

Acenocoumarol, an Anticoagulant Drug, Prevents Melanogenesis in B16F10 Melanoma Cells


Abstract
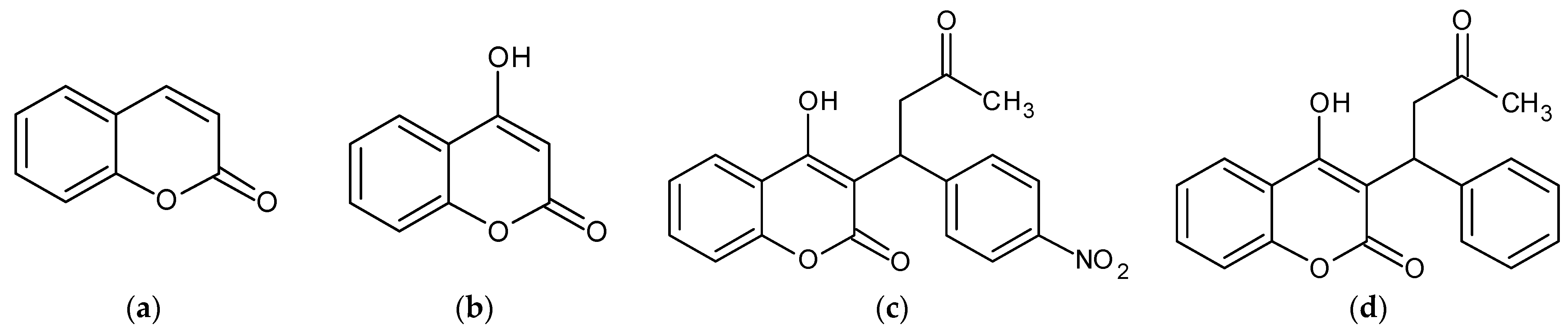
:1. Introduction
2. Results
2.1. Acenocoumarol and Warfarin Reduced Melanin Synthesis and Tyrosinase Activity in B16F10 Cells
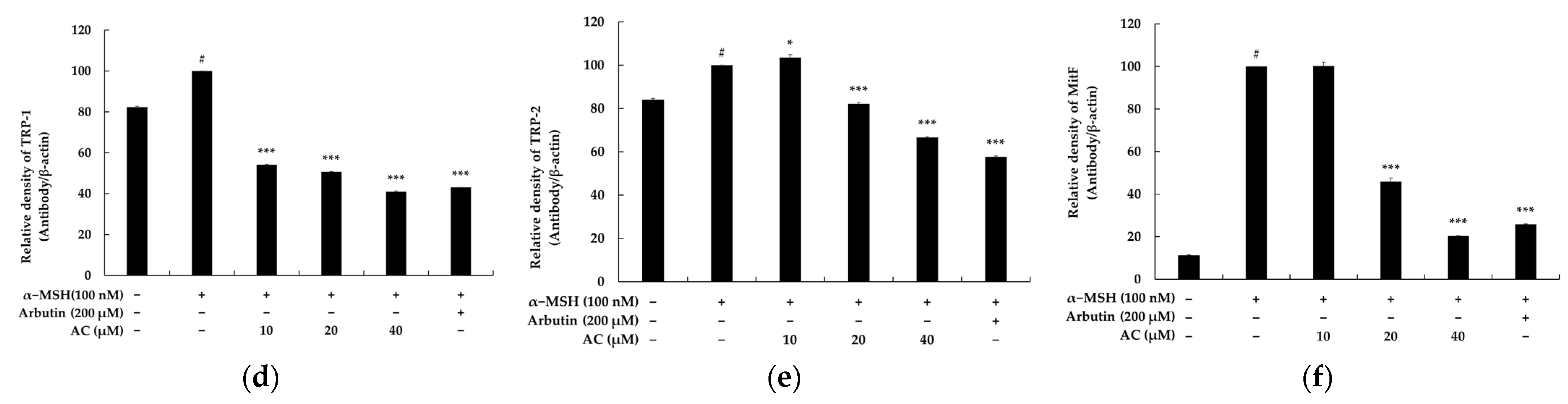
2.2. Acenocoumarol Regulated the Expression of Melanogenic Proteins
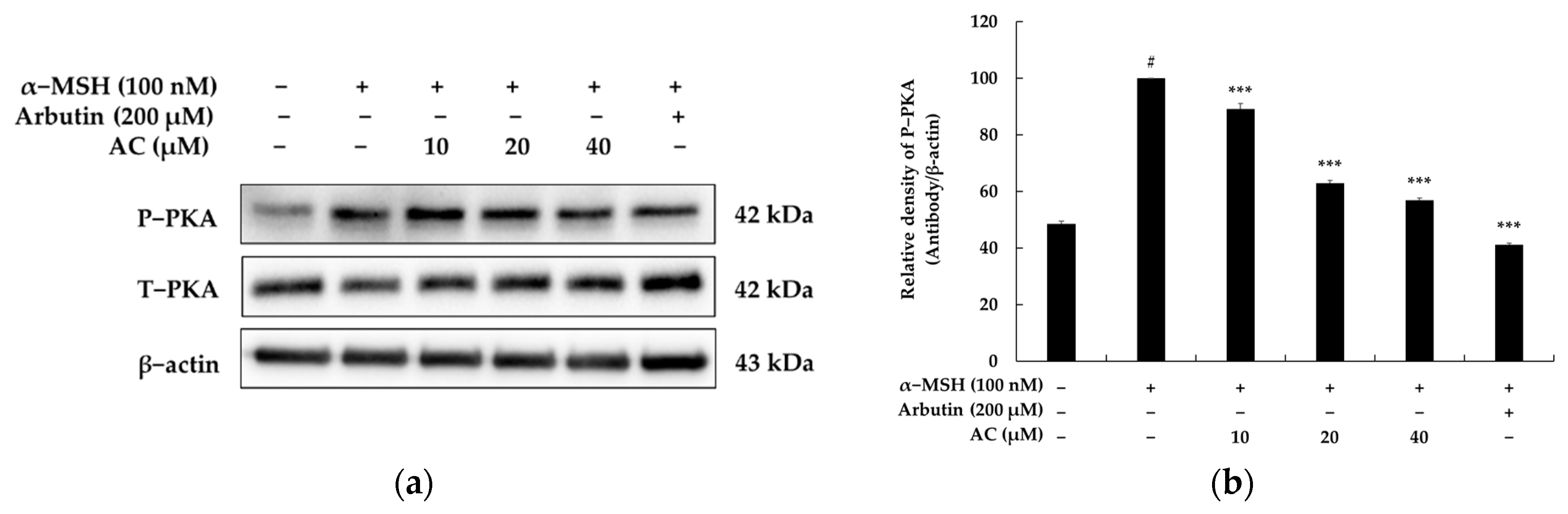
2.3. Acenocoumarol Inhibited Melanogenesis through the PKA Signaling Pathway
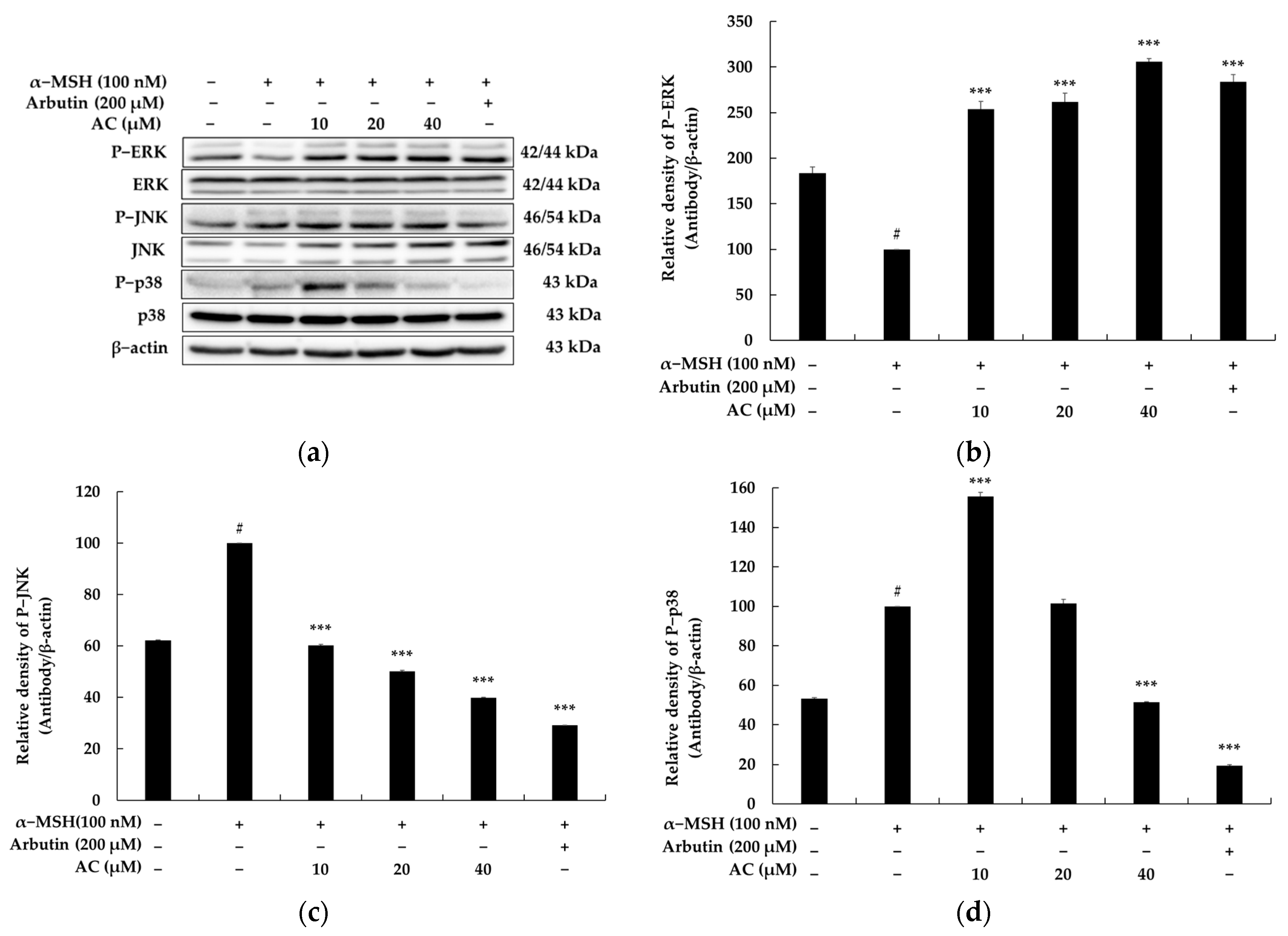
2.4. Acenocoumarol Suppressed Melanogenesis via the MAPK Signaling Pathway
2.5. Acenocoumarol Repressed Melanogenesis through PI3K/Akt/GSK-3β Signaling Pathways
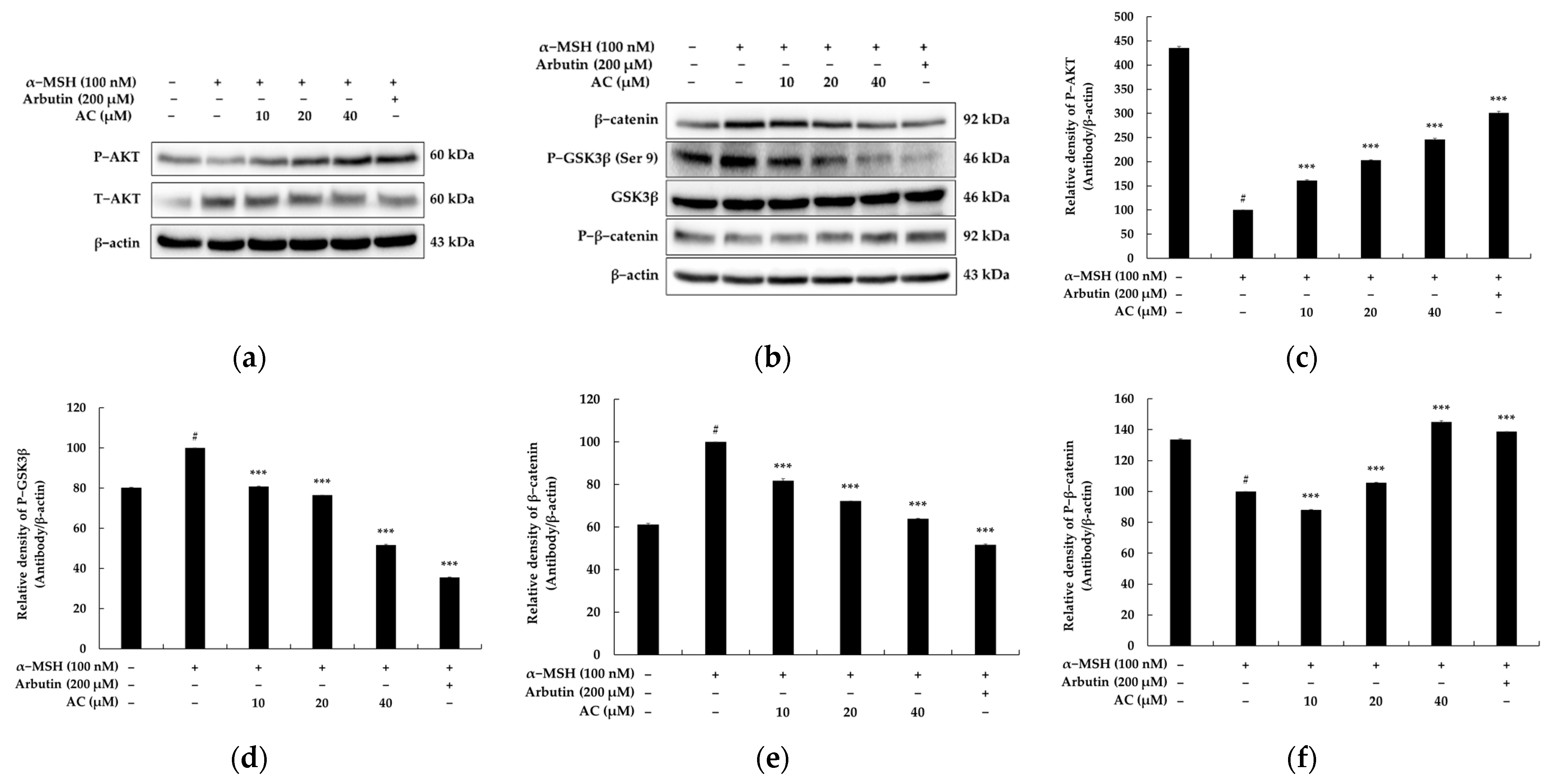
2.6. Acenocoumarol Repressed Melanogenesis through Wnt/β-Catenin Signaling Pathways
2.7. Acenocoumarol Was Found to Be a Safer Ingredient though a Human Primary Irritation Test
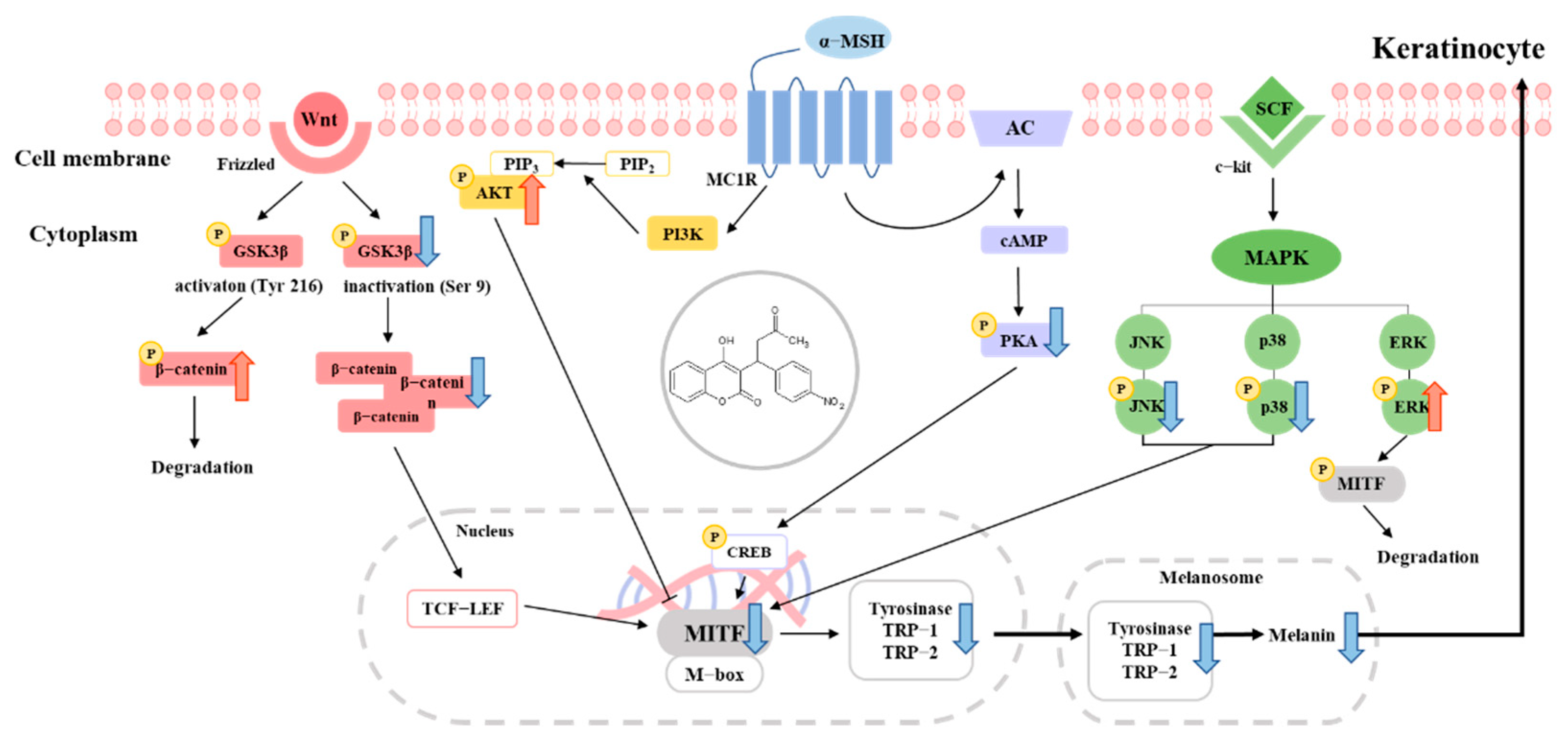
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. MTT Assay
4.4. Measurement of Melanin Contents
4.5. Measurement of Tyrosinase Activity
4.6. Western Blot Analysis
4.7. Primary Skin Irritation Test
4.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, G.; Dong, D.; Dong, Z.; Zhang, Q.; Fang, H.; Wang, C.; Zhang, S.; Wu, S.; Dong, Y.; Wan, Y. Drug repositioning: A bibliometric analysis. Front. Pharmacol. 2022, 13, 974849. [Google Scholar] [CrossRef]
