
Kinin B2 Receptor Mediates Cisplatin-Induced Painful Peripheral Neuropathy by Intracellular Kinase Pathways and TRPA1 Channel Sensitisation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
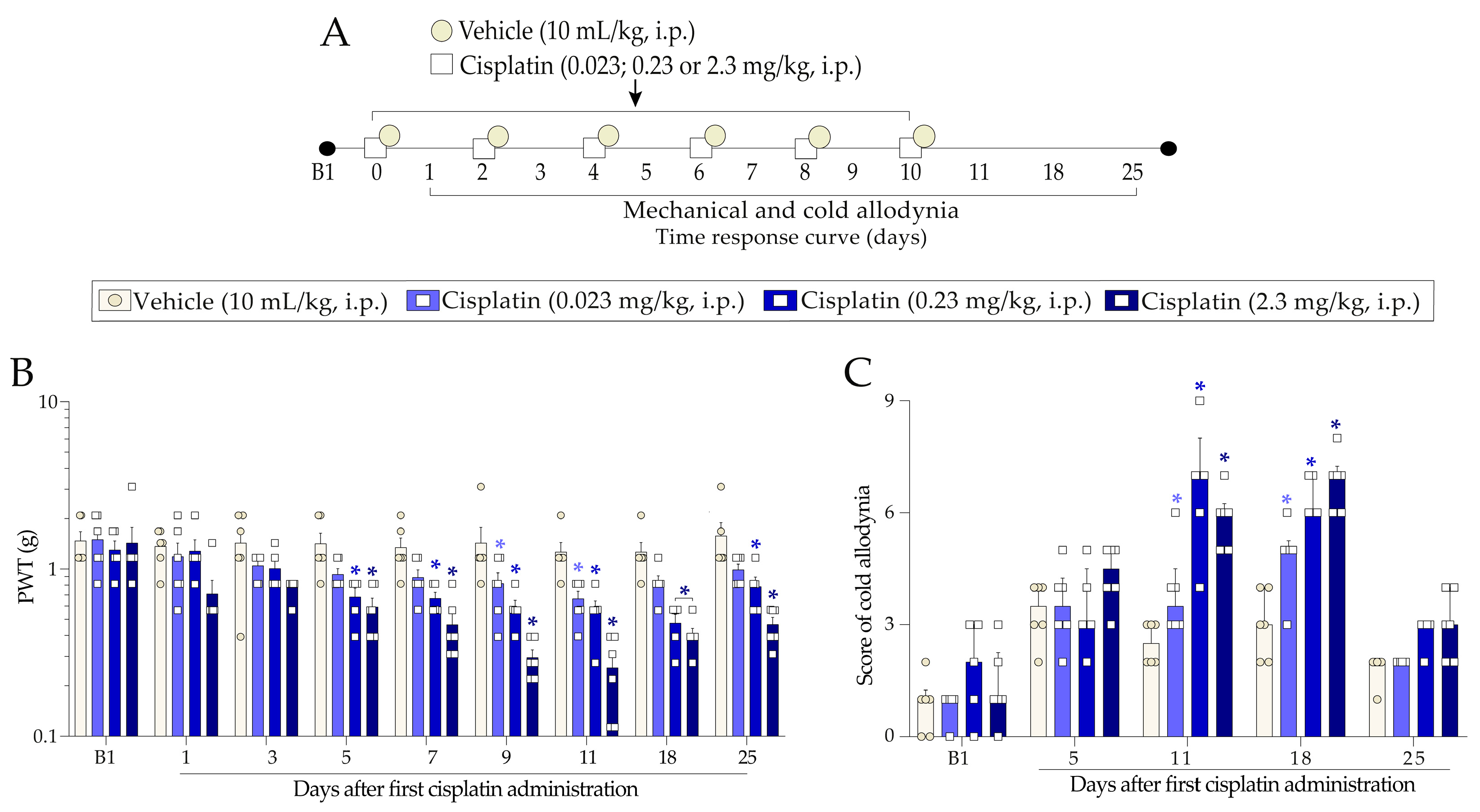
2.1. Cisplatin Induces Nociceptive Behaviours in Mice
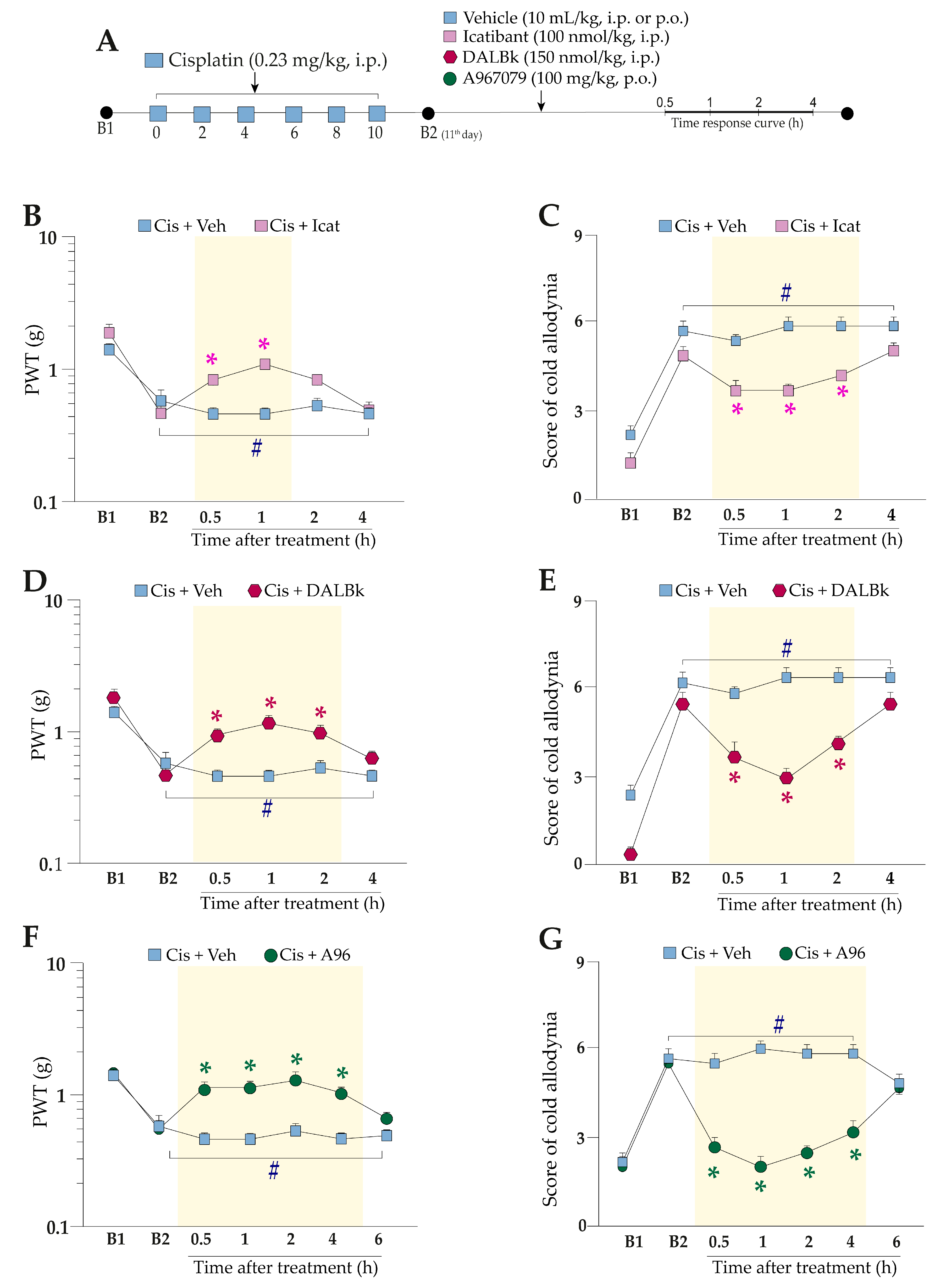
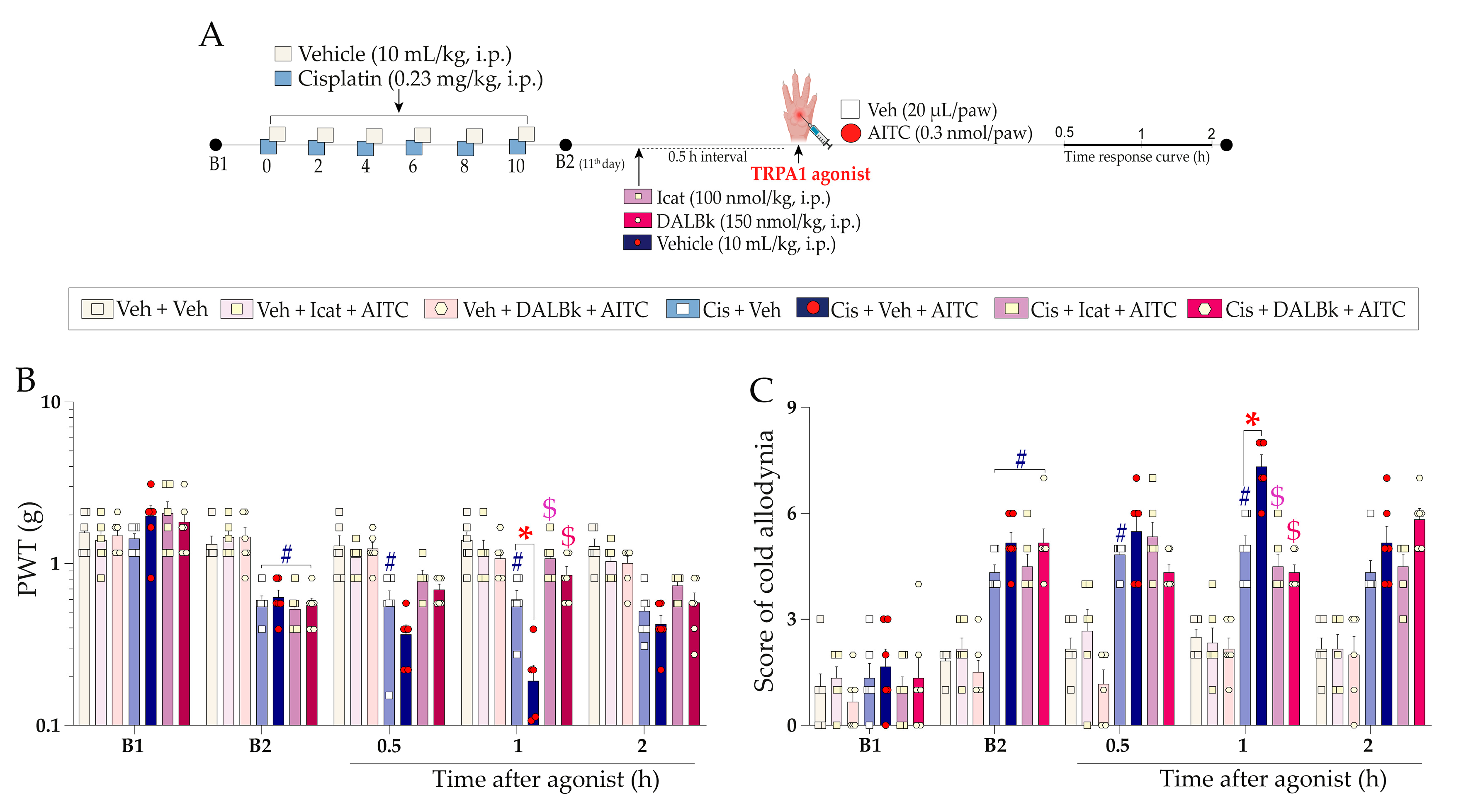
2.2. Kinin B2 and B1 Receptors and the TRPA1 Channel Antagonism Alleviate Pain Hypersensitivity Induced by Cisplatin
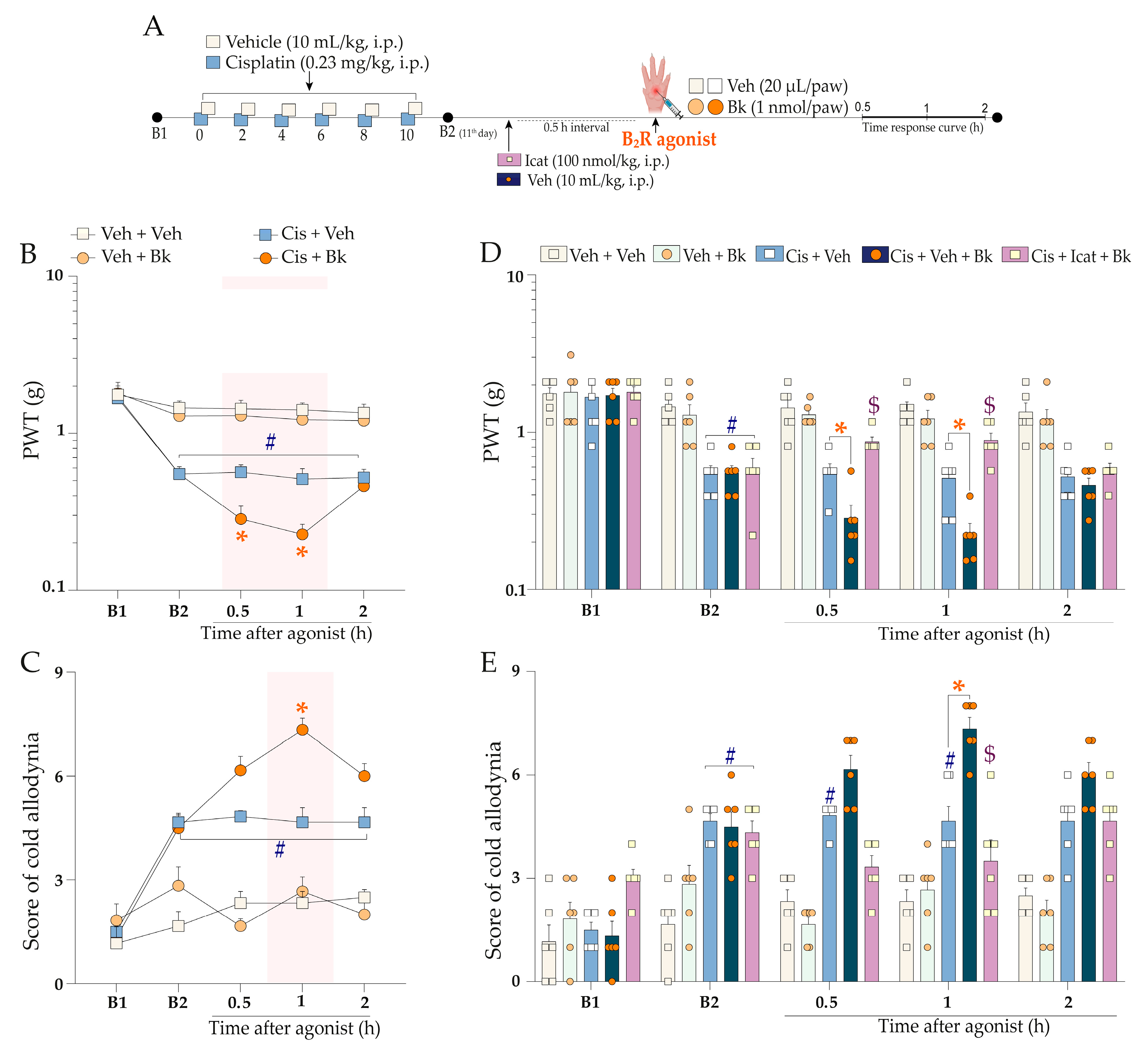
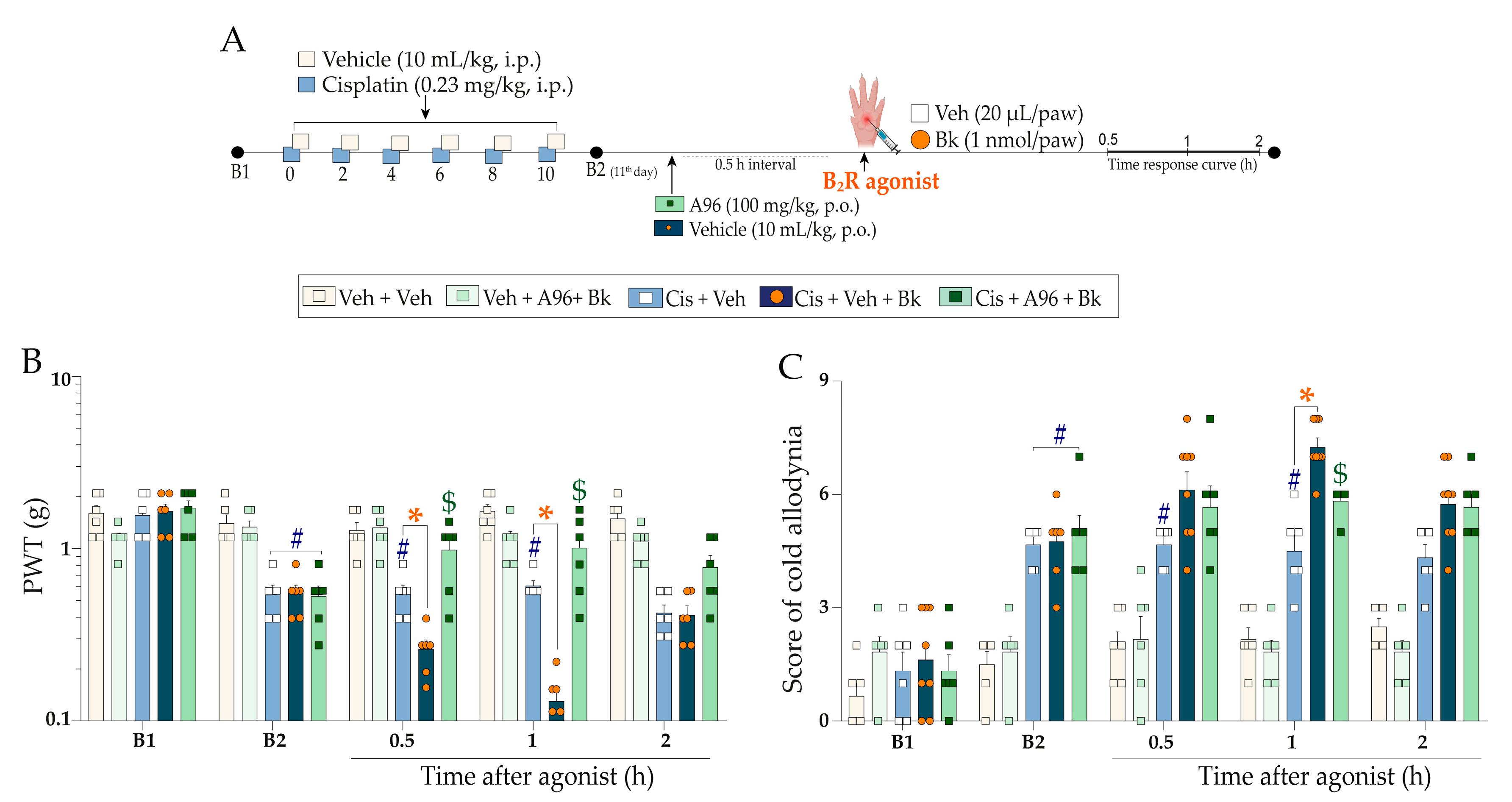
2.3. Functional Interaction of Kinin B2 Receptor and TRPA1 Channel Cooperate to Cisplatin-Induced Painful Peripheral Neuropathy
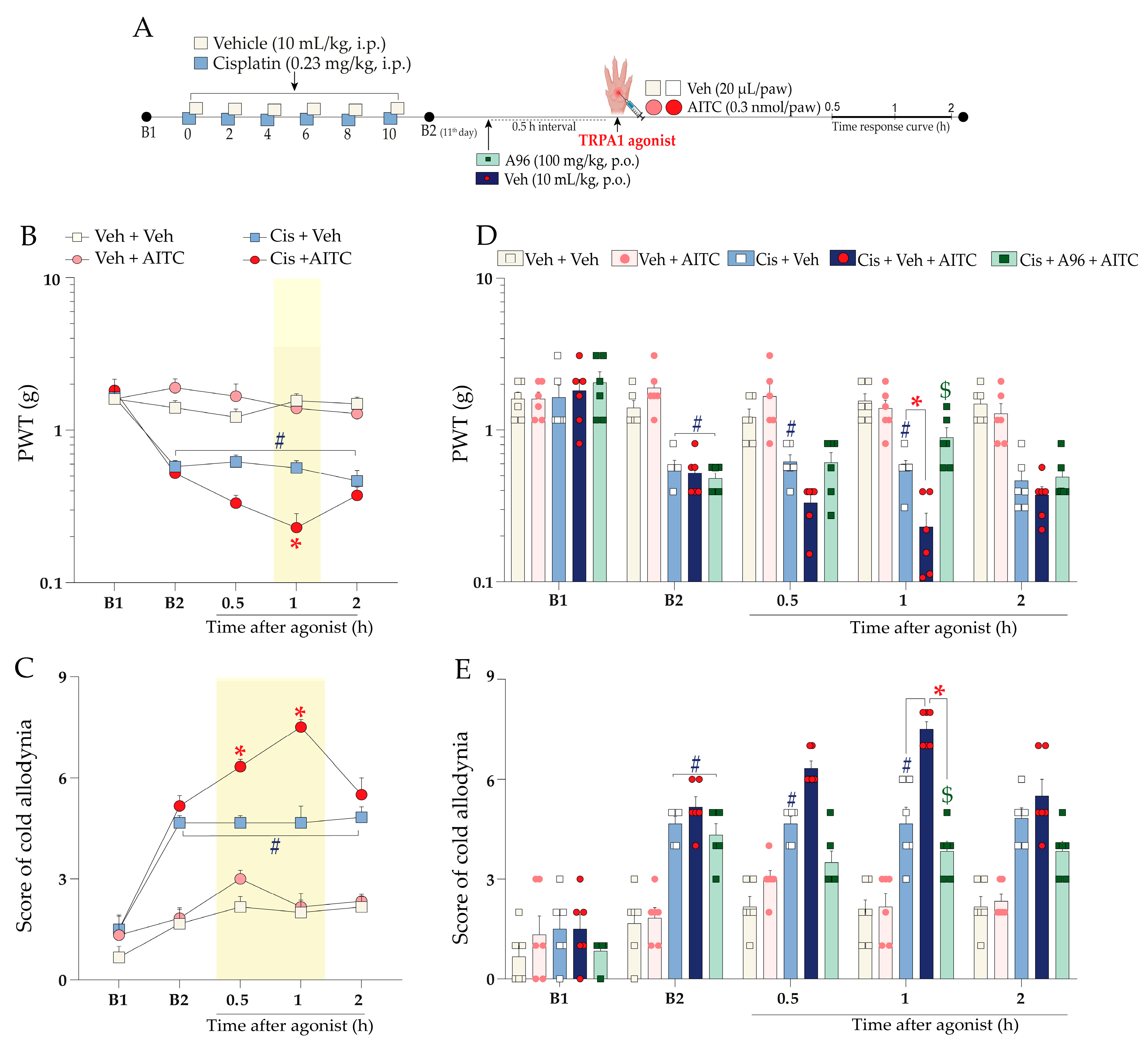
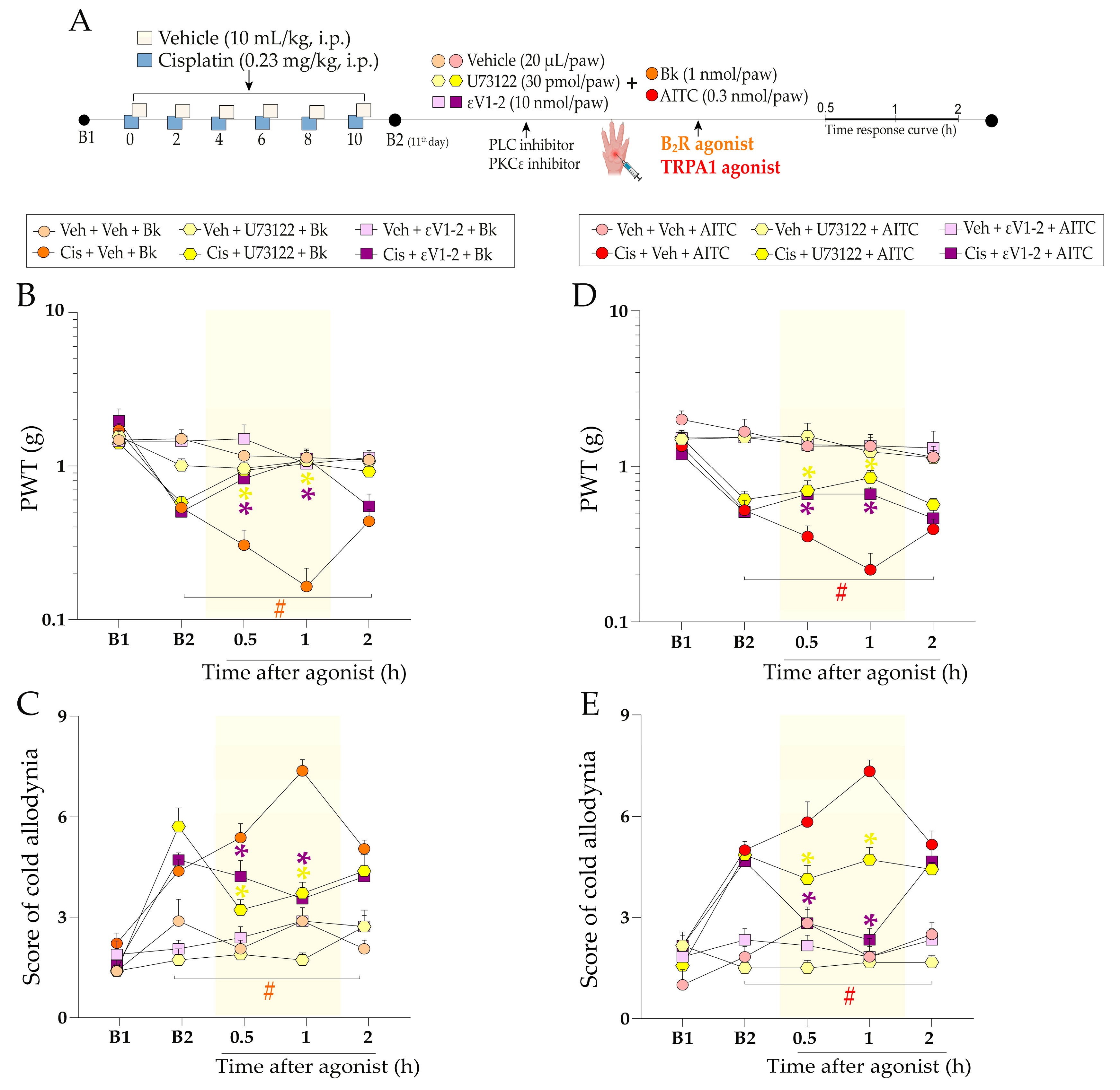
2.4. The Molecular Signalling Mechanisms Downstream of kinin B2 Receptor Activation Contribute to Sensitising the TRPA1 Channel in Mice Treated with Cisplatin
3. Discussion
4. Materials and Methods
4.1. Drugs and Reagents
4.2. Animals
4.3. Peripheral Neuropathy Model Induced by Cisplatin
4.4. Study Design
4.4.1. Characterisation of Cisplatin-Induced Painful Peripheral Neuropathy
4.4.2. Kinin B1 and B2 Receptors and TRPA1 Channel Involvement in Cisplatin-Induced Painful Peripheral Neuropathy
4.4.3. Functional Interaction between Kinin B2 Receptor and TRPA1 Channel in Cisplatin-Induced Painful Peripheral Neuropathy
4.4.4. Intracellular Pathways Dependent on Kinin B2 Receptor Activation and TRPA1 Channel Sensitisation
4.5. Nociceptive Parameters
4.5.1. Mechanical Allodynia Assessment
4.5.2. Cold Allodynia
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Nogueira, L.; Devasia, T.; Mariotto, A.B.; Yabroff, K.R.; Jemal, A.; Kramer, J.; Siegel, R.L. Cancer Treatment and Survivorship Statistics, 2022. CA Cancer J. Clin. 2022, 72, 409–436. [Google Scholar] [CrossRef] [PubMed]
- Emery, J.; Butow, P.; Lai-Kwon, J.; Nekhlyudov, L.; Rynderman, M.; Jefford, M. Management of Common Clinical Problems Experienced by Survivors of Cancer. Lancet 2022, 399, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, M.D.; Li, B.T.; Chaft, J.E.; Kris, M.G. Chemotherapy Remains an Essential Element of Personalized Care for Persons with Lung Cancers. Ann. Oncol. 2016, 27, 1829–1835. [Google Scholar] [CrossRef]
- Staff, N.P.; Grisold, A.; Grisold, W.; Windebank, A.J. Chemotherapy-Induced Peripheral Neuropathy: A Current Review. Ann. Neurol. 2017, 81, 772–781. [Google Scholar] [CrossRef]
- Argyriou, A.A.; Bruna, J.; Genazzani, A.A.; Cavaletti, G. Chemotherapy-Induced Peripheral Neurotoxicity: Management Informed by Pharmacogenetics. Nat. Rev. Neurol. 2017, 13, 492–504. [Google Scholar] [CrossRef]
