
Analysis of Giardia lamblia Nucleolus as Drug Target: A Review


 and
and 
Abstract
:1. Introduction
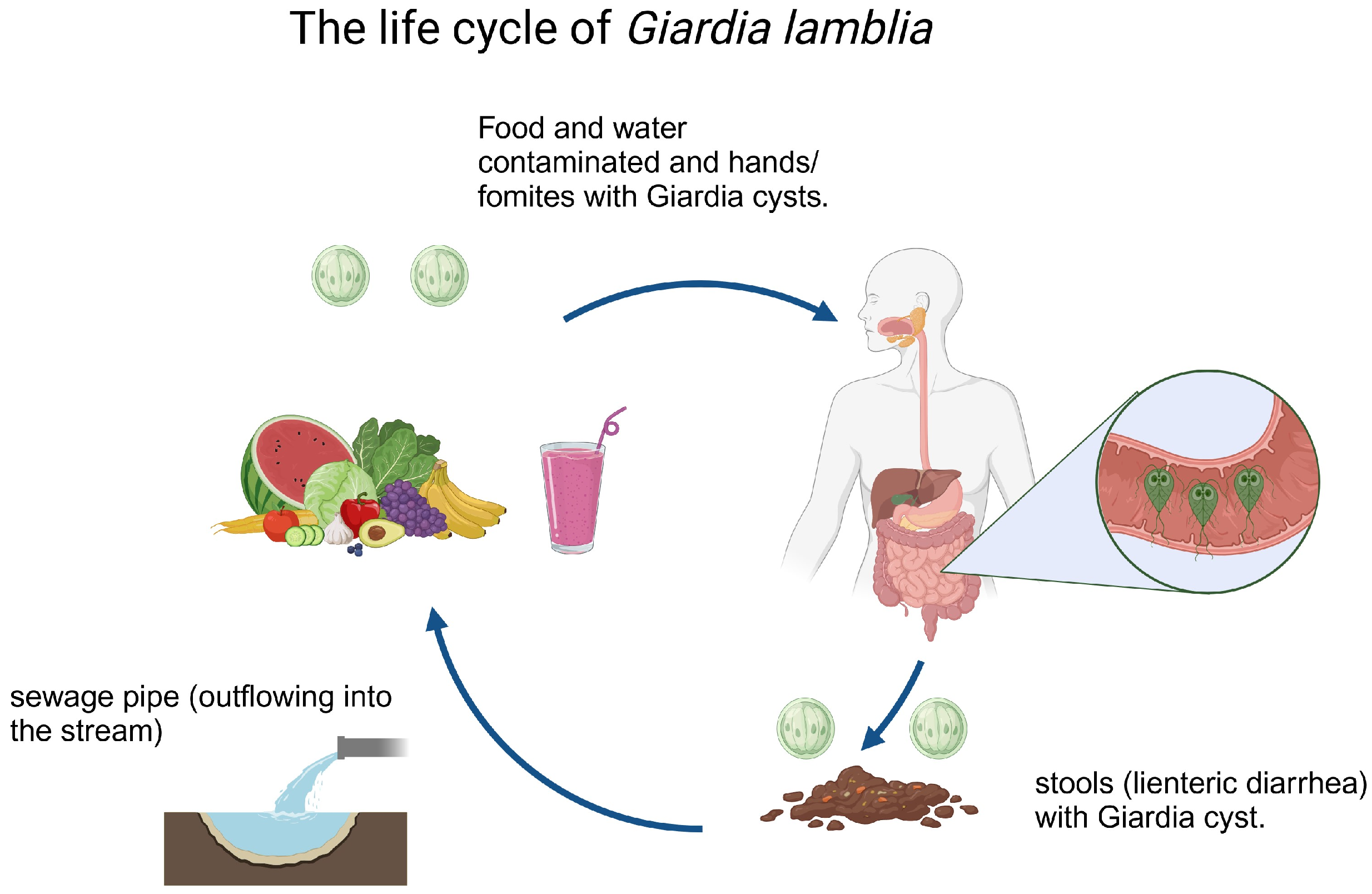
2. Cell Biology of G. lamblia
3. The Genome of G. lamblia
4. Nucleolus of G. lamblia
4.1. The Minimalist Nucleolus of G. lamblia as a Potential Therapeutic Target
4.2. G. lamblia Nucleolar Proteome
5. G. lamblia Has a Rudimentary Transcription Initiation Machinery
6. Drugs under Research for the Giardiasis
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, Regional, and National Causes of under-5 Mortality in 2000–15: An Updated Systematic Analysis with Implications for the Sustainable Development Goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef] [PubMed]
- Lanata, C.F.; Fischer-Walker, C.L.; Olascoaga, A.C.; Torres, C.X.; Aryee, M.J.; Black, R.E. Global Causes of Diarrheal Disease Mortality in Children <5 Years of Age: A Systematic Review. PLoS ONE 2013, 8, e72788. [Google Scholar] [CrossRef]
- Savioli, L.; Smith, H.; Thompson, A. Giardia and Cryptosporidium Join the ‘Neglected Diseases Initiative’. Trends Parasitol. 2006, 22, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Berkman, D.S.; Lescano, A.G.; Gilman, R.H.; Lopez, S.L.; Black, M.M. Effects of Stunting, Diarrhoeal Disease, and Parasitic Infection during Infancy on Cognition in Late Childhood: A Follow-up Study. Lancet 2002, 359, 564–571. [Google Scholar] [CrossRef]
- Correa, P.R. Factors Associated with Stunting among Children 0 to 59 Months of Age in Angola: A Cross-Sectional Study Using the 2015–2016 Demographic and Health Survey. PLoS Glob. Public Health 2022, 2, e0000983. [Google Scholar] [CrossRef]
- Júlio, C.; Vilares, A.; Oleastro, M.; Ferreira, I.; Gomes, S.; Monteiro, L.; Nunes, B.; Tenreiro, R.; Ângelo, H. Prevalence and Risk Factors for Giardia Duodenalis Infection among Children: A Case Study in Portugal. Parasites Vectors 2012, 5, 22. [Google Scholar] [CrossRef]
