
Affibody Molecules Intended for Receptor-Mediated Transcytosis via the Transferrin Receptor


Abstract
:1. Introduction
2. Results
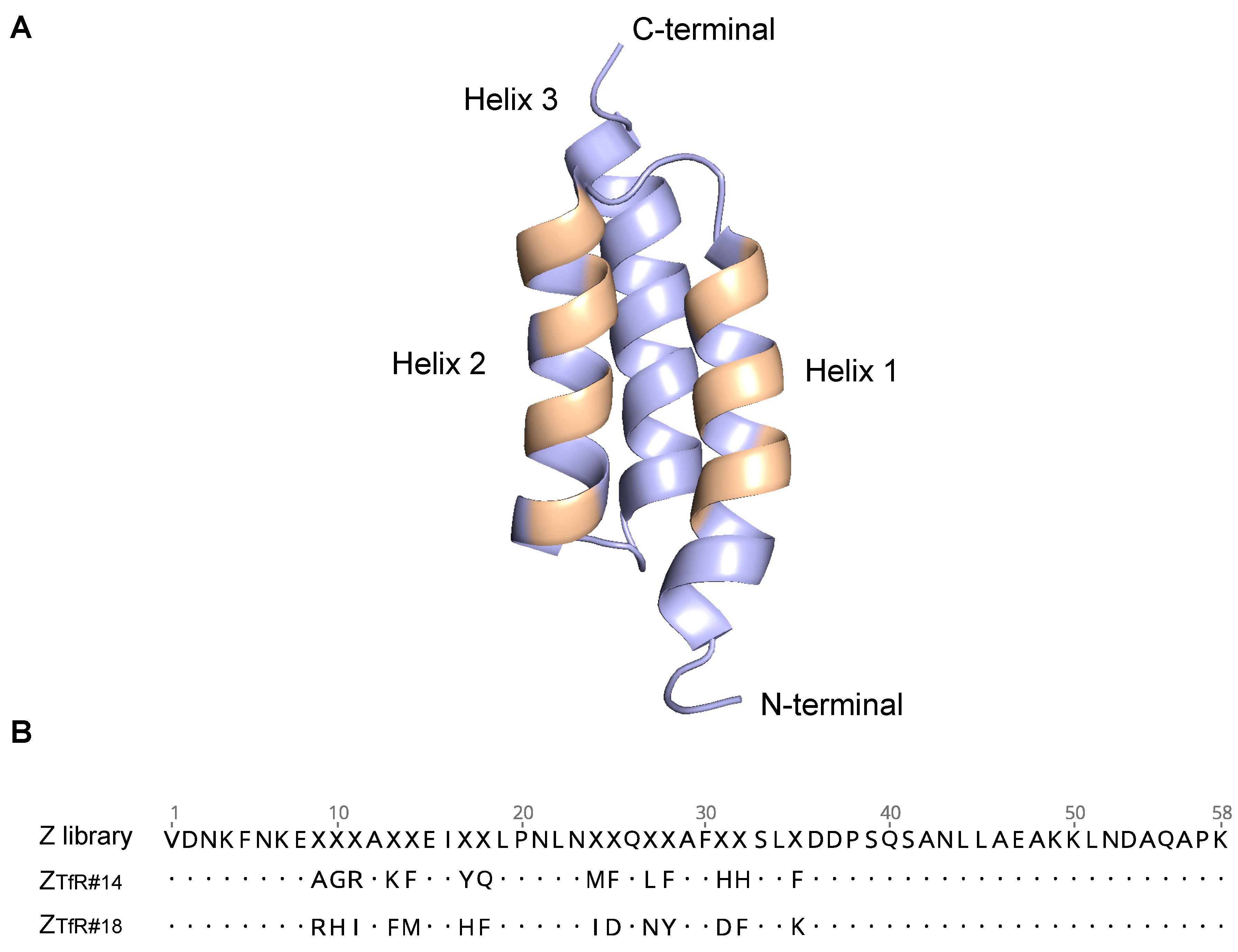
2.1. Phage Display Selections
2.2. Production of His6-Tagged and ABD-Fused Affibody Molecules
2.3. Flow Cytometry Analysis of ZTfR Candidates Aimed to Bind to Murine and Human Cells
2.4. pH-Dependent Binding to TfR Expressing Cells
2.5. Surface Plasmon Resonance Analysis of Binding between ZTfR and TfR
2.6. Single Amino Acid Mutagenesis of ZTfR#14 and ZTfR#18
2.7. Characterization of Second-Generation Affibody Molecules Binding TfR
2.8. Flow Cytometric Endocytosis Assay
2.9. Transcytosis across a Murine In Vitro BBB Model
3. Discussion
4. Materials and Methods
4.1. Protein Labeling
4.2. Phage Display Selections
4.3. Production of Recombinant Affibody Molecules in E. coli
4.4. Cultivation of TfR-Positive Cell Lines
4.5. Flow Cytometry Analysis of ZTfR-Binding to Murine and Human Cells
4.6. Circular Dichroism Analysis for Secondary Structure
4.7. Biosensor Analysis of the ZTfR and TfR Interaction of Both Murine and Human TfR
4.8. Single Amino Acid Mutagenesis and E. coli Surface Display Binding Analysis
4.9. Analysis of TfR-Binding Using E. coli Surface Display and Flow Cytometry
4.10. Production and Labeling of Second-Generation Affibody Molecules
4.11. Analysis of Endocytosis Using Flow Cytometry
4.12. Transcytosis In Vitro Murine BBB Model
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, A.C.; Carter, P.J. Therapeutic antibodies for autoimmunity and inflammation. Nat. Rev. Immunol. 2010, 10, 301–316. [Google Scholar] [CrossRef]
- Zahavi, D.; Weiner, L. Monoclonal Antibodies in Cancer Therapy. Antibodies 2020, 9, 34. [Google Scholar] [CrossRef]
- McConnell, H.L.; Kersch, C.N.; Woltjer, R.L.; Neuwelt, E.A. The translational significance of the neurovascular unit. J. Biol. Chem. 2017, 292, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Zlokovic, B.V. Neurovascular mechanisms of Alzheimer’s neurodegeneration. Trends Neurosci. 2005, 28, 202–208. [Google Scholar] [CrossRef]
- Barar, J.; Rafi, M.A.; Pourseif, M.M.; Omidi, Y. Blood-brain barrier transport machineries and targeted therapy of brain diseases. BioImpacts 2016, 6, 225–248. [Google Scholar] [CrossRef] [Green Version]
- Saunders, N.R.; Habgood, M.D.; Møllgård, K.; Dziegielewska, K.M. The biological significance of brain barrier mechanisms: Help or hindrance in drug delivery to the central nervous system? F1000Research 2016, 5, 313. [Google Scholar] [CrossRef] [Green Version]
- Pardridge, W.M. Blood-Brain Barrier and Delivery of Protein and Gene Therapeutics to Brain. Front. Aging Neurosci. 2020, 11, 373. [Google Scholar] [CrossRef]
