116213. [Google Scholar] [CrossRef] [PubMed]
- Mottram, J.C.; Brooks, D.R.; Coombs, G.H. Roles of Cysteine Proteinases of Trypanosomes and Leishmania in Host-Parasite Interactions. Curr. Opin. Microbiol. 1998, 1, 455–460. [Google Scholar] [CrossRef]
- McKerrow, J. Cysteine Protease Inhibitors as Chemotherapy for Parasitic Infections. Bioorg. Med. Chem. 1999, 7, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Federico, L.B.; Silva, G.M.; Gomes, S.Q.; Francischini, I.A.G.; Barcelos, M.P.; dos Santos, C.B.R.; Costa, L.T.; Campos Rosa, J.M.; de Paula da Silva, C.H.T. Potential Colchicine Binding Site Inhibitors Unraveled by Virtual Screening, Molecular Dynamics and MM/PBSA. Comput. Biol. Med. 2021, 137, 104817. [Google Scholar] [CrossRef]
- Singh, S.; Prajapati, V.K. Exploring Actinomycetes Natural Products to Identify Potential Multi-Target Inhibitors against Leishmania Donovani. 3 Biotech 2022, 12, 235. [Google Scholar] [CrossRef]
- Eberle, R.J.; Olivier, D.S.; Amaral, M.S.; Gering, I.; Willbold, D.; Arni, R.K.; Coronado, M.A. The Repurposed Drugs Suramin and Quinacrine Cooperatively Inhibit SARS-CoV-2 3CLpro In Vitro. Viruses 2021, 13, 873. [Google Scholar] [CrossRef]
- Kückelhaus, C.S.; Kückelhaus, S.A.S.; Tosta, C.E.; Muniz-Junqueira, M.I. Pravastatin Modulates Macrophage Functions of Leishmania (L.) Amazonensis-Infected BALB/c Mice. Exp. Parasitol. 2013, 134, 18–25. [Google Scholar] [CrossRef] [PubMed]
- De Araújo-Vilges, K.M.; de Oliveira, S.V.; Couto, S.C.P.; Fokoue, H.H.; Romero, G.A.S.; Kato, M.J.; Romeiro, L.A.S.; Leite, J.R.S.A.; Kuckelhaus, S.A.S. Effect of Piplartine and Cinnamides on Leishmania Amazonensis, Plasmodium Falciparum and on Peritoneal Cells of Swiss Mice. Pharm. Biol. 2017, 55, 1601–1607. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Deng, G.; Wang, X.; Luo, J.; Qian, X.; Ling, W. Cyanidin-3-O-β-Glucoside Polarizes LPS-Induced M1 into M2 Macrophage in J774 Cells via PPARγ-Mediated NF-ΚB and STAT6 Signaling Pathway. J. Funct. Foods 2021, 77, 104314. [Google Scholar] [CrossRef]
- Rodrigues, K.A.D.F.; Dias, C.N.D.S.; Neris, P.L.D.N.; Rocha, J.D.C.; Scotti, M.T.; Scotti, L.; Mascarenhas, S.R.; Veras, R.C.; De Medeiros, I.A.; Keesen, T.D.S.L.; et al. 2-Amino-Thiophene Derivatives Present Antileishmanial Activity Mediated by Apoptosis and Immunomodulation in Vitro. Eur. J. Med. Chem. 2015, 106, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz, A.C.N.; Brondani, D.J.; de Santana, T.I.; da Silva, L.O.; Borba, E.F.d.O.; de Faria, A.R.; de Albuquerque, J.F.C.; Piessard, S.; Ximenes, R.M.; Baratte, B.; et al. Biological Evaluation of Arylsemicarbazone Derivatives as Potential Anticancer Agents. Pharmaceuticals 2019, 12, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharova, O.; Nevinsky, G.; Politanskaya, L.; Baev, D.; Ovchinnikova, L.; Tretyakov, E. Evaluation of Antioxidant Activity and Cytotoxicity of Polyfluorinated Diarylacetylenes and Indoles toward Human Cancer Cells. J. Fluor. Chem. 2019, 226, 109353. [Google Scholar] [CrossRef]
- Hawash, M.; Qneibi, M.; Jaradat, N.; Abualhasan, M.; Amer, J.; Amer, E.H.; Ibraheem, T.; Hindieh, S.; Tarazi, S.; Sobuh, S. The Impact of Filtered Water-Pipe Smoke on Healthy versus Cancer Cells and Their Neurodegenerative Role on AMPA Receptor. Drug Chem. Toxicol. 2022, 45, 2292–2300. [Google Scholar] [CrossRef] [PubMed]