- Chaffey, L.; Roberti, A.; Greaves, D.R. Drug repurposing in cardiovascular inflammation: Successes, failures, and future opportunities. Front. Pharmacol. 2022, 13, 1046406. [Google Scholar] [CrossRef]
- Burchett, J.R.; Dailey, J.M.; Kee, S.A.; Pryor, D.T.; Kotha, A.; Kankaria, R.A.; Straus, D.B.; Ryan, J.J. Targeting Mast Cells in Allergic Disease: Current Therapies and Drug Repurposing. Cells 2022, 11, 3031. [Google Scholar] [CrossRef] [PubMed]
- Maghsoudi, S.; Taghavi Shahraki, B.; Rameh, F.; Nazarabi, M.; Fatahi, Y.; Akhavan, O.; Rabiee, M.; Mostafavi, E.; Lima, E.C.; Saeb, M.R.; et al. A review on computer-aided chemogenomics and drug repositioning for rational COVID-19 drug discovery. Chem. Biol. Drug Des. 2022, 100, 699–721. [Google Scholar] [CrossRef]
- A Sahoo, B.M.; Ravi Kumar, B.V.V.; Sruti, J.; Mahapatra, M.K.; Banik, B.K.; Borah, P. Drug Repurposing Strategy (DRS): Emerging Approach to Identify Potential Therapeutics for Treatment of Novel Coronavirus Infection. Front. Mol. Biosci. 2021, 8, 628144. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Solanki, K.; Are, R.; Lavrijsen, M.; Pan, Q.; Baig, M.S.; Li, P. Nirmatrelvir exerts distinct antiviral potency against different human coronaviruses. Antivir. Res. 2023, 211, 105555. [Google Scholar] [CrossRef]
- Gray, E.J.; Nguyen-Van-Tam, J.S. Molnupiravir for SARS-CoV-2 infection: Public health and policy implications. J. Infect. 2023, 86, 121–122. [Google Scholar] [CrossRef] [PubMed]
