
Isolation, Purification, and Structural Characterization of Polysaccharides from Codonopsis pilosula and Their Anti-Tumor Bioactivity by Immunomodulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
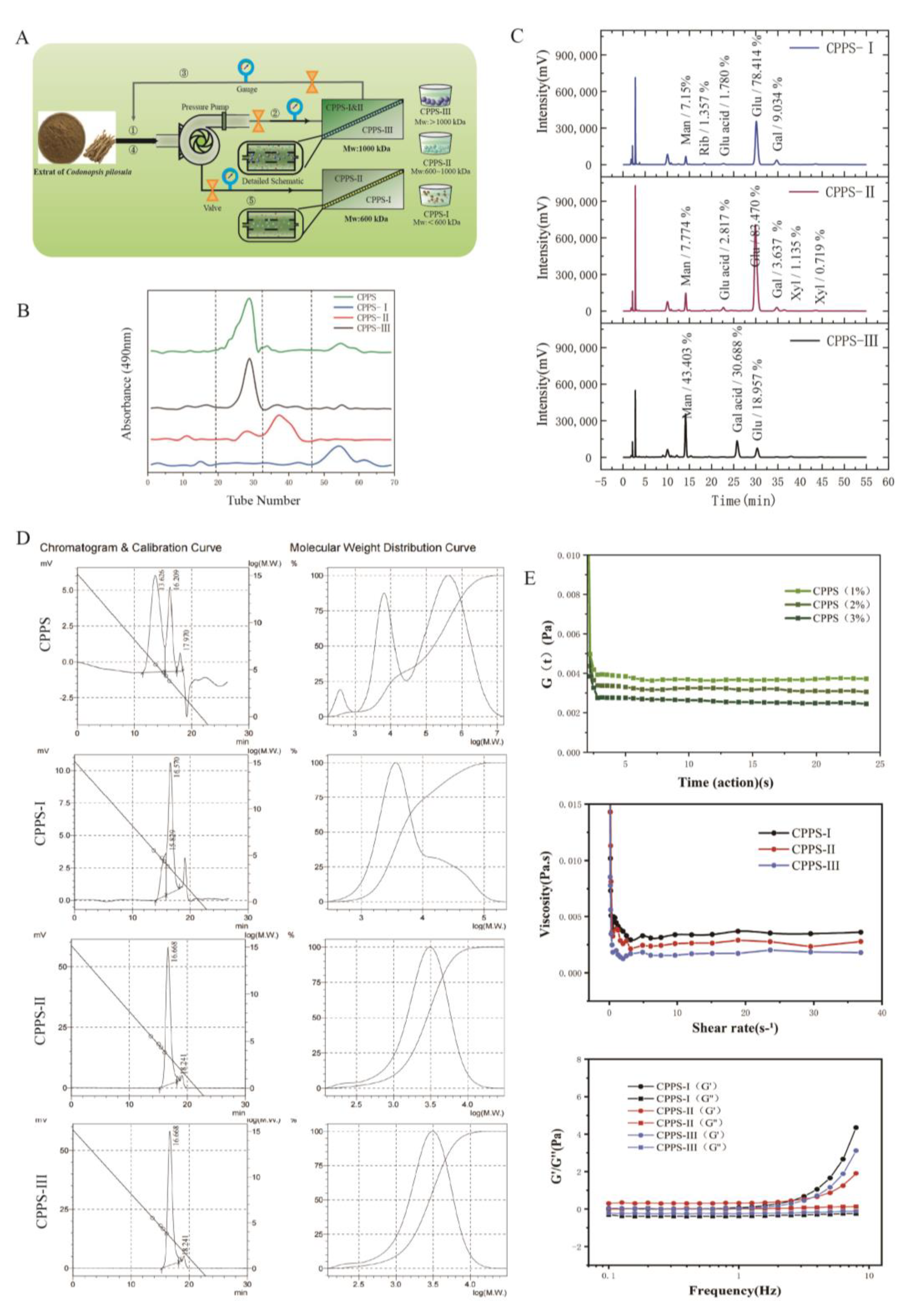
2.1. Purification and Structural Characterization of Polysaccharides
2.1.1. Chemical Composition Analysis
2.1.2. The Rheological Analysis
2.1.3. UV, FT-IR, and NMR Analyses
2.1.4. Morphological Properties
2.2. In Vitro Experiments
Antitumor and Immunomodulatory Activity of Three Polysaccharides
2.3. In Vivo Experiments
2.3.1. Antitumor Effect In Vivo
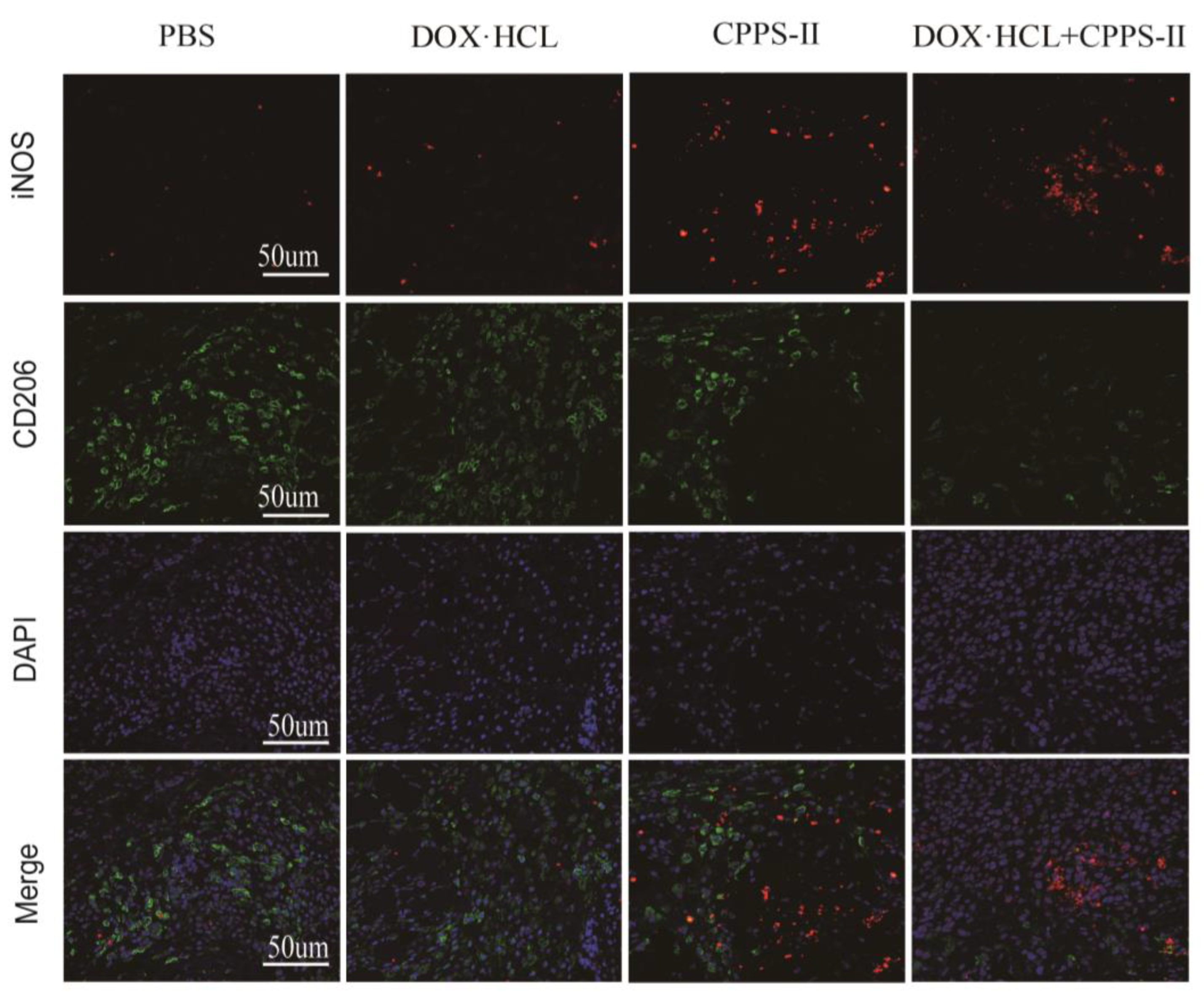
2.3.2. TAMs Population and Cytokine Expression Profiles in the TME
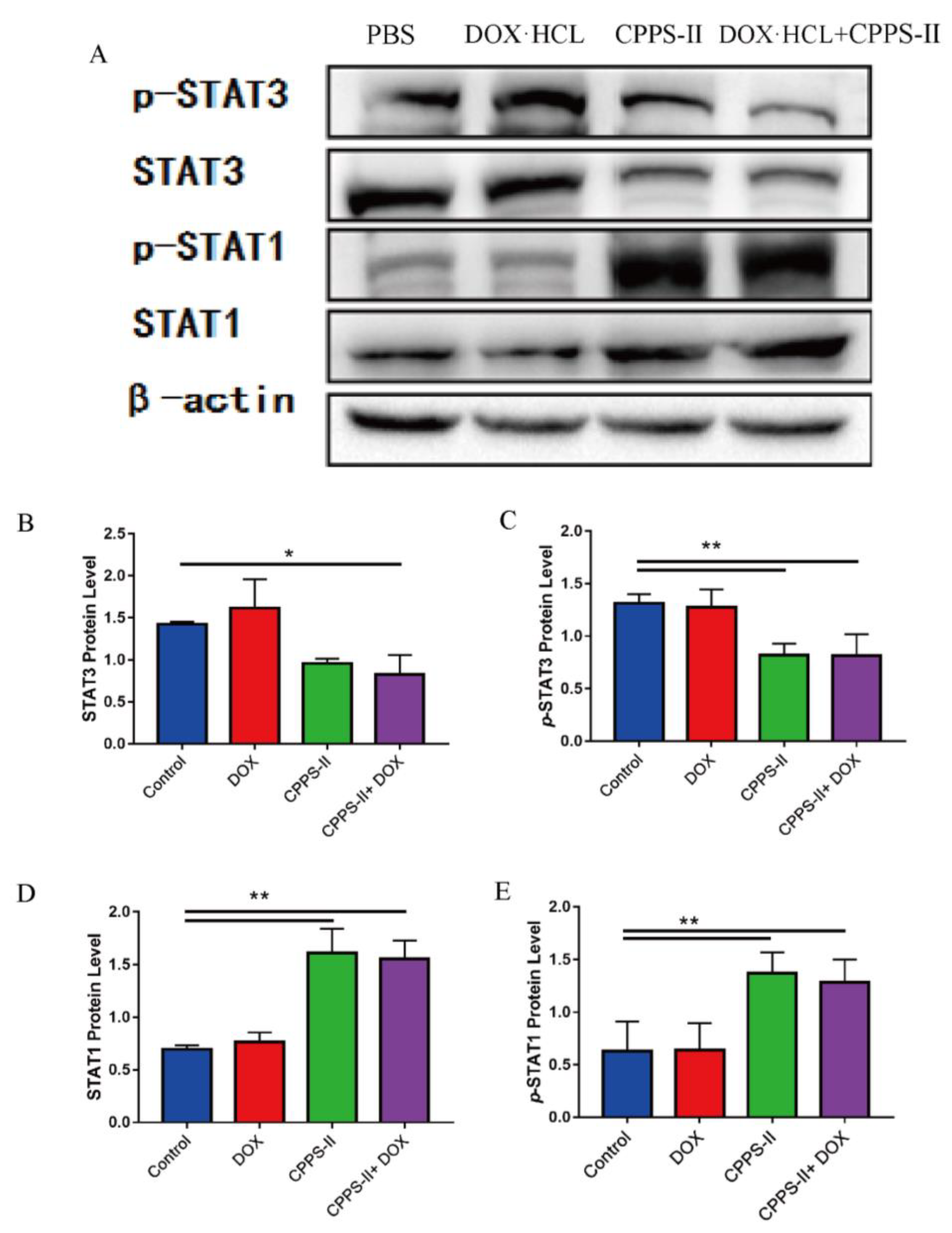
2.3.3. Signaling Pathway Determinations
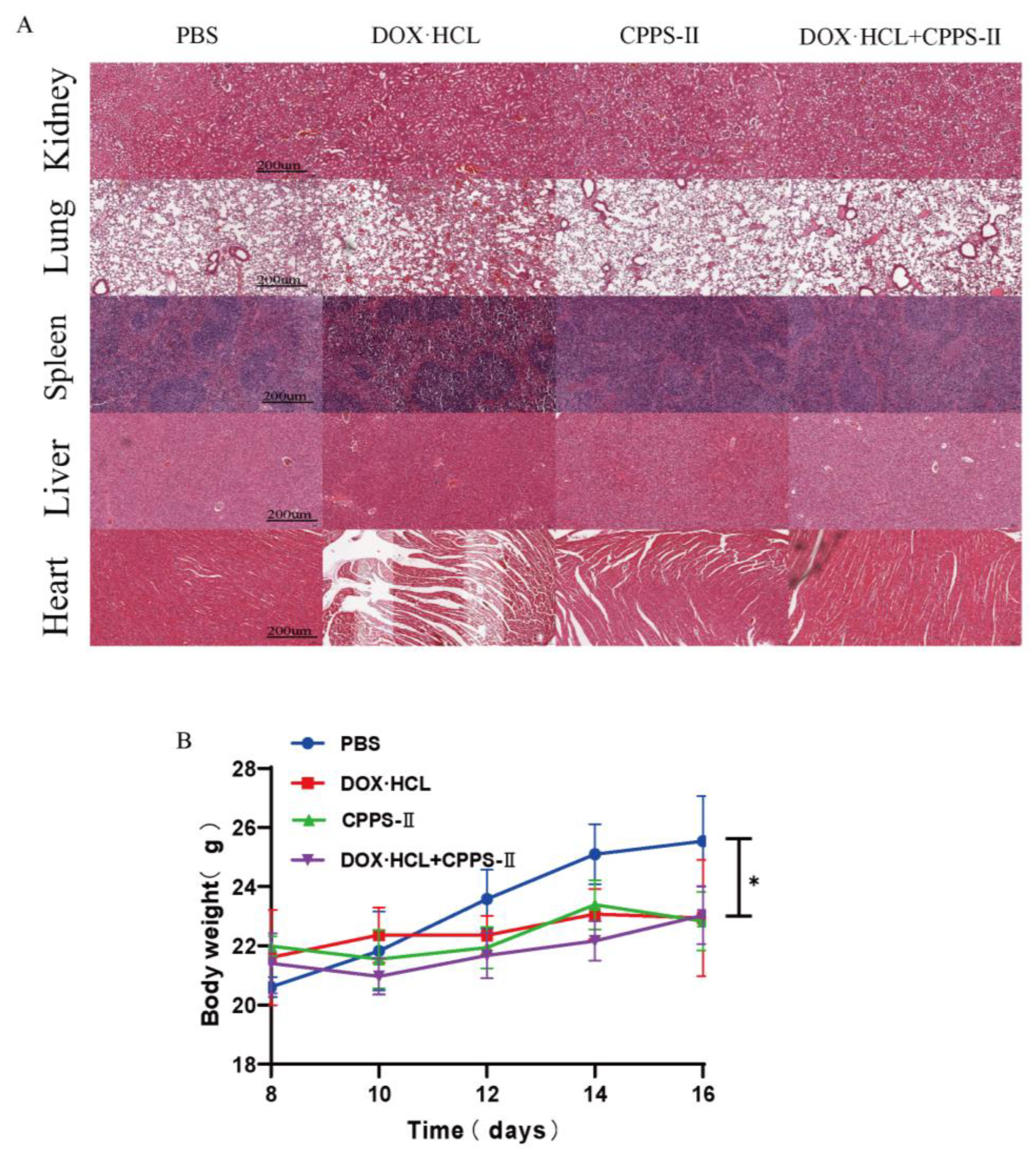
2.4. Safety Evaluation
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Polysaccharide Extraction and Purification
4.3. Polysaccharide Characterization
4.3.1. Molecular Weight and Homogeneity
4.3.2. Rheological Properties of CPPS
4.3.3. Qualitative Analysis of Monosaccharide Composition
4.3.4. UV and IR Spectrum Analysis
4.3.5. NMR Analysis
4.3.6. SEM Analysis
4.3.7. AFM Analysis
4.4. In Vitro Evaluation
4.4.1. Cell Culture
4.4.2. Cell Viability Assay
4.4.3. Measurement of NO
4.4.4. Macrophage Polarization
4.4.5. Cell Cycle
4.4.6. Annexin V-FITC/PI Detection
4.5. In Vivo Anti-Tumor Activity Evalution
4.5.1. Tumor Xenograft Models
4.5.2. Antitumor Efficiency
4.5.3. Immunohistochemical Staining Analysis
4.5.4. TUNEL Assay
4.5.5. Analysis of Macrophage Polarization by Flow Cytometry
4.5.6. Immunofluorescence Examination of Macrophages
4.5.7. ELISA Experiments