- Calls, A.; Carozzi, V.; Navarro, X.; Monza, L.; Bruna, J. Pathogenesis of Platinum-Induced Peripheral Neurotoxicity: Insights from Preclinical Studies. Exp. Neurol. 2020, 325, 113141. [Google Scholar] [CrossRef]
- Albany, C.; Dockter, T.; Wolfe, E.; Le-Rademacher, J.; Wagner-Johnston, N.; Einhorn, L.; Lafky, J.M.; Smith, E.; Pachman, D.; Staff, N.; et al. Cisplatin-Associated Neuropathy Characteristics Compared with Those Associated with Other Neurotoxic Chemotherapy Agents (Alliance A151724). Support. Care Cancer 2021, 29, 833–840. [Google Scholar] [CrossRef]
- Travis, L.B.; Fossa, S.D.; Sesso, H.D.; Frisina, R.D.; Herrmann, D.N.; Beard, C.J.; Feldman, D.R.; Pagliaro, L.C.; Miller, R.C.; Vaughn, D.J.; et al. Chemotherapy-Induced Peripheral Neurotoxicity and Ototoxicity: New Paradigms for Translational Genomics. J. Natl. Cancer Inst. 2014, 106, dju044. [Google Scholar] [CrossRef] [Green Version]
- Avan, A.; Postma, T.J.; Ceresa, C.; Avan, A.; Cavaletti, G.; Giovannetti, E.; Peters, G.J. Platinum-Induced Neurotoxicity and Preventive Strategies: Past, Present, and Future. Oncologist 2015, 20, 411–432. [Google Scholar] [CrossRef] [Green Version]
- Flatters, S.J.L.; Dougherty, P.M.; Colvin, L.A. Clinical and Preclinical Perspectives on Chemotherapy-Induced Peripheral Neuropathy (CIPN): A Narrative Review. Br. J. Anaesth. 2017, 119, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Seretny, M.; Currie, G.L.; Sena, E.S.; Ramnarine, S.; Grant, R.; Macleod, M.R.; Colvin, L.A.; Fallon, M. Incidence, Prevalence, and Predictors of Chemotherapy-Induced Peripheral Neuropathy: A Systematic Review and Meta-Analysis. Pain 2014, 155, 2461–2470. [Google Scholar] [CrossRef] [Green Version]
- Colvin, L.A. Chemotherapy-Induced Peripheral Neuropathy: Where Are We Now? Pain 2019, 160, S1–S10. [Google Scholar] [CrossRef]
- Ibrahim, E.Y.; Ehrlich, B.E. Prevention of Chemotherapy-Induced Peripheral Neuropathy: A Review of Recent Findings. Crit. Rev. Oncol. Hematol. 2020, 145, 102831. [Google Scholar] [CrossRef]
- Starobova, H.; Vetter, I. Pathophysiology of Chemotherapy-Induced Peripheral Neuropathy. Front. Mol. Neurosci. 2017, 10, 174. [Google Scholar] [CrossRef] [Green Version]
- Alberti, P. Platinum-Drugs Induced Peripheral Neurotoxicity: Clinical Course and Preclinical Evidence. Expert Opin. Drug Metab. Toxicol. 2019, 15, 487–497. [Google Scholar] [CrossRef]
- Starobova, H.; Mueller, A.; Deuis, J.R.; Carter, D.A.; Vetter, I. Inflammatory and Neuropathic Gene Expression Signatures of Chemotherapy-Induced Neuropathy Induced by Vincristine, Cisplatin, and Oxaliplatin in C57BL/6J Mice. J. Pain 2020, 21, 182–194. [Google Scholar] [CrossRef]