- Heyworth, M.F. Immunology of Giardia and Cryptosporidium Infections Martin. J. Infect. Dis. 1992, 166, 465–472. [Google Scholar] [CrossRef]
- Lipoldová, M. Giardia and Vilém Dušan Lambl. PLoS Negl. Trop. Dis. 2014, 8, e2686. [Google Scholar] [CrossRef]
- World Health Organization. Intestinal Protozoan and Helminthic Infections; World Health Organization Technical Report Series, No. 666; World Health Organization: Geneva, Switzerland, 1981; pp. 1–150. Available online: https://apps.who.int/iris/handle/10665/41519 (accessed on 1 July 2023).
- Hajare, S.T.; Chekol, Y.; Chauhan, N.M. Assessment of Prevalence of Giardia lamblia Infection and Its Associated Factors among Government Elementary School Children from Sidama Zone, SNNPR, Ethiopia. PLoS ONE 2022, 17, e0264812. [Google Scholar] [CrossRef]
- Hemphill, A.; Müller, N.; Müller, J. Comparative Pathobiology of the Intestinal Protozoan Parasites Giardia lamblia, Entamoeba histolytica, and Cryptosporidium parvum. Pathogens 2019, 8, 116. [Google Scholar] [CrossRef]
- Escobedo, A.A.; Cimerman, S. Giardiasis: A Pharmacotherapy Review. Expert Opin. Pharmacother. 2007, 8, 1885–1902. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.; Iqbal, W.; Adnan, F.; Wazir, S.; Khan, I.; Khayam, M.U.; Kamal, M.A.; Ahmad, S.; Ahmed, J.; Khan, I.N. Association of Metronidazole with Cancer: A Potential Risk Factor or Inconsistent Deductions? Curr. Drug Metab. 2018, 19, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Bendesky, A. Is Metronidazole Carcinogenic? Mutat. Res. 2002, 511, 133–144. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. IARC monographs on the evaluation of carcinogenic risks to humans. Ingested nitrate and nitrite, and cyanobacterial peptide toxins. IARC Monogr. Eval. Carcinog. Risks Hum. 2010, 94, 1–412. Available online: https://publications.iarc.fr/112 (accessed on 1 July 2023).