4.5.8. Western Blot Assay
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alatrash, G.; Jakher, H.; Stafford, P.D.; Mittendorf, E.A. Cancer immunotherapies, their safety and toxicity. Expert Opin. Drug Saf. 2013, 12, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, L.B.; Salama, A.K.S. A review of cancer immunotherapy toxicity. CA Cancer J. Clin. 2020, 70, 86–104. [Google Scholar] [CrossRef] [Green Version]
- Bai, R.B.; Zhang, Y.J.; Fan, J.M.; Jia, X.S.; Li, D.; Wang, Y.P.; Zhou, J.; Yan, Q.; Hu, F.D. Immune-enhancement effects of oligosaccharides from Codonopsis pilosula on cyclophosphamide induced immunosuppression in mice. Food Funct. 2020, 11, 3306–3315. [Google Scholar] [CrossRef] [PubMed]
- Luan, F.; Ji, Y.; Peng, L.; Liu, Q.; Cao, H.; Yang, Y.; He, X.; Zeng, N. Extraction, purification, structural characteristics and biological properties of the polysaccharides from Codonopsis pilosula: A review. Carbohydr. Polym. 2021, 261, 117863. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.M.; Liu, J.S.; Wang, M.; Cao, T.T.; Qi, Y.D.; Zhang, B.G.; Sun, X.B.; Liu, H.T.; Xiao, P.G. Traditional uses, phytochemistry, pharmacology and toxicology of Codonopsis: A review. J. Ethnopharmacol. 2018, 219, 50–70. [Google Scholar] [CrossRef]
- He, J.Y.; Ma, N.; Zhu, S.; Komatsu, K.; Li, Z.Y.; Fu, W.M. The genus Codonopsis (Campanulaceae): A review of phytochemistry, bioactivity and quality control. J. Nat. Med. 2015, 69, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Fan, Y.; Wang, D.; Hu, Y.; Liu, J.; Zhao, X.; Guo, L.; Zhao, X.; Yuan, J.; Zhang, F. Optimization on preparation condition of epimedium polysaccharide liposome and evaluation of its adjuvant activity. Int. J. Biol. Macromol. 2012, 50, 207–213. [Google Scholar] [CrossRef]