- Loprinzi, C.L.; Lacchetti, C.; Bleeker, J.; Cavaletti, G.; Chauhan, C.; Hertz, D.L.; Kelley, M.R.; Lavino, A.; Lustberg, M.B.; Paice, J.A.; et al. Prevention and Management of Chemotherapy-Induced Peripheral Neuropathy in Survivors of Adult Cancers: ASCO Guideline Update. J. Clin. Oncol. 2020, 38, 3325–3348. [Google Scholar] [CrossRef]
- Brusco, I.; Silva, C.R.; Trevisan, G.; de Campos Velho Gewehr, C.; Rigo, F.K.; La Rocca Tamiozzo, L.; Rossato, M.F.; Tonello, R.; Dalmolin, G.D.; de Almeida Cabrini, D.; et al. Potentiation of Paclitaxel-Induced Pain Syndrome in Mice by Angiotensin I Converting Enzyme Inhibition and Involvement of Kinins. Mol. Neurobiol. 2017, 54, 7824–7837. [Google Scholar] [CrossRef]
- Costa, R.; Motta, E.M.; Dutra, R.C.; Manjavachi, M.N.; Bento, A.F.; Malinsky, F.R.; Pesquero, J.B.; Calixto, J.B. Anti-nociceptive Effect of Kinin B 1 and B 2 Receptor Antagonists on Peripheral Neuropathy Induced by Paclitaxel in Mice. Br. J. Pharmacol. 2011, 164, 681–693. [Google Scholar] [CrossRef] [Green Version]
- Bujalska, M.; Makulska-Nowak, H. Bradykinin Receptor Antagonists and Cyclooxygenase Inhibitors in Vincristine- and Streptozotocin-Induced Hyperalgesia. Pharmacol. Rep. 2009, 61, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Becker, G.; Fialho, M.F.P.; Brusco, I.; Oliveira, S.M. Kinin B1 and B2 Receptors Contribute to Cisplatin-Induced Painful Peripheral Neuropathy in Male Mice. Pharmaceutics 2023, 15, 852. [Google Scholar] [CrossRef] [PubMed]
- Shim, H.S.; Bae, C.; Wang, J.; Lee, K.-H.; Hankerd, K.M.; Kim, H.K.; Chung, J.M.; La, J.-H. Peripheral and Central Oxidative Stress in Chemotherapy-Induced Neuropathic Pain. Mol. Pain 2019, 15, 174480691984009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nassini, R.; Gees, M.; Harrison, S.; De Siena, G.; Materazzi, S.; Moretto, N.; Failli, P.; Preti, D.; Marchetti, N.; Cavazzini, A.; et al. Oxaliplatin Elicits Mechanical and Cold Allodynia in Rodents via TRPA1 Receptor Stimulation. Pain 2011, 152, 1621–1631. [Google Scholar] [CrossRef]
- Wotherspoon, G.; Winter, J. Bradykinin B1 Receptor Is Constitutively Expressed in the Rat Sensory Nervous System. Neurosci. Lett. 2000, 294, 175–178. [Google Scholar] [CrossRef]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like Channel Expressed in Nociceptive Neurons, Is Activated by Cold Temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Jordt, S.E.; Bautista, D.M.; Chuang, H.H.; McKemy, D.D.; Zygmunt, P.M.; Högestätt, E.D.; Meng, I.D.; Julius, D. Mustard Oils and Cannabinoids Excite Sensory Nerve Fibres through the TRP Channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef]