- Kasperkiewicz, K.; Ponczek, M.B.; Owczarek, J.; Guga, P.; Budzisz, E. Antagonists of Vitamin K-Popular Coumarin Drugs and New Synthetic and Natural Coumarin Derivatives. Molecules 2020, 25, 1465. [Google Scholar] [CrossRef]
- Chen, X.; Jin, D.Y.; Stafford, D.W.; Tie, J.K. Evaluation of oral anticoagulants with vitamin K epoxide reductase in its native milieu. Blood 2018, 132, 1974–1984. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.Q.; Ye, Y.R. Predicting Glioma Cell Differentiation-inducing Drugs Using a Drug Repositioning Strategy. Comb. Chem. High Throughput Screen. 2023, 26, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Goka, B.; Klumpp, D.A. Synthesis of warfarin analogs: Conjugate addition reactions of alkenyl-substituted N-heterocycles with 4-hydroxycoumarin and related substrates. RSC Adv. 2023, 13, 4754–4756. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi Ziarani, G.; Moradi, R.; Ahmadi, T.; Gholamzadeh, P. The molecular diversity scope of 4-hydroxycoumarin in the synthesis of heterocyclic compounds via multicomponent reactions. Mol. Divers. 2019, 23, 1029–1064. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.B.; Zhao, Y.N.; Zhang, K.; Mack, P. Daphnetin, one of coumarin derivatives, is a protein kinase inhibitor. Biochem. Biophys. Res. Commun. 1999, 260, 682–685. [Google Scholar] [CrossRef]