- Sun, Y.X. Immunological adjuvant effect of a water-soluble polysaccharide, CPP, from the roots of Codonopsis pilosula on the immune responses to ovalbumin in mice. Chem. Biodivers. 2009, 6, 890–896. [Google Scholar] [CrossRef]
- Zou, Y.F.; Zhang, Y.Y.; Paulsen, B.S.; Rise, F.; Chen, Z.L.; Jia, R.Y.; Li, L.X.; Song, X.; Feng, B.; Tang, H.Q.; et al. Structural features of pectic polysaccharides from stems of two species of Radix Codonopsis and their antioxidant activities. Int. J. Biol. Macromol. 2020, 159, 704–713. [Google Scholar] [CrossRef]
- Feng, Y.J.; Wang, X.X.; Zhuang, P.Y.; Zhang, D.Y.; Gao, L.; Chen, J.M.; Han, G. Study on chemical constituents of Codonopsis pilosula. China J. Chin. Mater. Med. 2017, 42, 135–139. [Google Scholar]
- Qin, F.Y.; Cheng, L.Z.; Yan, Y.M.; Liu, B.H.; Cheng, Y.X. Two novel proline-containing catechin glucoside from water-soluble extract of Codonopsis pilosula. Molecules 2018, 23, 180. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Liu, Y.; Guo, Q.; Xu, C.; Zhu, C.; Shi, J. Sesquiterpene glycosides from the roots of Codonopsis pilosula. Acta Pharm. Sin. B 2016, 6, 46–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, T.; Cheng, L.Z.; Yan, Y.M.; Liu, B.H.; Qin, F.Y.; Xu, F.R.; Cheng, Y.X. Two new triterpenoids from the roots of Codonopsis pilosula. Molecules 2018, 23, 383. [Google Scholar] [CrossRef] [Green Version]