- Bradley, M.; Horton, J. Assessing the Risk of Benzimidazole Therapy during Pregnancy. Trans. R. Soc. Trop. Med. Hyg. 2001, 95, 72–73. [Google Scholar] [CrossRef] [PubMed]
- Tsiboukis, D.; Sazakli, E.; Jelastopulu, E.; Leotsinidis, M. Anthelmintics Residues in Raw Milk. Assessing Intake by a Children Population. Pol. J. Vet. Sci. 2013, 16, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Tang, S.; Chen, Q.; Zou, J.; Zhang, T.; Liu, F.; Zhang, S.; Sun, C.; Xiao, X. Furazolidone Induced Oxidative DNA Damage via Up-Regulating ROS That Caused Cell Cycle Arrest in Human Hepatoma G2 Cells. Toxicol. Lett. 2011, 201, 205–212. [Google Scholar] [CrossRef]
- Hussar, D.A. New Drugs of 2003. J. Am. Pharm. Assoc. 2004, 44, 168–210. [Google Scholar] [CrossRef]
- Watkins, R.R.; Eckmann, L. Treatment of Giardiasis: Current Status and Future Directions. Curr. Infect. Dis. Rep. 2014, 16, 396. [Google Scholar] [CrossRef]
- Singh, N.; Narayan, S. Nitazoxanide: A Broad Spectrum Antimicrobial. Med. J. Armed Forces India 2011, 67, 67–68. [Google Scholar] [CrossRef]
- Mayo Clinic Health System. Available online: https://www.mayoclinichealthsystem.org/ (accessed on 1 July 2023).
- Edwards, D.I. Mechanisms of Selective Toxicity of Metronidazole and Other Nitroimidazole Drugs. Sex. Transm. Infect. 1980, 56, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Dingsdag, S.A.; Hunter, N. Metronidazole: An Update on Metabolism, Structure–Cytotoxicity and Resistance Mechanisms. J. Antimicrob. Chemother. 2018, 73, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Movahedi, F.; Li, L.; Gu, W.; Xu, Z.P. Nanoformulations of Albendazole as Effective Anticancer and Antiparasite Agents. Nanomedicine 2017, 12, 2555–2574. [Google Scholar] [CrossRef] [PubMed]
- De Witt, M.; Gamble, A.; Hanson, D.; Markowitz, D.; Powell, C.; Al Dimassi, S.; Atlas, M.; Boockvar, J.; Ruggieri, R.; Symons, M. Repurposing Mebendazole as a Replacement for Vincristine for the Treatment of Brain Tumors. Mol. Med. 2017, 23, 50–56. [Google Scholar] [CrossRef]
- Meng, J.; Mangat, S.S.; Grudzinski, I.P.; Law, F.C.P. Evidence of 14C-Furazolidone Metabolite Binding to the Hepatic DNA of Trout. Drug Metab. Drug Interact. 1998, 14, 209–220. [Google Scholar] [CrossRef]
- Somvanshi, V.S.; Ellis, B.L.; Hu, Y.; Aroian, R.V. Nitazoxanide: Nematicidal Mode of Action and Drug Combination Studies. Mol. Biochem. Parasitol. 2014, 193, 1–8. [Google Scholar] [CrossRef]
- Norman, F.F.; Comeche, B.; Chamorro, S.; Pérez-Molina, J.-A.; López-Vélez, R. Update on the Major Imported Protozoan Infections in Travelers and Migrants. Future Microbiol. 2020, 15, 213–225. [Google Scholar] [CrossRef]
- Norman, F.F.; Monge-Maillo, B.; Martínez-Pérez, Á.; Perez-Molina, J.A.; López-Vélez, R. Parasitic Infections in Travelers and Immigrants: Part I Protozoa. Future Microbiol. 2015, 10, 69–86. [Google Scholar] [CrossRef]