- Han, H.J.; Hyun, C.G. Acenocoumarol Exerts Anti-Inflammatory Activity via the Suppression of NF-κB and MAPK Pathways in RAW 264.7 Cells. Molecules 2023, 28, 2075. [Google Scholar] [CrossRef]
- Abdou, M.M.; Rasha, A.; El-Saeed, R.A.; Bondock, S. Recent advances in 4-hydroxycoumarin chemistry. Part 1: Synthesis and reactions. Arab. J. Chem. 2019, 12, 88–121. [Google Scholar] [CrossRef]
- Trailokya, A.; Hiremath, J.S.; Sawhney, J.; Mishra, Y.K.; Kanhere, V.; Srinivasa, R.; Tiwaskar, M. Acenocoumarol: A Review of Anticoagulant Efficacy and Safety. J. Assoc. Physicians India 2016, 64, 88–93. [Google Scholar]
- Schroecksnadel, S.; Gostner, J.; Jenny, M.; Kurz, K.; Schennach, H.; Weiss, G.; Fuchs, D. Immunomodulatory effects in vitro of vitamin K antagonist acenocoumarol. Thromb. Res. 2013, 131, e264–e269. [Google Scholar] [CrossRef]
- Lee, M.S.; Chung, Y.C.; Moon, S.H.; Hyun, C.G. Lincomycin induces melanogenesis through the activation of MITF via p38 MAPK, AKT, and PKA signaling pathways. J. Appl. Biol. Chem. 2021, 64, 323–331. [Google Scholar] [CrossRef]
- Choi, H.; Yoon, J.H.; Youn, K.; Jun, M. Decursin prevents melanogenesis by suppressing MITF expression through the regulation of PKA/CREB, MAPKs, and PI3K/Akt/GSK-3β cascades. Biomed. Pharmacother. 2022, 147, 112651. [Google Scholar] [CrossRef]
- Uto, T.; Ohta, T.; Katayama, K.; Shoyama, Y. Silibinin promotes melanogenesis through the PKA and p38 MAPK signaling pathways in melanoma cells. Biomed. Res. 2022, 43, 31–39. [Google Scholar] [CrossRef]