- Kückelhaus, S.A.S.; de Aquino, D.S.; Borges, T.K.; Moreira, D.C.; Leite, L.d.M.; Muniz-Junqueira, M.I.; Kückelhaus, C.S.; Sierra Romero, G.A.; Prates, M.V.; Bloch, C.; et al. Phylloseptin-1 Is Leishmanicidal for Amastigotes of Leishmania Amazonensis inside Infected Macrophages. Int. J. Environ. Res. Public Health 2020, 17, 4856. [Google Scholar] [CrossRef]
- Wolfe, A.; Shimer, G.H.; Meehan, T. Polycyclic Aromatic Hydrocarbons Physically Intercalate into Duplex Regions of Denatured DNA. Biochemistry 1987, 26, 6392–6396. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, R.G.; Ribeiro, A.G.; Segundo, M.Â.S.P.; de Oliveira, J.F.; de Lima, M.d.C.A.; de Lima Souza, T.R.C.; de Almeida, S.M.V.; de Moura, R.O. Synthesis, DNA and Protein Interactions and Human Topoisomerase Inhibition of Novel Spiroacridine Derivatives. Bioorg. Med. Chem. 2018, 26, 5911–5921. [Google Scholar] [CrossRef] [PubMed]
- Kückelhaus, S.A.S.; Leite, J.R.S.A.; Muniz-Junqueira, M.I.; Sampaio, R.N.; Bloch, C.; Tosta, C.E. Antiplasmodial and Antileishmanial Activities of Phylloseptin-1, an Antimicrobial Peptide from the Skin Secretion of Phyllomedusa Azurea (Amphibia). Exp. Parasitol. 2009, 123, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Schröder, J.; Noack, S.; Marhöfer, R.J.; Mottram, J.C.; Coombs, G.H.; Selzer, P.M. Identification of Semicarbazones, Thiosemicarbazones and Triazine Nitriles as Inhibitors of Leishmania Mexicana Cysteine Protease CPB. PLoS ONE 2013, 8, e77460. [Google Scholar] [CrossRef] [Green Version]
- Freitas, P.; Castilho, T.; de Almeida, L.; Maciel-Rezende, C.; Costa, L.; Viegas Junior, C.; Marques, M.; dos Santos, M.; da Silveira, N. An in Silico Study of Benzophenone Derivatives as Potential Non-Competitive Inhibitors of Trypanosoma Cruzi and Leishmania Amazonensis Cysteine Proteinases. J. Braz. Chem. Soc. 2017, 29, 515–527. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI Reference Sequences (RefSeq): A Curated Non-Redundant Sequence Database of Genomes, Transcripts and Proteins. Nucleic Acids Res. 2007, 35, D61–D65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillmor, S.A.; Craik, C.S.; Fletterick, R.J. Structural Determinants of Specificity in the Cysteine Protease Cruzain. Protein Sci. 1997, 6, 1603–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwede, T. SWISS-MODEL: An Automated Protein Homology-Modeling Server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Hawash, M.; Qaoud, M.T.; Jaradat, N.; Abdallah, S.; Issa, S.; Adnan, N.; Hoshya, M.; Sobuh, S.; Hawash, Z. Anticancer Activity of Thiophene Carboxamide Derivatives as CA-4 Biomimetics: Synthesis, Biological Potency, 3D Spheroid Model, and Molecular Dynamics Simulation. Biomimetics 2022, 7, 247. [Google Scholar] [CrossRef]
- Canals, A.; Purciolas, M.; Aymamí, J.; Coll, M. The Anticancer Agent Ellipticine Unwinds DNA by Intercalative Binding in an Orientation Parallel to Base Pairs. Acta Crystallogr. Sect. D Biol. Crystallogr. 2005, 61, 1009–1012. [Google Scholar] [CrossRef] [Green Version]