- Wakana, D.; Kawahara, N.; Goda, Y. Two new pyrrolidine alkaloids, codonopsinol C and codonopiloside A, isolated from Codonopsis pilosula. Chem. Pharm. Bull. 2013, 61, 1315–1317. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Sun, L.; Shen, D.; Ren, A.; Ma, F.; Tai, G.; Fan, L.; Zhou, Y. Beta-1,6 glucan converts tumor-associated macrophages into an M1-like phenotype. Carbohydr. Polym. 2020, 247, 116715. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhao, J.; Wei, Y.; Yu, G.; Li, F.; Li, Q. Structural characterization and mechanisms of macrophage immunomodulatory activity of a pectic polysaccharide from Cucurbita moschata Duch. Carbohydr. Polym. 2021, 269, 118288. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Zhang, Y.; Jia, X.; Fan, J.; Hou, X.; Wang, Y.; Li, X.; Han, J.; Hu, F. Isolation, characterization and immunomodulatory activity of oligosaccharides from Codonopsis pilosula. J. Funct. Foods 2020, 72, 104070. [Google Scholar] [CrossRef]
- Liu, H.; Amakye, W.K.; Ren, J. Codonopsis pilosula polysaccharide in synergy with dacarbazine inhibits mouse melanoma by repolarizing M2-like tumor-associated macrophages into M1-like tumor-associated macrophages. Biomed. Pharmacother. 2021, 142, 112016. [Google Scholar] [CrossRef]
- Saha, S.; Galhardi, L.C.; Yamamoto, K.A.; Linhares, R.E.; Bandyopadhyay, S.S.; Sinha, S.; Nozawa, C.; Ray, B. Water-extracted polysaccharides from Azadirachta indica leaves: Structural features, chemical modification and anti-bovine herpesvirus type 1 (BoHV-1) activity. Int. J. Biol. Macromol. 2010, 47, 640–645. [Google Scholar] [CrossRef]
- Cai, M.; Xing, H.; Tian, B.; Xu, J.; Li, Z.; Zhu, H.; Yang, K.; Sun, P. Characteristics and antifatigue activity of graded polysaccharides from Ganoderma lucidum separated by cascade membrane technology. Carbohydr. Polym. 2021, 269, 118329. [Google Scholar] [CrossRef]