- Wang, Z.; Tang, H.; Yang, L.; Li, Y.; Wu, H. Tanshinone IIA, a melanogenic ingredient basis of Salvia miltiorrhiza Bunge. Dermatol. Sin. 2021, 39, 33–40. [Google Scholar]
- Zhou, S.; Sakamoto, K. Pyruvic acid/ethyl pyruvate inhibits melanogenesis in B16F10 melanoma cells through PI3K/AKT, GSK3β, and ROS-ERK signaling pathways. Genes Cells 2019, 24, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Yen, H.; Lu, J.Y.; Chang, T.M.; Hii, C.H. Theophylline enhances melanogenesis in B16F10 murine melanoma cells through the activation of the MEK 1/2, and Wnt/β-catenin signaling pathways. Food Chem. Toxicol. 2020, 137, 111165. [Google Scholar] [CrossRef] [PubMed]
- Germanas, J.P.; Unni, E.; Kim, K.; Germanas, T.Y. Toward New Depigmenting Agents through Repurposing Existing Drugs: Substituted Hydroxyquinolines as Melanogenesis Inhibitors. J. Investig. Dermatol. 2023, 143, 176–179. [Google Scholar] [CrossRef]
- Guo, N.; You, X.; Wu, Y.; Du, D.; Zhang, L.; Shang, Q.; Liu, W. Continuous Fluorometric Method for Determining the Monophenolase Activity of Tyrosinase on L-Tyrosine, through Quenching L-DOPA Fluorescence by Borate. Anal. Chem. 2020, 92, 5780–5786. [Google Scholar] [CrossRef]
- Kuo, T.C.; Wang, P.H.; Wang, Y.K.; Chang, C.I.; Chang, C.Y.; Tseng, Y.J. RSDB: A rare skin disease database to link drugs with potential drug targets for rare skin diseases. Sci. Data 2022, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, R.; Paul, A.; Raj, V.S.; Ojha, K.K.; Kumar, S.; Panda, A.K.; Chaurasia, A.; Yadav, M.K. A Drug Repurposing Approach to Identify Therapeutics by Screening Pathogen Box Exploiting SARS-CoV-2 Main Protease. Chem. Biodivers. 