- Müller, J.; Hemphill, A. Drug Target Identification in Protozoan Parasites. Expert Opin. Drug Discov. 2016, 11, 815–824. [Google Scholar] [CrossRef]
- Kourbeli, V.; Chontzopoulou, E.; Moschovou, K.; Pavlos, D.; Mavromoustakos, T.; Papanastasiou, I.P. An Overview on Target-Based Drug Design against Kinetoplastid Protozoan Infections: Human African Trypanosomiasis, Chagas Disease and Leishmaniases. Molecules 2021, 26, 4629. [Google Scholar] [CrossRef]
- Inge, P.M.; Edson, C.M.; Farthing, M.J. Attachment of Giardia lamblia to Rat Intestinal Epithelial Cells. Gut 1988, 29, 795–801. [Google Scholar] [CrossRef]
- LeChevallier, M.W.; Norton, W.D.; Lee, R.G. Giardia and Cryptosporidium Spp. in Filtered Drinking Water Supplies. Appl. Environ. Microbiol. 1991, 57, 2617–2621. [Google Scholar] [CrossRef] [PubMed]
- Jarroll, E.L.; Bingham, A.K.; Meyer, E.A. Effect of Chlorine on Giardia lamblia Cyst Viability. Appl. Environ. Microbiol. 1981, 41, 483–487. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Yee, J.; Bermúdez-Cruz, R.M. An Update on Cell Division of Giardia Duodenalis Trophozoites. Microbiol. Res. 2021, 250, 126807. [Google Scholar] [CrossRef]
- Gibson, W. The Sexual Side of Parasitic Protists. Mol. Biochem. Parasitol. 2021, 243, 111371. [Google Scholar] [CrossRef]
- Morrison, H.G.; McArthur, A.G.; Gillin, F.D.; Aley, S.B.; Adam, R.D.; Olsen, G.J.; Best, A.A.; Cande, W.Z.; Chen, F.; Cipriano, M.J.; et al. Genomic Minimalism in the Early Diverging Intestinal Parasite Giardia lamblia. Science 2007, 317, 1921–1926. [Google Scholar] [CrossRef] [PubMed]
- Adam, R.D. The Giardia lamblia Genome. Int. J. Parasitol. 2000, 30, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Yichoy, M.; Duarte, T.T.; De Chatterjee, A.; Mendez, T.L.; Aguilera, K.Y.; Roy, D.; Roychowdhury, S.; Aley, S.B.; Das, S. Lipid Metabolism in Giardia: A Post-Genomic Perspective. Parasitology 2011, 138, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Jiang, H.; Flores, R.; Wen, J. Gene Duplication in the Genome of Parasitic Giardia lamblia. BMC Evol. Biol. 2010, 10, 49. [Google Scholar] [CrossRef]
- Xue, M.; Chen, B.; Ye, Q.; Shao, J.; Lyu, Z.; Wen, J. Sense-Antisense Gene Overlap Is Probably a Cause for Retaining the Few Introns in Giardia Genome and the Implications. Biol. Direct 2018, 13, 23. [Google Scholar] [CrossRef]
- Best, A.A.; Morrison, H.G.; McArthur, A.G.; Sogin, M.L.; Olsen, G.J. Evolution of Eukaryotic Transcription: Insights from the Genome of Giardia lamblia. Genome Res. 2004, 14, 1537–1547. [Google Scholar] [CrossRef] [PubMed]
- Li, J. Characterization of Giardia Cell Nucleus: Its Implication on the Nature and Origin of the Primitive Cell Nucleus. Cell Res. 1995, 5, 115–124. [Google Scholar] [CrossRef]