- Ma, Q.P. The Expression of Bradykinin B1 Receptors on Primary Sensory Neurones That Give Rise to Small Caliber Sciatic Nerve Fibres in Rats. Neuroscience 2001, 107, 665–673. [Google Scholar] [CrossRef]
- Ma, Q.P.; Heavens, R. Basal Expression of Bradykinin B1 Receptor in the Spinal Cord in Humans and Rats. Neuroreport 2001, 12, 2311–2314. [Google Scholar] [CrossRef]
- Brusco, I.; Fialho, M.F.P.; Becker, G.; Brum, E.S.; Favarin, A.; Marquezin, L.P.; Serafini, P.T.; Oliveira, S.M. Kinins and Their B1 and B2 Receptors as Potential Therapeutic Targets for Pain Relief. Life Sci. 2023, 314, 121302. [Google Scholar] [CrossRef]
- Calixto, Ä.B.; Cabrini, D.A.; Ferreira, J.; Campos, M.M. Kinins in Pain and Inflammation. Pain 2000, 87, 1–5. [Google Scholar] [CrossRef]
- Bautista, D.M.; Jordt, S.E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 Mediates the Inflammatory Actions of Environmental Irritants and Proalgesic Agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Dai, Y.; Fukuoka, T.; Yamanaka, H.; Kobayashi, K.; Obata, K.; Cui, X.; Tominaga, M.; Noguchi, K. Phospholipase C and Protein Kinase A Mediate Bradykinin Sensitization of TRPA1: A Molecular Mechanism of Inflammatory Pain. Brain 2008, 131, 1241–1251. [Google Scholar] [CrossRef] [Green Version]
- Andrade, E.L.; Meotti, F.C.; Calixto, J.B. TRPA1 Antagonists as Potential Analgesic Drugs. Pharmacol. Ther. 2012, 133, 189–204. [Google Scholar] [CrossRef]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious Cold Ion Channel TRPA1 Is Activated by Pungent Compounds and Bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Meotti, F.C.; Figueiredo, C.P.; Manjavachi, M.; Calixto, J.B. The Transient Receptor Potential Ankyrin-1 Mediates Mechanical Hyperalgesia Induced by the Activation of B 1 Receptor in Mice. Biochem. Pharmacol. 2017, 125, 75–83. [Google Scholar] [CrossRef]
- Chovanec, M.; Lauritsen, J.; Bandak, M.; Oing, C.; Kier, G.G.; Kreiberg, M.; Rosenvilde, J.; Wagner, T.; Bokemeyer, C.; Daugaard, G. Late Adverse Effects and Quality of Life in Survivors of Testicular Germ Cell Tumour. Nat. Rev. Urol. 2021, 18, 227–245. [Google Scholar] [CrossRef]
- Dolan, M.E.; El Charif, O.; Wheeler, H.E.; Gamazon, E.R.; Ardeshir-, S.; Monahan, P.; Feldman, D.R.; Hamilton, R.J.; David, J.; Beard, C.J.; et al. Clinical and Genome Wide Analysis of Cisplatin-Induced Peripheral Neuropathy in Survivors of Adult-Onset Cancer. Clin Cancer Res. 2018, 23, 5757–5768. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Du, W.; Acklin, S.; Jin, S.; Xia, F. SIRT2 Protects Peripheral Neurons from Cisplatin-Induced Injury by Enhancing Nucleotide Excision Repair. J. Clin. Investig. 2020, 130, 2953–2965. [Google Scholar] [CrossRef]