- Baiocco, P.; Poce, G.; Alfonso, S.; Cocozza, M.; Porretta, G.C.; Colotti, G.; Biava, M.; Moraca, F.; Botta, M.; Yardley, V.; et al. Inhibition of Leishmania Infantum Trypanothione Reductase by Azole-Based Compounds: A Comparative Analysis with Its Physiological Substrate by X-ray Crystallography. ChemMedChem 2013, 8, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Hargrove, T.Y.; Wawrzak, Z.; Liu, J.; Nes, W.D.; Waterman, M.R.; Lepesheva, G.I. Substrate Preferences and Catalytic Parameters Determined by Structural Characteristics of Sterol 14α-Demethylase (CYP51) from Leishmania Infantum. J. Biol. Chem. 2011, 286, 26838–26848. [Google Scholar] [CrossRef] [Green Version]
- Davies, D.R.; Mushtaq, A.; Interthal, H.; Champoux, J.J.; Hol, W.G.J. The Structure of the Transition State of the Heterodimeric Topoisomerase I of Leishmania Donovani as a Vanadate Complex with Nicked DNA. J. Mol. Biol. 2006, 357, 1202–1210. [Google Scholar] [CrossRef]
- Tschoeke, D.A.; Nunes, G.L.; Jardim, R.; Lima, J.; Dumaresq, A.S.R.; Gomes, M.R.; Pereira, L.d.M.; Loureiro, D.R.; Stoco, P.H.; de Matos Guedes, H.L.; et al. The Comparative Genomics and Phylogenomics of Leishmania Amazonensis Parasite. Evol. Bioinform. 2014, 10, 131–153. [Google Scholar] [CrossRef]
- De Almeida, P.S.V.B.; de Arruda, H.J.; Sousa, G.L.S.; Ribeiro, F.V.; de Azevedo-França, J.A.; Ferreira, L.A.; Guedes, G.P.; Silva, H.; Kummerle, A.E.; Neves, A.P. Cytotoxicity Evaluation and DNA Interaction of RuII-Bipy Complexes Containing Coumarin-Based Ligands. Dalt. Trans. 2021, 50, 14908–14919. [Google Scholar] [CrossRef] [PubMed]
- Zoete, V.; Cuendet, M.A.; Grosdidier, A.; Michielin, O. SwissParam: A Fast Force Field Generation Tool for Small Organic Molecules. J. Comput. Chem. 2011, 32, 2359–2368. [Google Scholar] [CrossRef]
- Roque Marques, K.M.; do Desterro, M.R.; de Arruda, S.M.; de Araújo Neto, L.N.; do Carmo Alves de Lima, M.; de Almeida, S.M.V.; da Silva, E.C.D.; de Aquino, T.M.; da Silva-Júnior, E.F.; de Araújo-Júnior, J.X.; et al. 5-Nitro-Thiophene-Thiosemicarbazone Derivatives Present Antitumor Activity Mediated by Apoptosis and DNA Intercalation. Curr. Top. Med. Chem. 2019, 19, 1075–1091. [Google Scholar] [CrossRef] [PubMed]
- Silva-Junior, E.F.; Barcellos Franca, P.H.; Quintans-Junior, L.J.; Mendonca-Junior, F.J.B.; Scotti, L.; Scotti, M.T.; de Aquino, T.M.; de Araujo-Junior, J.X. Dynamic Simulation, Docking and DFT Studies Applied to a Set of Anti-Acetylcholinesterase Inhibitors in the Enzyme β-Secretase (BACE-1): An Important Therapeutic Target in Alzheimer’s Disease. Curr. Comput. Aided Drug Des. 2017, 13, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Braga, T.C.; Silva, T.F.; Maciel, T.M.S.; da Silva, E.C.D.; da Silva-Júnior, E.F.; Modolo, L.V.; Figueiredo, I.M.; Santos, J.C.C.; de Aquino, T.M.; de Fátima, Â. Ionic Liquid-Assisted Synthesis of Dihydropyrimidin(Thi)One Biginelli Adducts and Investigation of Their Mechanism of Urease Inhibition. New J. Chem. 2019, 43, 15187–15200. [Google Scholar] [CrossRef]
- Kumari, R.; Kumar, R.; Lynn, A. G_mmpbsa—A GROMACS Tool for High-Throughput MM-PBSA Calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef] [PubMed]
- Sarma, H.; Mattaparthi, V.S.K. Structure-Based Virtual Screening of High-Affinity ATP-Competitive Inhibitors Against Human Lemur Tyrosine Kinase-3 (LMTK3) Domain: A Novel Therapeutic Target for Breast Cancer. Interdiscip. Sci. Comput. Life Sci. 2019, 11, 527–541. [Google Scholar] [CrossRef]