- Xu, Z.; Li, X.; Feng, S.; Liu, J.; Zhou, L.; Yuan, M.; Ding, C. Characteristics and bioactivities of different molecular weight polysaccharides from camellia seed cake. Int. J. Biol. Macromol. 2016, 91, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Ma, Q.; Xue, Z.; Gao, X.; Chen, H. Studies on the binding characteristics of three polysaccharides with different molecular weight and flavonoids from corn silk (Maydis stigma). Carbohydr. Polym. 2018, 198, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Lv, X.; Huang, W.; Yao, W.; Gao, X. Characterization and hypoglycemic effect of a neutral polysaccharide extracted from the residue of Codonopsis pilosula. Carbohydr. Polym. 2018, 197, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.F.; Zhang, Y.Y.; Fu, Y.P.; Inngjerdingen, K.T.; Paulsen, B.S.; Feng, B.; Zhu, Z.K.; Li, L.X.; Jia, R.Y.; Huang, C.; et al. A polysaccharide Isolated from Codonopsis pilosula with immunomodulation effects both in vitro and in vivo. Molecules 2019, 24, 3632. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, C.; Li, M.; Liu, C.; Wang, J.; Ou, X.; Han, Y. Antimicrobial peptides mediate apoptosis by changing mitochondrial membrane permeability. Int. J. Mol. Sci. 2022, 23, 12732. [Google Scholar] [CrossRef]
- Gunassekaran, G.R.; Poongkavithai Vadevoo, S.M.; Baek, M.C.; Lee, B. M1 macrophage exosomes engineered to foster M1 polarization and target the IL-4 receptor inhibit tumor growth by reprogramming tumor-associated macrophages into M1-like macrophages. Biomaterials 2021, 278, 121137. [Google Scholar] [CrossRef]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-associated macrophages in tumor immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef]