2023, 20, e202200600. [Google Scholar] [CrossRef]
- Ahmed, F.; Ho, S.G.; Samantasinghar, A.; Memon, F.H.; Rahim, C.S.A.; Soomro, A.M.; Pratibha Sunildutt, N.; Kim, K.H.; Choi, K.H. Drug repurposing in psoriasis, performed by reversal of disease-associated gene expression profiles. Comput. Struct. Biotechnol. J. 2022, 20, 6097–6107. [Google Scholar] [CrossRef]
- Kim, H.M.; Hyun, C.G. Miglitol, an Oral Antidiabetic Drug, Downregulates Melanogenesis in B16F10 Melanoma Cells through the PKA, MAPK, and GSK3β/β-Catenin Signaling Pathways. Molecules 2022, 28, 115. [Google Scholar] [CrossRef]
- Prasannanjaneyulu, V.; Nene, S.; Jain, H.; Nooreen, R.; Otavi, S.; Chitlangya, P.; Srivastava, S. Old drugs, new tricks: Emerging role of drug repurposing in the management of atopic dermatitis. Cytokine Growth Factor Rev. 2022, 65, 12–26. [Google Scholar] [CrossRef]
- Sotiropoulou, G.; Zingkou, E.; Pampalakis, G. Redirecting drug repositioning to discover innovative cosmeceuticals. Exp. Dermatol. 2021, 30, 628–644. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.C.; Lee, S.H. Protocatechuic Aldehyde Inhibits α-MSH-Induced Melanogenesis in B16F10 Melanoma Cells via PKA/CREB-Associated MITF Downregulation. Int. J. Mol. Sci. 2021, 22, 3861. [Google Scholar] [CrossRef]