- Hawkins, R.A.; Mokashi, A.; Simpson, I.A. An active transport system in the blood-brain barrier may reduce levodopa availability. Exp. Neurol. 2005, 195, 267–271. [Google Scholar] [CrossRef]
- Walter, F.R.; Santa-Maria, A.R.; Mészáros, M.; Veszelka, S.; Dér, A.; Deli, M.A. Surface charge, glycocalyx, and blood-brain barrier function. Tissue Barriers 2021, 9, 1904773. [Google Scholar] [CrossRef]
- Lajoie, J.M.; Katt, M.E.; Waters, E.A.; Herrin, B.R.; Shusta, E.V. Identification of lamprey variable lymphocyte receptors that target the brain vasculature. Sci. Rep. 2022, 12, 6044. [Google Scholar] [CrossRef]
- Atwal, J.K.; Chen, Y.; Chiu, C.; Mortensen, D.L.; Meilandt, W.J.; Liu, Y.; Heise, C.E.; Hoyte, K.; Luk, W.; Lu, Y.; et al. A Therapeutic Antibody Targeting BACE1 Inhibits Amyloid-β Production In Vivo. Sci. Transl. Med. 2011, 3, 4ra43. [Google Scholar] [CrossRef]
- Yadav, D.B.; Maloney, J.A.; Wildsmith, K.R.; Fuji, R.N.; Meilandt, W.J.; Solanoy, H.; Lu, Y.; Peng, K.; Wilson, B.; Chan, P.; et al. Widespread brain distribution and activity following i.c.v. infusion of anti-β-secretase (BACE1) in nonhuman primates. Br. J. Pharmacol. 2017, 174, 4173–4185. [Google Scholar] [CrossRef] [Green Version]
- Alata, W.; Yogi, A.; Brunette, E.; Delaney, C.E.; Faassen, H.; Hussack, G.; Iqbal, U.; Kemmerich, K.; Haqqani, A.S.; Moreno, M.J.; et al. Targeting insulin-like growth factor-1 receptor (IGF1R) for brain delivery of biologics. FASEB J. 2022, 36, e22208. [Google Scholar] [CrossRef]
- Boado, R.J.; Zhang, Y.; Zhang, Y.; Xia, C.F.; Wang, Y.; Pardridge, W.M. Genetic engineering of a lysosomal enzyme fusion protein for targeted delivery across the human blood-brain barrier. Biotechnol. Bioeng. 2008, 99, 475–484. [Google Scholar] [CrossRef]
- Stocki, P.; Szary, J.; Rasmussen, C.L.M.; Demydchuk, M.; Northall, L.; Logan, D.B.; Gauhar, A.; Thei, L.; Moos, T.; Walsh, F.S.; et al. Blood-brain barrier transport using a high affinity, brain-selective VNAR antibody targeting transferrin receptor 1. FASEB J. 2021, 35, e21172. [Google Scholar] [CrossRef]
- Kariolis, M.S.; Wells, R.C.; Getz, J.A.; Kwan, W.; Mahon, C.S.; Tong, R.; Kim, J.; Srivastava, A.; Bedard, C.; Henne, K.R.; et al. Brain delivery of therapeutic proteins using an Fc fragment blood-brain barrier transport vehicle in mice and monkeys. Sci. Transl. Med. 2020, 12, eaay1359. [Google Scholar] [CrossRef]
- Wouters, Y.; Jaspers, T.; Rué, L.; Serneels, L.; De Strooper, B.; Dewilde, M. VHHs as tools for therapeutic protein delivery to the central nervous system. Fluids Barriers CNS 2022, 19, 79. [Google Scholar] [CrossRef]