- Liu, K.; Ren, T.; Huang, Y.; Sun, K.; Bao, X.; Wang, S.; Zheng, B.; Guo, W. Apatinib promotes autophagy and apoptosis through VEGFR2/STAT3/BCL-2 signaling in osteosarcoma. Cell Death Dis. 2017, 8, 3015. [Google Scholar] [CrossRef] [Green Version]
- Huangfu, N.; Zheng, W.; Xu, Z.; Wang, S.; Wang, Y.; Cheng, J.; Li, Z.; Cheng, K.; Zhang, S.; Chen, X.; et al. RBM4 regulates M1 macrophages polarization through targeting STAT1-mediated glycolysis. Int. Immunopharmacol. 2020, 83, 106432. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.; Li, J.; Chen, Y.; Zhang, L. The structures and biological functions of polysaccharides from traditional Chinese herbs. Prog. Mol. Biol. Transl. Sci. 2019, 163, 423–444. [Google Scholar]
- Tardi, P.G.; Boman, N.L.; Cullis, P.R. Liposomal doxorubicin. J. Drug Target. 1996, 4, 129–140. [Google Scholar] [CrossRef]
- Xu, C.; Liu, Y.; Yuan, G.; Guan, M. The contribution of side chains to antitumor activity of a polysaccharide from Codonopsis pilosula. Int. J. Biol. Macromol. 2012, 50, 891–894. [Google Scholar] [CrossRef]
- Yang, S.; Sun, J.; Gu, D.; Li, P.; Yao, L.; Shi, D.; Guo, S.; Liu, C. Antioxidant activities of sulfated Codonopsis polysaccharides in acute oxidative stress. J. Food Biochem. 2021, 45, 13974. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.; Zhang, L.; Yuan, Y.; Yu, D. Codonopsis lanceolata polysaccharide CLPS alleviates high fat/high sucrose diet-induced insulin resistance via anti-oxidative stress. Int. J. Biol. Macromol. 2020, 145, 944–949. [Google Scholar] [CrossRef]