- Krukowski, K.; Ma, J.; Golonzhka, O.; Laumet, G.O.; Gutti, T.; Van Duzer, J.H.; Mazitschek, R.; Jarpe, M.B.; Heijnen, C.J.; Kavelaars, A. HDAC6 Inhibition Effectively Reverses Chemotherapy-Induced Peripheral Neuropathy. Pain 2017, 158, 1126–1137. [Google Scholar] [CrossRef]
- Zhang, J.; Junigan, J.M.; Trinh, R.; Kavelaars, A.; Heijnen, C.J.; Grace, P.M. HDAC6 Inhibition Reverses Cisplatin-Induced Mechanical Hypersensitivity via Tonic Delta Opioid Receptor Signaling. J. Neurosci. 2022, 42, 7862–7874. [Google Scholar] [CrossRef] [PubMed]
- Dewaeles, E.; Carvalho, K.; Fellah, S.; Sim, J.; Boukrout, N.; Caillierez, R.; Ramakrishnan, H.; Van der Hauwaert, C.; Vijaya Shankara, J.; Martin, N.; et al. Istradefylline Protects from Cisplatin-Induced Nephrotoxicity and Peripheral Neuropathy While Preserving Cisplatin Antitumor Effects. J. Clin. Investig. 2022, 132, e152924. [Google Scholar] [CrossRef] [PubMed]
- Khasabova, I.A.; Khasabov, S.G.; Olson, J.K.; Uhelski, M.L.; Kim, A.H.; Albino-Ramírez, A.M.; Wagner, C.L.; Seybold, V.S.; Simone, D.A. Pioglitazone, a PPARγ Agonist, Reduces Cisplatin-Evoked Neuropathic Pain by Protecting against Oxidative Stress. Pain 2019, 160, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Guindon, J.; Deng, L.; Fan, B.; Wager-miller, J.; Hohmann, A.G. Optimization of a Cisplatin Model of Chemotherapy- Induced Peripheral Neuropathy in Mice: Use of Vitamin C and Sodium Bicarbonate Pretreatments to Reduce Nephrotoxicity and Improve Animal Health Status. Mol. Pain 2014, 10, 56. [Google Scholar] [CrossRef] [Green Version]
- Bujalska, M.; Tatarkiewicz, J.; Gumułka, S.W. Effect of Bradykinin Receptor Antagonists on Vincristine- and Streptozotocin-Induced Hyperalgesia in a Rat Model of Chemotherapy-Induced and Diabetic Neuropathy. Pharmacology 2008, 81, 158–163. [Google Scholar] [CrossRef]
- Hori, K.; Ozaki, N.; Suzuki, S.; Sugiura, Y. Upregulations of P2X3 and ASIC3 Involve in Hyperalgesia Induced by Cisplatin Administration in Rats. Pain 2010, 149, 393–405. [Google Scholar] [CrossRef]
- Ta, L.E.; Bieber, A.J.; Carlton, S.M.; Loprinzi, C.L.; Low, P.A.; Windebank, A.J. Transient Receptor Potential Vanilloid 1 Is Essential for Cisplatin-Induced Heat Hyperalgesia in Mice. Mol. Pain 2010, 6, 1744–8069. [Google Scholar] [CrossRef] [Green Version]
- Vandewauw, I.; De Clercq, K.; Mulier, M.; Held, K.; Pinto, S.; Van Ranst, N.; Segal, A.; Voet, T.; Vennekens, R.; Zimmermann, K.; et al. A TRP Channel Trio Mediates Acute Noxious Heat Sensing. Nature 2018, 555, 662–666. [Google Scholar] [CrossRef]