- Yoon, J.H.; Youn, K.; Jun, M. Discovery of Pinostrobin as a Melanogenic Agent in cAMP/PKA and p38 MAPK Signaling Pathway. Nutrients 2022, 14, 3713. [Google Scholar] [CrossRef]
- Zhao, N.; Su, X.; Li, H.; Li, Z.; Wang, Y.; Chen, J.; Zhuang, W. Schisandrin B inhibits α-melanocyte-stimulating hormone-induced melanogenesis in B16F10 cells via downregulation of MAPK and CREB signaling pathways. Biosci. Biotechnol. Biochem. 2021, 85, 834–841. [Google Scholar] [CrossRef]
- Shin, S.; Ko, J.; Kim, M.; Song, N.; Park, K. Morin Induces Melanogenesis via Activation of MAPK Signaling Pathways in B16F10 Mouse Melanoma Cells. Molecules 2021, 26, 2150. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Dai, G.F.; Hsu, J.L.; Lin, J.J.; Wu, W.T.; Su, C.C.; Wu, Y.J. Antimelanogenesis Effect of Methyl Gallate through the Regulation of PI3K/Akt and MEK/ERK in B16F10 Melanoma Cells. Evid. Based Complement. Alternat. Med. 2022, 2022, 5092655. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Yotsumoto, H.; Tian, Y.; Sakamoto, K. α-Mangostin suppressed melanogenesis in B16F10 murine melanoma cells through GSK3β and ERK signaling pathway. Biochem. Biophys. Rep. 2021, 15, 100949. [Google Scholar] [CrossRef]
- Kim, T.; Hyun, C.G. Imperatorin Positively Regulates Melanogenesis through Signaling Pathways Involving PKA/CREB, ERK, AKT, and GSK3β/β-Catenin. Molecules 2022, 27, 6512. [Google Scholar] [CrossRef]