- Islas-Morales, P.F.; Cárdenas, A.; Mosqueira, M.J.; Jiménez-García, L.F.; Voolstra, C.R. Ultrastructural and Proteomic Evidence for the Presence of a Putative Nucleolus in an Archaeon. Front. Microbiol. 2023, 14, 1075071. [Google Scholar] [CrossRef] [PubMed]
- Narcisi, E.M.; Glover, C.V.C.; Fechheimer, M. Fibrillarin, A Conserved Pre-Ribosomal RNA Processing Protein of Giardia. J. Eukaryot. Microbiol. 1998, 45, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kwon, I. Phase Transition of Fibrillarin LC Domain Regulates Localization and Protein Interaction of Fibrillarin. Biochem. J. 2021, 478, 799–810. [Google Scholar] [CrossRef]
- Shubina, M.Y.; Musinova, Y.R.; Sheval, E.V. Nucleolar Methyltransferase Fibrillarin: Evolution of Structure and Functions. Biochemistry 2016, 81, 941–950. [Google Scholar] [CrossRef]
- David, E.; McNeil, J.B.; Basile, V.; Pearlman, R.E. An Unusual Fibrillarin Gene and Protein: Structure and Functional Implications. Mol. Biol. Cell 1997, 8, 1051–1061. [Google Scholar] [CrossRef]
- Shubina, M.Y.; Arifulin, E.A.; Sorokin, D.V.; Sosina, M.A.; Tikhomirova, M.A.; Serebryakova, M.V.; Smirnova, T.; Sokolov, S.S.; Musinova, Y.R.; Sheval, E.V. The GAR Domain Integrates Functions That Are Necessary for the Proper Localization of Fibrillarin (FBL) inside Eukaryotic Cells. PeerJ 2020, 8, e9029. [Google Scholar] [CrossRef]
- Kolev, N.G.; Tschudi, C.; Ullu, E. RNA Interference in Protozoan Parasites: Achievements and Challenges. Eukaryot. Cell 2011, 10, 1156–1163. [Google Scholar] [CrossRef]
- Guo, J.; Chen, Y.; Zhou, K.; Li, J. Distribution of RDNA in the Nucleus of Giardia lamblia: Detection by Ag-I Silver Stain. Biotech. Histochem. 2005, 80, 31–34. [Google Scholar] [CrossRef]
- Gromadka, R.; Kaniak, A.; Slonimski, P.P.; Rytka, J. A Novel Cross-Phylum Family of Proteins Comprises a KRR1 (YCL059c) Gene Which Is Essential for Viability of Saccharomyces Cerevisiae Cells. Gene 1996, 171, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Xin, D.-D.; Wen, J.-F.; He, D.; Lu, S.-Q. Identification of a Giardia Krr1 Homolog Gene and the Secondarily Anucleolate Condition of Giaridia Lamblia. Mol. Biol. Evol. 2005, 22, 391–394. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Zhang, X.; Liu, Q.; Yin, J.; Yao, L.; Zhao, Y.; Cao, L. Inhibition of Krr1 Gene Expression in Giardia Canis by a Virus-Mediated Hammerhead Ribozyme. Vet. Parasitol. 2007, 143, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T. Small Nucleolar RNAs. Cell 2002, 109, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-Y.; Zhou, H.; Luo, J.; Qu, L.-H. Identification of 20 SnoRNA-like RNAs from the Primitive Eukaryote, Giardia lamblia. Biochem. Biophys. Res. Commun. 2005, 328, 1224–1231. [Google Scholar] [CrossRef]
- Saraiya, A.A.; Wang, C.C. SnoRNA, a Novel Precursor of MicroRNA in Giardia lamblia. PLoS Pathog. 2008, 4, e1000224. [Google Scholar] [CrossRef]