- Fusi, C.; Materazzi, S.; Benemei, S.; Coppi, E.; Trevisan, G.; Marone, I.M.; Minocci, D.; De Logu, F.; Tuccinardi, T.; Di Tommaso, M.R.; et al. Steroidal and Non-Steroidal Third-Generation Aromatase Inhibitors Induce Pain-like Symptoms via TRPA1. Nat. Commun. 2014, 5, 5736. [Google Scholar] [CrossRef] [Green Version]
- Rossato, M.F.; Rigo, F.K.; Oliveira, S.M.; Guerra, G.P.; Silva, C.R.; Cunha, T.M.; Gomez, M.V.; Ferreira, J.; Trevisan, G. Participation of Transient Receptor Potential Vanilloid 1 in Paclitaxel-Induced Acute Visceral and Peripheral Nociception in Rodents. Eur. J. Pharmacol. 2018, 828, 42–51. [Google Scholar] [CrossRef]
- Brusco, I.; Benatti, A.; Regina, C.; Fischer, S.; Mattar, T.; Scussel, R.; Machado-de-ávila, R.A.; Ferreira, J.; Marchesan, S. Kinins and Their B 1 and B 2 Receptors Are Involved in Fibromyalgia-like Pain Symptoms in Mice. Biochem. Pharmacol. 2019, 168, 119–132. [Google Scholar] [CrossRef]
- Cross, L.J.M.; Heaney, L.G.; Ennis, M. Histamine Release from Human Bronchoalveolar Lavage Mast Cells by Neurokinin A and Bradykinin. Inflamm. Res. 1997, 46, 306–309. [Google Scholar] [CrossRef]
- Wei, C.C.; Hase, N.; Inoue, Y.; Bradley, E.W.; Yahiro, E.; Li, M.; Naqvi, N.; Powell, P.C.; Shi, K.; Takahashi, Y.; et al. Mast Cell Chymase Limits the Cardiac Efficacy of Ang I-Converting Enzyme Inhibitor Therapy in Rodents. J. Clin. Investig. 2010, 120, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Balemans, D.; Aguilera-Lizarraga, J.; Florens, M.V.; Jain, P.; Denadai-Souza, A.; Viola, M.F.; Alpizar, Y.A.; Van Der Merwe, S.; Vanden Berghe, P.; Talavera, K.; et al. Histamine-Mediated Potentiation of Transient Receptor Potential (TRP) Ankyrin 1 and TRP Vanilloid 4 Signaling in Submucosal Neurons in Patients with Irritable Bowel Syndrome. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G338–G349. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, C.; Wang, Z.J. Proteinase-Activated Receptor 2 Sensitizes Transient Receptor Potential Vanilloid 1, Transient Receptor Potential Vanilloid 4, and Transient Receptor Potential Ankyrin 1 in Paclitaxel-Induced Neuropathic Pain. Neuroscience 2011, 193, 440–451. [Google Scholar] [CrossRef]
- Costa, R.; Bicca, M.A.; Manjavachi, M.N.; Segat, G.C.; Dias, F.C.; Fernandes, E.S.; Calixto, J.B. Kinin Receptors Sensitize TRPV4 Channel and Induce Mechanical Hyperalgesia: Relevance to Paclitaxel-Induced Peripheral Neuropathy in Mice. Mol. Neurobiol. 2018, 55, 2150–2161. [Google Scholar] [CrossRef]
- Pethö, G.; Reeh, P.W. Sensory and Signaling Mechanisms of Bradykinin, Eicosanoids, Platelet-Activating Factor, and Nitric Oxide in Peripheral Nociceptors. Physiol. Rev. 2012, 92, 1699–1775. [Google Scholar] [CrossRef]