- Wang, F.; Wu, F.-X.; Li, C.-Z.; Jia, C.-Y.; Su, S.-W.; Hao, G.-F.; Yang, G.-F. ACID: A Free Tool for Drug Repurposing Using Consensus Inverse Docking Strategy. J. Cheminform. 2019, 11, 73. [Google Scholar] [CrossRef] [PubMed]









| COMPOUND | IC50AMA (µg mL−1) | IC50PRO (µg mL−1) | CC50 (µg mL−1) | EC50 (µg mL−1) | SI (CC50/IC50AMA) |
|---|---|---|---|---|---|
| ACW-02 | 6.57 ± 0.04 | 94.97 ± 0.06 | >256.00 | 9.46 ± 0.06 | >38.94 |
| AMPHOTERICIN B | 2.94 ± 0.07 | 1.41 ± 0.003 | 3.09 ± 0.01 | 2.80 ± 0.0093 | 1.05 |
| ACW-2 (µg mL−1) | Infected Cells (%) | Mean of Leishmanias | Infection Rate (%) |
|---|---|---|---|
| 0 | 22.00 | 2.40 | 53.00 |
| 1 | 12.00 | 1.20 | 14.50 |
| 16 | 11.50 | 1.20 | 13.50 |
| 32 | 11.50 | 1.80 | 21.00 |
| Compound | λmax Free (nm) | λmax Bound (nm) | ∆λ (nm) | Hyperchr a (%) | Hypochr b (%) | Kb (M−1) |
|---|---|---|---|---|---|---|
| ACW-02 + DNA | 460 | 461 | 1 | 47.53 | - | 1.17 × 106 |
| 2B9S (Topo I) | CPBLa | |
|---|---|---|
| ΔGbing (KJ/mol) | −42.17 ± 21.36 | −103.13 ± 8.11 |
| SASA Energy (KJ/mol) | −10.66 ± 1.69 | −13.15 ± 0.78 |
| Polar solvation energy (KJ/mol) | 65.34 ± 23.63 | 61.09 ± 5.20 |
| Electrostatic energy (KJ/mol) | −1.87 ± 19.59 | 0.68 ± 3.60 |
| Van der Waals energy (KJ/mol) | −94.98 ± 25.52 | −151.75 ± 7.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albino, S.L.; da Silva Moura, W.C.; Reis, M.M.L.d.; Sousa, G.L.S.; da Silva, P.R.; de Oliveira, M.G.C.; Borges, T.K.d.S.; Albuquerque, L.F.F.; de Almeida, S.M.V.; de Lima, M.d.C.A.; et al. ACW-02 an Acridine Triazolidine Derivative Presents Antileishmanial Activity Mediated by DNA Interaction and Immunomodulation. Pharmaceuticals 2023, 16, 204. https://doi.org/10.3390/ph16020204
Albino SL, da Silva Moura WC, Reis MMLd, Sousa GLS, da Silva PR, de Oliveira MGC, Borges TKdS, Albuquerque LFF, de Almeida SMV, de Lima MdCA, et al. ACW-02 an Acridine Triazolidine Derivative Presents Antileishmanial Activity Mediated by DNA Interaction and Immunomodulation. Pharmaceuticals. 2023; 16(2):204. https://doi.org/10.3390/ph16020204
Chicago/Turabian StyleAlbino, Sonaly Lima, Willian Charles da Silva Moura, Malu Maria Lucas dos Reis, Gleyton Leonel Silva Sousa, Pablo Rayff da Silva, Mayara Gabriele Carvalho de Oliveira, Tatiana Karla dos Santos Borges, Lucas Fraga Friaça Albuquerque, Sinara Mônica Vitalino de Almeida, Maria do Carmo Alves de Lima, and et al. 2023. "ACW-02 an Acridine Triazolidine Derivative Presents Antileishmanial Activity Mediated by DNA Interaction and Immunomodulation" Pharmaceuticals 16, no. 2: 204. https://doi.org/10.3390/ph16020204
APA StyleAlbino, S. L., da Silva Moura, W. C., Reis, M. M. L. d., Sousa, G. L. S., da Silva, P. R., de Oliveira, M. G. C., Borges, T. K. d. S., Albuquerque, L. F. F., de Almeida, S. M. V., de Lima, M. d. C. A., Kuckelhaus, S. A. S., Nascimento, I. J. d. S., Junior, F. J. B. M., da Silva, T. G., & de Moura, R. O. (2023). ACW-02 an Acridine Triazolidine Derivative Presents Antileishmanial Activity Mediated by DNA Interaction and Immunomodulation. Pharmaceuticals, 16(2), 204. https://doi.org/10.3390/ph16020204