- Jing, Y.; Li, A.; Liu, Z.; Yang, P.; Wei, J.; Chen, X.; Zhao, T.; Bai, Y.; Zha, L.; Zhang, C. Absorption of Codonopsis pilosula saponins by coexisting polysaccharides alleviates gut microbial dysbiosis with dextran sulfate sodium-induced colitis in model mice. Biomed. Res. Int. 2018, 2018, 1781036. [Google Scholar] [CrossRef] [Green Version]
- Su, X.L.; Wang, J.W.; Che, H.; Wang, C.F.; Jiang, H.; Lei, X.; Zhao, W.; Kuang, H.X.; Wang, Q.H. Clinical application and mechanism of traditional Chinese medicine in treatment of lung cancer. Chin. Med. J. 2020, 133, 2987–2997. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Yang, K.; Zhang, G.; Li, S.; Gong, H.; Liu, M.; Dai, X. Structure characteristics of low molecular weight pectic polysaccharide and its anti-aging capability by modulating the intestinal homeostasis. Carbohydr. Polym. 2023, 303, 120467. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; Hecquet, P.E.; Kouach, M.; Thuru, X.; Goossens, J.F. Chemical reactivity and uses of 1-phenyl-3-methyl-5-pyrazolone (PMP), also known as edaravone. Bioorg. Med. Chem. 2020, 28, 115463. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.X.; Li, N.; Han, M.M.; Ye, F.; Liu, T.; Ye, H.Y.; Zheng, T.T.; Wu, J.J.; Li, Y.; Lv, S.; et al. Rhodiola rosea polysaccharides-based nanoparticles loaded with DOX boosts chemo-immunotherapy for triple-negative breast cancer by re-educating Tumor-associated macrophages. Int. J. Biol. Macromol. 2023, 239, 124110. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Song, K.; Wang, S.; Zhang, C.; Zhuang, M.; Wang, Y.; Liu, T. Anti-tumor potential of astragalus polysaccharides on breast cancer cell line mediated by macrophage activation. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 98, 685–695. [Google Scholar] [CrossRef]