- Mei, Y.-P.; Liao, J.-P.; Shen, J.; Yu, L.; Liu, B.-L.; Liu, L.; Li, R.-Y.; Ji, L.; Dorsey, S.G.; Jiang, Z.-R.; et al. Small Nucleolar RNA 42 Acts as an Oncogene in Lung Tumorigenesis. Oncogene 2012, 31, 2794–2804. [Google Scholar] [CrossRef]
- Toden, S.; Zumwalt, T.J.; Goel, A. Non-Coding RNAs and Potential Therapeutic Targeting in Cancer. Biochim. Biophys. Acta (BBA) Rev. Cancer 2021, 1875, 188491. [Google Scholar] [CrossRef]
- Siprashvili, Z.; Webster, D.E.; Johnston, D.; Shenoy, R.M.; Ungewickell, A.J.; Bhaduri, A.; Flockhart, R.; Zarnegar, B.J.; Che, Y.; Meschi, F.; et al. The Noncoding RNAs SNORD50A and SNORD50B Bind K-Ras and Are Recurrently Deleted in Human Cancer. Nat. Genet. 2016, 48, 53–58. [Google Scholar] [CrossRef]
- Nyga, A.; Ganguli, S.; Matthews, H.K.; Baum, B. The Role of RAS Oncogenes in Controlling Epithelial Mechanics. Trends Cell Biol. 2023, 33, 60–69. [Google Scholar] [CrossRef]
- García-Huerta, E.; Espinoza-Corona, S.; Lagunas-Rangel, F.A.; Bazan-Tejeda, M.L.; Vazquez-Cobix, Y.; Ortega-Pierres, M.G.; Bermúdez-Cruz, R.M. Implementation of a Tunable T-CRISPRi System for Gene Regulation in Giardia Duodenalis. Plasmid 2022, 122, 102641. [Google Scholar] [CrossRef] [PubMed]
- McInally, S.G.; Hagen, K.D.; Nosala, C.; Williams, J.; Nguyen, K.; Booker, J.; Jones, K.; Dawson, S.C. Robust and Stable Transcriptional Repression in Giardia Using CRISPRi. Mol. Biol. Cell 2019, 30, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Buhrman, G.; Gagnon, K.; Mattos, C.; Brown, B.A.; Maxwell, E.S. Comparative Analysis of the 15.5kD Box C/D SnoRNP Core Protein in the Primitive Eukaryote Giardia lamblia Reveals Unique Structural and Functional Features. Biochemistry 2011, 50, 2907–2918. [Google Scholar] [CrossRef]
- Li, L.; Wang, C.C. Identification in the Ancient Protist Giardia lamblia of Two Eukaryotic Translation Initiation Factor 4E Homologues with Distinctive Functions. Eukaryot. Cell 2005, 4, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A.; Yee, J.; Bazán-Tejeda, M.L.; García-Villa, E.; Bermúdez-Cruz, R.M. Sirtuin GdSir2.4 Participates in the Regulation of RRNA Transcription in the Giardia Duodenalis Parasite. Mol. Microbiol. 2021, 115, 1039–1053. [Google Scholar] [CrossRef]
- Jimenez Garcia, L.; Zavala, G.; Chavez Munguia, B.; Ramos Godinez, M.; Lopez Velazquez, G.; Segura Valdez, M.; Montanez, C.; Hehl, A.; Arguello Garcia, R.; Ortega Pierres, G. Identification of Nucleoli in the Early Branching Protist Giardia duodenalis. Int. J. Parasitol. 2008, 38, 1297–1304. [Google Scholar] [CrossRef]
- Boisvert, F.-M.; van Koningsbruggen, S.; Navascués, J.; Lamond, A.I. The Multifunctional Nucleolus. Nat. Rev. Mol. Cell Biol. 2007, 8, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, P.; Pecoraro, A.; Palma, G.; Russo, G.; Russo, A. Therapeutic Approaches Targeting Nucleolus in Cancer. Cells 2019, 8, 1090. [Google Scholar] [CrossRef]