- Zhang, H.; Cang, C.-L.; Kawasaki, Y.; Liang, L.-L.; Zhang, Y.-Q.; Ji, R.-R.; Zhao, Z.-Q. Neurokinin-1 Receptor Enhances TRPV1 Activity in Primary Sensory Neurons via PKCε: A Novel Pathway for Heat Hyperalgesia. J. Neurosci. 2007, 27, 12067–12077. [Google Scholar] [CrossRef] [Green Version]
- Dina, O.A.; Chen, X.; Reichling, D.; Levine, J.D. Role of Protein Kinase Cε and Protein Kinase A in a Model of Paclitaxel-Induced Painful Peripheral Neuropathy in the Rat. Neuroscience 2001, 108, 507–515. [Google Scholar] [CrossRef]
- Honda, K.; Shinoda, M.; Kondo, M.; Shimizu, K.; Yonemoto, H.; Otsuki, K.; Akasaka, R.; Furukawa, A.; Iwata, K. Sensitization of TRPV1 and TRPA1 via Peripheral MGluR5 Signaling Contributes to Thermal and Mechanical Hypersensitivity. Pain 2017, 158, 1754–1764. [Google Scholar] [CrossRef]
- Cesare, P.; Dekker, L.V.; Sardini, A.; Parker, P.J.; McNaughton, P.A. Specific Involvement of PKC-ε in Sensitization of the Neuronal Response to Painful Heat. Neuron 1999, 23, 617–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.; Trichês, K.M.; Medeiros, R.; Calixto, J.B. Mechanisms Involved in the Nociception Produced by Peripheral Protein Kinase c Activation in Mice. Pain 2005, 117, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.; Trichês, K.M.; Medeiros, R.; Cabrini, D.A.; Mori, M.A.S.; Pesquero, J.B.; Bader, M.; Calixto, J.B. The Role of Kinin B1 Receptors in the Nociception Produced by Peripheral Protein Kinase C Activation in Mice. Neuropharmacology 2008, 54, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Rebecchi, M.J.; Pentyala, S.N. Structure, Function, and Control of Phosphoinositide-Specific Phospholipase C. Phisiological Rev. 2000, 80, 1291–1324. [Google Scholar] [CrossRef] [Green Version]
- de Almeida, A.S.; Rigo, F.K.; De Prá, S.D.-T.; Milioli, A.M.; Pereira, G.C.; Lückemeyer, D.D.; Antoniazzi, C.T.; Kudsi, S.Q.; Araújo, D.M.P.A.; Oliveira, S.M.; et al. Role of Transient Receptor Potential Ankyrin 1 (TRPA1) on Nociception Caused by a Murine Model of Breast Carcinoma. Pharmacol. Res. 2020, 152, 104576. [Google Scholar] [CrossRef]
- Fialho, M.F.P.; Brum, E.S.; Becker, G.; Brusco, I.; Oliveira, S.M. Kinin B2 and B1 Receptors Activation Sensitize the TRPA1 Channel Contributing to Anastrozole-Induced Pain Symptoms. Pharmaceutics 2023, 15, 1136. [Google Scholar] [CrossRef]
- McGrath, J.C.; Lilley, E. Implementing Guidelines on Reporting Research Using Animals (ARRIVE Etc.): New Requirements for Publication in BJP. Br. J. Pharmacol. 2015, 172, 3189–3193. [Google Scholar] [CrossRef] [Green Version]
- Ministério da Ciência, Tecnologia e Inovações. Resolução Normativa No 39, de 20 de Junho de 2018; Ministério da Ciência, Tecnologia e Inovações: Brasília, Brazil, 2018; Volume 120, p. 7.