- Wu, C.; Zheng, T.; Chen, H.; Zou, P.; Zhang, M.; Wang, J.; Li, N.; Zhang, Y.; Li, Y.; Dong, Z. Effect and Mechanism of Pharmaceutical Excipients on Berberine to Alleviate Ulcerative Colitis via Regulating Gut Microbiota. Molecules 2022, 27, 5997. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, N.; Xiong, Y.-X.; Ye, F.; Jin, B.; Wu, J.-J.; Han, M.-M.; Liu, T.; Fan, Y.-K.; Li, C.-Y.; Liu, J.-S.; et al. Isolation, Purification, and Structural Characterization of Polysaccharides from Codonopsis pilosula and Their Anti-Tumor Bioactivity by Immunomodulation. Pharmaceuticals 2023, 16, 895. https://doi.org/10.3390/ph16060895
Li N, Xiong Y-X, Ye F, Jin B, Wu J-J, Han M-M, Liu T, Fan Y-K, Li C-Y, Liu J-S, et al. Isolation, Purification, and Structural Characterization of Polysaccharides from Codonopsis pilosula and Their Anti-Tumor Bioactivity by Immunomodulation. Pharmaceuticals. 2023; 16(6):895. https://doi.org/10.3390/ph16060895
Chicago/Turabian StyleLi, Nan, Ying-Xia Xiong, Fan Ye, Bing Jin, Jin-Jia Wu, Miao-Miao Han, Tian Liu, Yi-Kai Fan, Cun-Yu Li, Jiu-Shi Liu, and et al. 2023. "Isolation, Purification, and Structural Characterization of Polysaccharides from Codonopsis pilosula and Their Anti-Tumor Bioactivity by Immunomodulation" Pharmaceuticals 16, no. 6: 895. https://doi.org/10.3390/ph16060895
APA StyleLi, N., Xiong, Y. -X., Ye, F., Jin, B., Wu, J. -J., Han, M. -M., Liu, T., Fan, Y. -K., Li, C. -Y., Liu, J. -S., Zhang, Y. -H., Sun, G. -B., Zhang, Y., & Dong, Z. -Q. (2023). Isolation, Purification, and Structural Characterization of Polysaccharides from Codonopsis pilosula and Their Anti-Tumor Bioactivity by Immunomodulation. Pharmaceuticals, 16(6), 895. https://doi.org/10.3390/ph16060895