- Parlato, R.; Kreiner, G. Nucleolar Activity in Neurodegenerative Diseases: A Missing Piece of the Puzzle? J. Mol. Med. 2013, 91, 541–547. [Google Scholar] [CrossRef]
- Corman, A.; Sirozh, O.; Lafarga, V.; Fernandez-Capetillo, O. Targeting the Nucleolus as a Therapeutic Strategy in Human Disease. Trends Biochem. Sci. 2023, 48, 274–287. [Google Scholar] [CrossRef]
- Pickard, A.J.; Bierbach, U. The Cell’s Nucleolus: An Emerging Target for Chemotherapeutic Intervention. ChemMedChem 2013, 8, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.-M.; Yang, C.-L.; Tian, H.-F.; Wang, J.-X.; Wen, J.-F. Identification and Evolutionary Analysis of the Nucleolar Proteome of Giardia lamblia. BMC Genom. 2020, 21, 269. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A. Ribosomal RNA Transcription Machineries in Intestinal Protozoan Parasites: A Bioinformatic Analysis. Acta Parasitol. 2022, 67, 1788–1799. [Google Scholar] [CrossRef] [PubMed]
- Klug, A. Opening the Gateway. Nature 1993, 365, 486–487. [Google Scholar] [CrossRef]
- Kim, J.L.; Nikolov, D.B.; Burley, S.K. Co-Crystal Structure of TBP Recognizing the Minor Groove of a TATA Element. Nature 1993, 365, 520–527. [Google Scholar] [CrossRef]
- Hernandez, N. TBP, a Universal Eukaryotic Transcription Factor? Genes Dev. 1993, 7, 1291–1308. [Google Scholar] [CrossRef]
- Parra-Marín, O.; López-Pacheco, K.; Hernández, R.; López-Villaseñor, I. The Highly Diverse TATA Box-Binding Proteins among Protists: A Review. Mol. Biochem. Parasitol. 2020, 239, 111312. [Google Scholar] [CrossRef]
- Santiago, Á.; Razo-Hernández, R.S.; Pastor, N. The TATA-binding Protein DNA-binding Domain of Eukaryotic Parasites Is a Potentially Druggable Target. Chem. Biol. Drug Des. 2020, 95, 130–149. [Google Scholar] [CrossRef]
- Drummond, D.A.; Bloom, J.D.; Adami, C.; Wilke, C.O.; Arnold, F.H. Why Highly Expressed Proteins Evolve Slowly. Proc. Natl. Acad. Sci. USA 2005, 102, 14338–14343. [Google Scholar] [CrossRef]
- Bensaude, O. Inhibiting Eukaryotic Transcription. Which Compound to Choose? How to Evaluate Its Activity? Transcription 2011, 2, 103–108. [Google Scholar] [CrossRef]
- Sogin, M.L.; Gunderson, J.H.; Elwood, H.J.; Alonso, R.A.; Peattie, D.A. Phylogenetic Meaning of the Kingdom Concept: An Unusual Ribosomal RNA from Giardia lamblia. Science 1989, 243, 75–77. [Google Scholar] [CrossRef]
- Jordan, P.; Carmo-Fonseca, M. Cisplatin Inhibits Synthesis of Ribosomal RNA in Vivo. Nucleic Acids Res. 1998, 26, 2831–2836. [Google Scholar] [CrossRef]
- Zisi, A.; Bartek, J.; Lindström, M.S. Targeting Ribosome Biogenesis in Cancer: Lessons Learned and Way Forward. Cancers 2022, 14, 2126. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A. The Nucleolus of Giardia and Its Ribosomal Biogenesis. Parasitol. Res. 2023, 122, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Available online: https://www.clinicaltrials.gov/ (accessed on 6 August 2023).