- Mayle, K.M.; Le, A.M.; Kamei, D.T. The intracellular trafficking pathway of transferrin. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 264–281. [Google Scholar] [CrossRef] [Green Version]
- Lamb, J.E.; Ray, F.; Ward, J.H.; Kushner, J.P.; Kaplan, J. Internalization and subcellular localization of transferrin and transferrin receptors in HeLa cells. J. Biol. Chem. 1983, 258, 8751–8758. [Google Scholar] [CrossRef]
- Kissel, K.; Hamm, S.; Schulz, M.; Vecchi, A.; Garlanda, C.; Engelhardt, B. Immunohistochemical localization of the murine transferrin receptor (TfR) on blood–tissue barriers using a novel anti-TfR monoclonal antibody. Histochem. Cell Biol. 1998, 110, 63–72. [Google Scholar] [CrossRef]
- Hultqvist, G.; Syvänen, S.; Fang, X.T.; Lannfelt, L.; Sehlin, D. Bivalent brain shuttle increases antibody uptake by monovalent binding to the transferrin receptor. Theranostics 2017, 7, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.J.; Zhang, Y.; Kenrick, M.; Hoyte, K.; Luk, W.; Lu, Y.; Atwal, J.; Elliott, J.M.; Prabhu, S.; Watts, R.J.; et al. Boosting Brain Uptake of a Therapeutic Antibody by Reducing Its Affinity for a Transcytosis Target. Sci. Transl. Med. 2011, 3, 84ra44. [Google Scholar] [CrossRef] [PubMed]
- Niewoehner, J.; Bohrmann, B.; Collin, L.; Urich, E.; Sade, H.; Maier, P.; Rueger, P.; Stracke, J.O.; Lau, W.; Tissot, A.C.; et al. Increased Brain Penetration and Potency of a Therapeutic Antibody Using a Monovalent Molecular Shuttle. Neuron 2014, 81, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, F.; Bohrmann, B.; Niewoehner, J.; Fischer, J.A.A.; Rueger, P.; Tiefenthaler, G.; Moelleken, J.; Bujotzek, A.; Brady, K.; Singer, T.; et al. Brain Shuttle Antibody for Alzheimer’s Disease with Attenuated Peripheral Effector Function due to an Inverted Binding Mode. Cell Rep. 2018, 22, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessard, E.; Rennie, K.; Haqqani, A.; Ling, B.; Whitfield, J.; Paradis, A.; Araujo, J.; Yoganathan, N.; Gillard, J.; Stanimirovic, D.; et al. Pharmacokinetics and Pharmacodynamic Effect of a Blood-Brain Barrier-Crossing Fusion Protein Therapeutic for Alzheimer’s Disease in Rat and Dog. Pharm. Res. 2022, 39, 1497–1507. [Google Scholar] [CrossRef]
- Stocki, P.; Szary, J.M.; Jacobsen, C.L.M.; Demydchuk, M.; Northall, L.; Moos, T.; Walsh, F.S.; Lynn Rutkowski, J. High efficiency blood-brain barrier transport using a VNAR targeting the Transferrin Receptor 1 (TfR1). bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Crook, Z.R.; Girard, E.; Sevilla, G.P.; Merrill, M.; Friend, D.; Rupert, P.B.; Pakiam, F.; Nguyen, E.; Yin, C.; Ruff, R.O.; et al. A TfR-Binding Cystine-Dense Peptide Promotes Blood–Brain Barrier Penetration of Bioactive Molecules. J. Mol. Biol. 2020, 432, 3989–4009. [Google Scholar] [CrossRef]