- Zimmermann, M. Ethical Guidelines for Investigations of Experimental Pain in Conscious Animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Park, H.J.; Stokes, J.A.; Pirie, E.; Skahen, J.; Shtaerman, Y.; Yaksh, T.L. Persistent Hyperalgesia in the Cisplatin-Treated Mouse as Defined by Threshold Measures, the Conditioned Place Preference Paradigm, and Changes in Dorsal Root Ganglia Activated Transcription Factor 3: The Effects of Gabapentin, Ketorolac, and Etanercept. Anesth. Analg. 2013, 116, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Stokes, J.A.; Corr, M.; Yaksh, T.L. Toll-like Receptor Signaling Regulates Cisplatin-Induced Mechanical Allodynia in Mice. Cancer Chemother. Pharmacol. 2014, 73, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative Assessment of Tactile Allodynia in the Rat Paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Oliveira, S.M.; Silva, C.R.; Ferreira, J. Critical Role of Protease-Activated Receptor 2 Activation by Mast Cell Tryptase in the Development of Postoperative Pain. Anesthesiology 2013, 118, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Brum, E.d.S.; Fialho, M.F.P.; Fischer, S.P.M.; Hartmann, D.D.; Gonçalves, D.F.; Scussel, R.; Machado-de-Ávila, R.A.; Dalla Corte, C.L.; Soares, F.A.A.; Oliveira, S.M. Relevance of Mitochondrial Dysfunction in the Reserpine-Induced Experimental Fibromyalgia Model. Mol. Neurobiol. 2020, 57, 4202–4217. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, G.; Fialho, M.F.P.; Brum, E.S.; Oliveira, S.M. Kinin B2 Receptor Mediates Cisplatin-Induced Painful Peripheral Neuropathy by Intracellular Kinase Pathways and TRPA1 Channel Sensitisation. Pharmaceuticals 2023, 16, 959. https://doi.org/10.3390/ph16070959
Becker G, Fialho MFP, Brum ES, Oliveira SM. Kinin B2 Receptor Mediates Cisplatin-Induced Painful Peripheral Neuropathy by Intracellular Kinase Pathways and TRPA1 Channel Sensitisation. Pharmaceuticals. 2023; 16(7):959. https://doi.org/10.3390/ph16070959
Chicago/Turabian StyleBecker, Gabriela, Maria Fernanda Pessano Fialho, Evelyne Silva Brum, and Sara Marchesan Oliveira. 2023. "Kinin B2 Receptor Mediates Cisplatin-Induced Painful Peripheral Neuropathy by Intracellular Kinase Pathways and TRPA1 Channel Sensitisation" Pharmaceuticals 16, no. 7: 959. https://doi.org/10.3390/ph16070959
APA StyleBecker, G., Fialho, M. F. P., Brum, E. S., & Oliveira, S. M. (2023). Kinin B2 Receptor Mediates Cisplatin-Induced Painful Peripheral Neuropathy by Intracellular Kinase Pathways and TRPA1 Channel Sensitisation. Pharmaceuticals, 16(7), 959. https://doi.org/10.3390/ph16070959