- Lee, Y.; Hyun, C.G. Mechanistic Insights into the Ameliorating Effect of Melanogenesis of Psoralen Derivatives in B16F10 Melanoma Cells. Molecules 2022, 27, 2613. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Test Sample | No. of Respondents | 20 min after Removal | 24 h after Removal | Reaction Grade (R) * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| +1 | +2 | +3 | +4 | +1 | +2 | +3 | +4 | 24 h | 48 h | Mean | |||
| 1 | Acenocoumarol n (20 μM) | 0 | - | - | - | - | 0 | - | - | - | 0 | 0 | 0 |
| 2 | Acenocoumarol (40 μM) | 1 | - | - | - | - | 1 | - | - | - | 0 | 1 | 0.4 |
| 3 | Squalene | 0 | - | - | - | - | - | - | - | - | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.; Hyun, C. Acenocoumarol, an Anticoagulant Drug, Prevents Melanogenesis in B16F10 Melanoma Cells. Pharmaceuticals 2023, 16, 604. https://doi.org/10.3390/ph16040604
Han H, Hyun C. Acenocoumarol, an Anticoagulant Drug, Prevents Melanogenesis in B16F10 Melanoma Cells. Pharmaceuticals. 2023; 16(4):604. https://doi.org/10.3390/ph16040604
Chicago/Turabian StyleHan, Hyunju, and Changgu Hyun. 2023. "Acenocoumarol, an Anticoagulant Drug, Prevents Melanogenesis in B16F10 Melanoma Cells" Pharmaceuticals 16, no. 4: 604. https://doi.org/10.3390/ph16040604
APA StyleHan, H., & Hyun, C. (2023). Acenocoumarol, an Anticoagulant Drug, Prevents Melanogenesis in B16F10 Melanoma Cells. Pharmaceuticals, 16(4), 604. https://doi.org/10.3390/ph16040604