- Bien-Ly, N.; Yu, Y.J.; Bumbaca, D.; Elstrott, J.; Boswell, C.A.; Zhang, Y.; Luk, W.; Lu, Y.; Dennis, M.S.; Weimer, R.M.; et al. Transferrin receptor (TfR) trafficking determines brain uptake of TfR antibody affinity variants. J. Exp. Med. 2014, 211, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Couch, J.A.; Yu, Y.J.; Zhang, Y.; Tarrant, J.M.; Fuji, R.N.; Meilandt, W.J.; Solanoy, H.; Tong, R.K.; Hoyte, K.; Luk, W.; et al. Addressing safety liabilities of TfR bispecific antibodies that cross the blood-brain barrier. Sci. Transl. Med. 2013, 5, 183ra57. [Google Scholar] [CrossRef]
- Villaseñor, R.; Schilling, M.; Sundaresan, J.; Lutz, Y.; Collin, L. Sorting Tubules Regulate Blood-Brain Barrier Transcytosis. Cell Rep. 2017, 21, 3256–3270. [Google Scholar] [CrossRef] [Green Version]
- Faresjö, R.; Bonvicini, G.; Fang, X.T.; Aguilar, X.; Sehlin, D.; Syvänen, S. Brain pharmacokinetics of two BBB penetrating bispecific antibodies of different size. Fluids Barriers CNS 2021, 18, 26. [Google Scholar] [CrossRef] [PubMed]
- Ståhl, S.; Gräslund, T.; Eriksson Karlström, A.; Frejd, F.Y.; Nygren, P.Å.; Löfblom, J. Affibody Molecules in Biotechnological and Medical Applications. Trends Biotechnol. 2017, 35, 691–712. [Google Scholar] [CrossRef] [PubMed]
- Engfeldt, T.; Renberg, B.; Brumer, H.; Nygren, P.Å.; Karlström, A.E. Chemical synthesis of triple-labelled three-helix bundle binding proteins for specific fluorescent detection of unlabelled protein. ChemBioChem 2005, 6, 1043–1050. [Google Scholar] [CrossRef]
- Andersson, K.G.; Persson, J.; Ståhl, S.; Löfblom, J. Autotransporter-Mediated Display of a Naïve Affibody Library on the Outer Membrane of Escherichia coli. Biotechnol. J. 2019, 14, e1800359. [Google Scholar] [CrossRef]
- Grönwall, C.; Jonsson, A.; Lindström, S.; Gunneriusson, E.; Ståhl, S.; Herne, N. Selection and characterization of Affibody ligands binding to Alzheimer amyloid β peptides. J. Biotechnol. 2007, 128, 162–183. [Google Scholar] [CrossRef]
- Klint, S.; Feldwisch, J.; Gudmundsdotter, L.; Dillner Bergstedt, K.; Gunneriusson, E.; Höidén Guthenberg, I.; Wennborg, A.; Nyborg, A.C.; Kamboj, A.P.; Peloso, P.M.; et al. Izokibep: Preclinical development and first-in-human study of a novel IL-17A neutralizing Affibody molecule in patients with plaque psoriasis. mAbs 2023, 15, 2209920. [Google Scholar] [CrossRef]
- Affibody, A.B. Izokibep, a Bispecific Molecule Targeting Interleukin-17A (IL-17). 2022. Available online: https://www.affibody.se/ (accessed on 25 January 2023).