- Mhashilkar, A.S.; Vankayala, S.L.; Liu, C.; Kearns, F.; Mehrotra, P.; Tzertzinis, G.; Palli, S.R.; Woodcock, H.L.; Unnasch, T.R. Identification of Ecdysone Hormone Receptor Agonists as a Therapeutic Approach for Treating Filarial Infections. PLoS Negl. Trop. Dis. 2016, 10, e0004772. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Drug | Mechanism of Action | Side Effects [22] |
|---|---|---|
 Benzoylmetronidazole | Prevents the production of genetic material. DNA strand rupture. Potential suppression of DNA repair [23,24]. | Fever, headache, irritability, lack of coordination, nausea, blurred vision. |
 Metronidazole | Prevents the production of genetic material. DNA strand rupture. Potential suppression of DNA repair [23,24]. | Fever, headache, irritability, lack of coordination, nausea, blurred vision. |
 Ornidazole | Prevents the production of genetic material. DNA strand rupture. Potential suppression of DNA repair [23,24]. | Fever, headache, irritability, lack of coordination, nausea, blurred vision. |
 Tinidazole | Prevents the production of genetic material. DNA strand rupture. Potential suppression of DNA repair [23,24]. | Bitter taste, metallic taste, difficulty having a bowel movement, dizziness, general feeling of discomfort or illness. |
 Albendazole | Tubulin assembly suppressor. Fumarate reductase suppressor. VEGF and HIF-1α suppressor. Halt the progression of both the G2 and M phases within the cell cycle [25]. | Stomach pain, chest pain, chills, fever. |
 Mebendazole | Tubulin assembly suppressor. Inhibiting glucose absorption [26]. | Abdominal or stomach pain or upset, Black, tarry stools, chills. |
 Furazolidone | It is suggested to work through DNA cross-linkage [27]. | Joint pain, fever, itching, skin rash or redness, sore throat. |
 Nitazoxanide | Blocks the activity of pyruvate: ferredoxin/flavodoxin oxidoreductases. Suppression of protein disulfide isomerase activity. Possible binding to nitroreductase GlNR1. Modulation of the expression of heat-shock proteins [28]. | Abdominal or stomach pain, headache, nausea, urine changes. |
| Therapeutic Target | Therapeutic Strategy | Function |
|---|---|---|
| Fibrillarin | Target-based drug design and gene-silencing therapies. | rRNA processing and maturation. |
| Krr1 | Target-based drug design and gene-silencing therapies. | Assembly 40S ribosomal subunit. |
| snoRNP (15.5kD Box C/D) | Target-based drug design. | Methylation and Pseudouridylation of rRNA. |
| GdSir2.4 | Target-based drug design. | Deacetylase (modulation rRNA genes expression) |
| GlTBP | Target-based drug design. | Essential to form PIC. |
| eIF4E1 | Target-based drug design. | Putative methylation and Pseudouridylation of rRNA. |
| snoRNA (GlsR3, GlsR9, GlsR10 and GlsR11) | Gene-silencing therapies. | Methylation and Pseudouridylation of rRNA. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaona-López, C.; Martínez-Vázquez, A.V.; Villalobos-Rocha, J.C.; Juárez-Rendón, K.J.; Rivera, G. Analysis of Giardia lamblia Nucleolus as Drug Target: A Review. Pharmaceuticals 2023, 16, 1168. https://doi.org/10.3390/ph16081168
Gaona-López C, Martínez-Vázquez AV, Villalobos-Rocha JC, Juárez-Rendón KJ, Rivera G. Analysis of Giardia lamblia Nucleolus as Drug Target: A Review. Pharmaceuticals. 2023; 16(8):1168. https://doi.org/10.3390/ph16081168
Chicago/Turabian StyleGaona-López, Carlos, Ana Verónica Martínez-Vázquez, Juan Carlos Villalobos-Rocha, Karina Janett Juárez-Rendón, and Gildardo Rivera. 2023. "Analysis of Giardia lamblia Nucleolus as Drug Target: A Review" Pharmaceuticals 16, no. 8: 1168. https://doi.org/10.3390/ph16081168
APA StyleGaona-López, C., Martínez-Vázquez, A. V., Villalobos-Rocha, J. C., Juárez-Rendón, K. J., & Rivera, G. (2023). Analysis of Giardia lamblia Nucleolus as Drug Target: A Review. Pharmaceuticals, 16(8), 1168. https://doi.org/10.3390/ph16081168