- Hjelm, L.C.; Lindberg, H.; Ståhl, S.; Löfblom, J. Construction and Validation of a New Naïve Sequestrin Library for Directed Evolution of Binders against Aggregation-Prone Peptides. Int. J. Mol. Sci. 2023, 24, 836. [Google Scholar] [CrossRef] [PubMed]
- Boutajangout, A.; Lindberg, H.; Awwad, A.; Paul, A.; Baitalmal, R.; Almokyad, I.; Höidén-Guthenberg, I.; Gunneriusson, E.; Frejd, F.Y.; Härd, T.; et al. Affibody-Mediated Sequestration of Amyloid β Demonstrates Preventive Efficacy in a Transgenic Alzheimer’s Disease Mouse Model. Front. Aging Neurosci. 2019, 11, 64. [Google Scholar] [CrossRef]
- Meister, S.W.; Hjelm, L.C.; Dannemeyer, M.; Tegel, H.; Lindberg, H.; Ståhl, S.; Löfblom, J. An affibody molecule is actively transported into the cerebrospinal fluid via binding to the transferrin receptor. Int. J. Mol. Sci. 2020, 21, 2999. [Google Scholar] [CrossRef] [Green Version]
- Jonsson, A.; Dogan, J.; Herne, N.; Abrahmsén, L.; Nygren, P.Å. Engineering of a femtomolar affinity binding protein to human serum albumin. Protein Eng. Des. Sel. 2008, 21, 515–527. [Google Scholar] [CrossRef]
- Tolmachev, V.; Orlova, A.; Pehrson, R.; Galli, J.; Baastrup, B.; Andersson, K.; Sandström, M.; Rosik, D.; Carlsson, J.; Lundqvist, H.; et al. Radionuclide therapy of HER2-positive microxenografts using a 177Lu-labeled HER2-specific affibody molecule. Cancer Res. 2007, 67, 2773–2782. [Google Scholar] [CrossRef] [Green Version]
- Eklund, M.; Axelsson, L.; Uhlén, M.; Nygren, P.Å. Anti-idiotypic protein domains selected from protein A-based affibody libraries. Proteins Struct. Funct. Genet. 2002, 48, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Neiveyans, M.; Melhem, R.; Arnoult, C.; Bourquard, T.; Jarlier, M.; Busson, M.; Laroche, A.; Cerutti, M.; Pugnière, M.; Ternant, D.; et al. A recycling anti transferrin receptor 1 monoclonal antibody as an efficient therapy for erythroleukemia through target up regulation and antibody. mAbs 2019, 11, 359–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, B.; Moks, T.; Jansson, B.; Abrahmsén, L.; Elmblad, A.; Holmgren, E.; Henrichson, C.; Jones, T.A.; Uhlén, M. A synthetic IgG-binding domain based on staphylococcal protein A. Protein Eng. Des. Sel. 1987, 1, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlova, A.; Magnusson, M.; Eriksson, T.L.J.; Nilsson, M.; Larsson, B.; Höiden-Guthenberg, I.; Widström, C.; Carlsson, J.; Tolmachev, V.; Ståhl, S.; et al. Tumor imaging using a picomolar affinity HER2 binding Affibody molecule. Cancer Res. 2006, 66, 4339–4348. [Google Scholar] [CrossRef] [Green Version]
- Hjelm, L.C.; Hedhammar, M.; Löfblom, J. In vitro Blood–Brain barrier model based on recombinant spider silk protein nanomembranes for evaluation of transcytosis capability of biomolecules. Biochem. Biophys. Res. Commun. 2023, 669, 77–84. [Google Scholar] [CrossRef]
- Morrison, J.I.; Metzendorf, N.G.; Rofo, F.; Petrovic, A.; Hultqvist, G. A single-chain fragment constant design enables easy production of a monovalent blood-brain barrier transporter and provides an improved brain uptake at elevated doses. J. Neurochem. 2023, 165, 413–425. [Google Scholar] [CrossRef]
- Sade, H.; Baumgartner, C.; Hugenmatter, A.; Moessner, E.; Freskgård, P.O.; Niewoehner, J. A human blood-brain barrier transcytosis assay reveals antibody transcytosis influenced by pH-dependent receptor binding. PLoS ONE 2014, 9, e96340. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.J.; Atwal, J.K.; Zhang, Y.; Tong, R.K.; Wildsmith, K.R.; Tan, C.; Bien-Ly, N.; Hersom, M.; Maloney, J.A.; Meilandt, W.J.; et al. Therapeutic bispecific antibodies cross the blood-brain barrier in nonhuman primates. Sci. Transl. Med. 2014, 6, 261ra154. [Google Scholar] [CrossRef]
- Toth, A.E.; Nielsen, S.S.E.; Tomaka, W.; Abbott, N.J.; Nielsen, M.S. The endo-lysosomal system of bEnd.3 and hCMEC/D3 brain endothelial cells. Fluids Barriers CNS 2019, 16, 14. [Google Scholar] [CrossRef] [Green Version]
- Stocki, P.; Szary, J.; Demydchuk, M.; Northall, L.; Rasmussen, C.L.M.; Logan, D.B.; Gauhar, A.; Thei, L.; Coker, S.F.; Moos, T.; et al. CDR3 Variants of the TXB2 Shuttle with Increased TfR1 Association Rate and Enhanced Brain Penetration. Pharmaceutics 2023, 15, 739. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Gudmundsdotter, L.; Akal, A.; Gunneriusson, E.; Frejd, F.; Nygren, P.Å. An affibody-adalimumab hybrid blocks combined IL-6 and tnf-triggered serum amyloid a secretion in vivo. mAbs 2014, 6, 1598–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurer, J.; Jose, J.; Meyer, T.F.; Molekulare Biologie, A. Autodisplay: One-Component System for Efficient Surface Display and Release of Soluble Recombinant Proteins from Escherichia coli. J. Bacteriol. 1997, 179, 794–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | Tm [°C] | Refolding Capability |
|---|---|---|
| (HE)3-ZTfR#14_A9H-biotin | 64 | Yes |
| (HE)3-ZTfR#14_L27H-biotin | 66 | Partly |
| (HE)3-ZTfR#14_A9H_L27H-biotin | 62 | Partly |
| (HE)3-ZTfR#18_I11N-biotin | 52 | Partly |
| (HE)3-ZTfR#18_M14H--biotin | 62 | Yes |
| (HE)3-ZTfR#18_I11N_M14H-biotin | 63 | Yes |
| (HE)3-ZHER2-biotin | 62 | Yes |
| Sample | Calculated pH Environment by Standard Curve | Endocytosis Classification |
|---|---|---|
| (HE)3-ZHER2-FITC | 5.75 ± 0.02 | Late/lysosomal |
| (HE)3-ZTfR#14_A9H-FITC | 7.15 ± 0.31 | Partial cell surface or vesicle |
| (HE)3-ZTfR#14_L27H-FITC | 6.94 ± 0.02 | Early |
| (HE)3-ZTfR#14_A9H_L27H-FITC | 6.18 ± 0.04 | Recycling |
| (HE)3-ZTfR#18_I11N-FITC | 5.13 ± 0.02 | Lysosome |
| (HE)3-ZTfR#18_M14H-FITC | 6.56 ± 0.02 | Early |
| (HE)3-ZTfR#18_I11N_M14H-FITC | 5.16 ± 0.01 | Lysosome |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hjelm, L.C.; Lindberg, H.; Ståhl, S.; Löfblom, J. Affibody Molecules Intended for Receptor-Mediated Transcytosis via the Transferrin Receptor. Pharmaceuticals 2023, 16, 956. https://doi.org/10.3390/ph16070956
Hjelm LC, Lindberg H, Ståhl S, Löfblom J. Affibody Molecules Intended for Receptor-Mediated Transcytosis via the Transferrin Receptor. Pharmaceuticals. 2023; 16(7):956. https://doi.org/10.3390/ph16070956
Chicago/Turabian StyleHjelm, Linnea Charlotta, Hanna Lindberg, Stefan Ståhl, and John Löfblom. 2023. "Affibody Molecules Intended for Receptor-Mediated Transcytosis via the Transferrin Receptor" Pharmaceuticals 16, no. 7: 956. https://doi.org/10.3390/ph16070956
APA StyleHjelm, L. C., Lindberg, H., Ståhl, S., & Löfblom, J. (2023). Affibody Molecules Intended for Receptor-Mediated Transcytosis via the Transferrin Receptor. Pharmaceuticals, 16(7), 956. https://doi.org/10.3390/ph16070956