

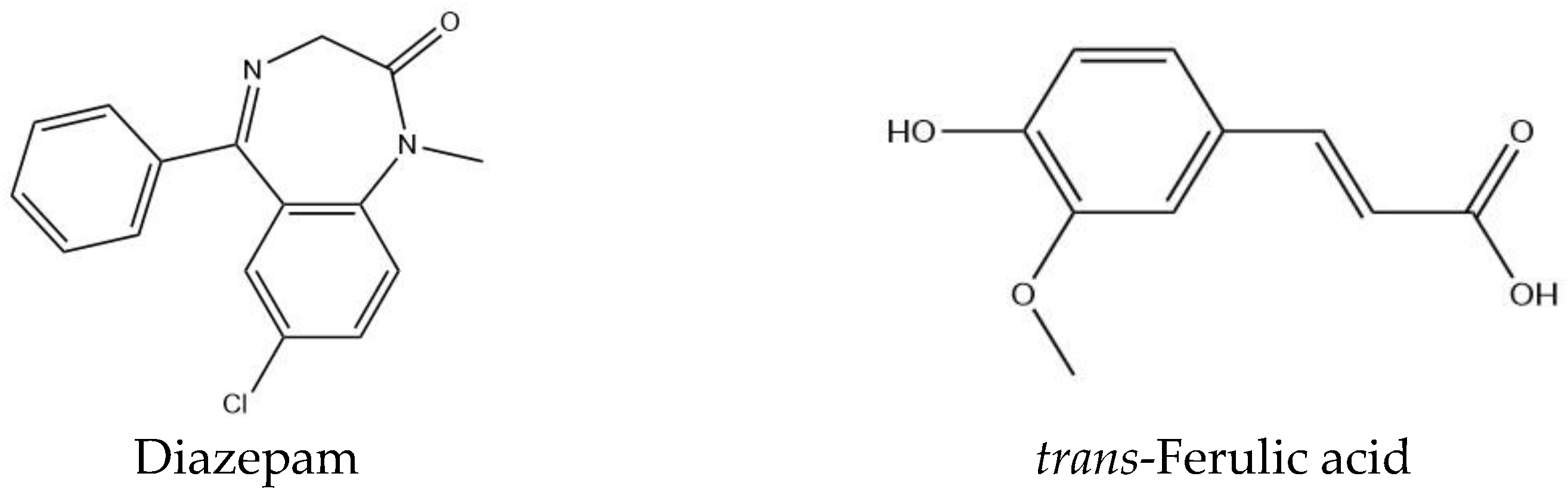
Anxiolytic-like Effects by trans-Ferulic Acid Possibly Occur through GABAergic Interaction Pathways
 ,
,  , , , ,
, , , ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. In Vivo Studies
2.1.1. Open Field Study
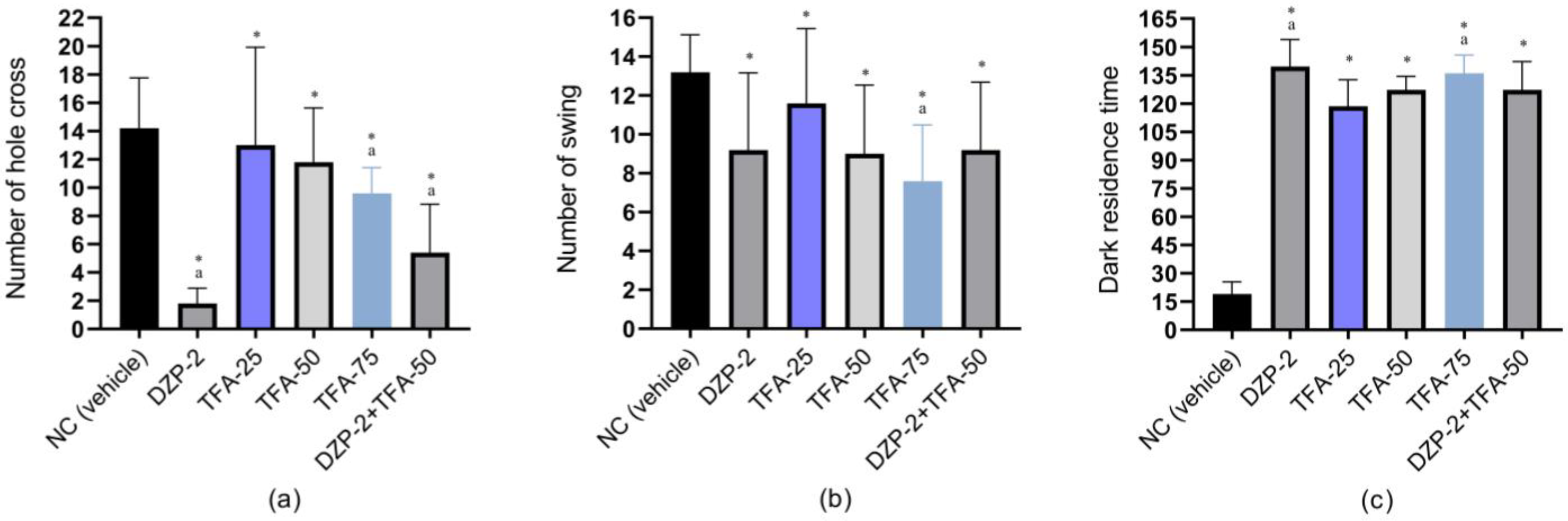
2.1.2. Hole Cross Study
2.1.3. Swing Study
2.1.4. Dark–Light Study
2.2. In Silico Study
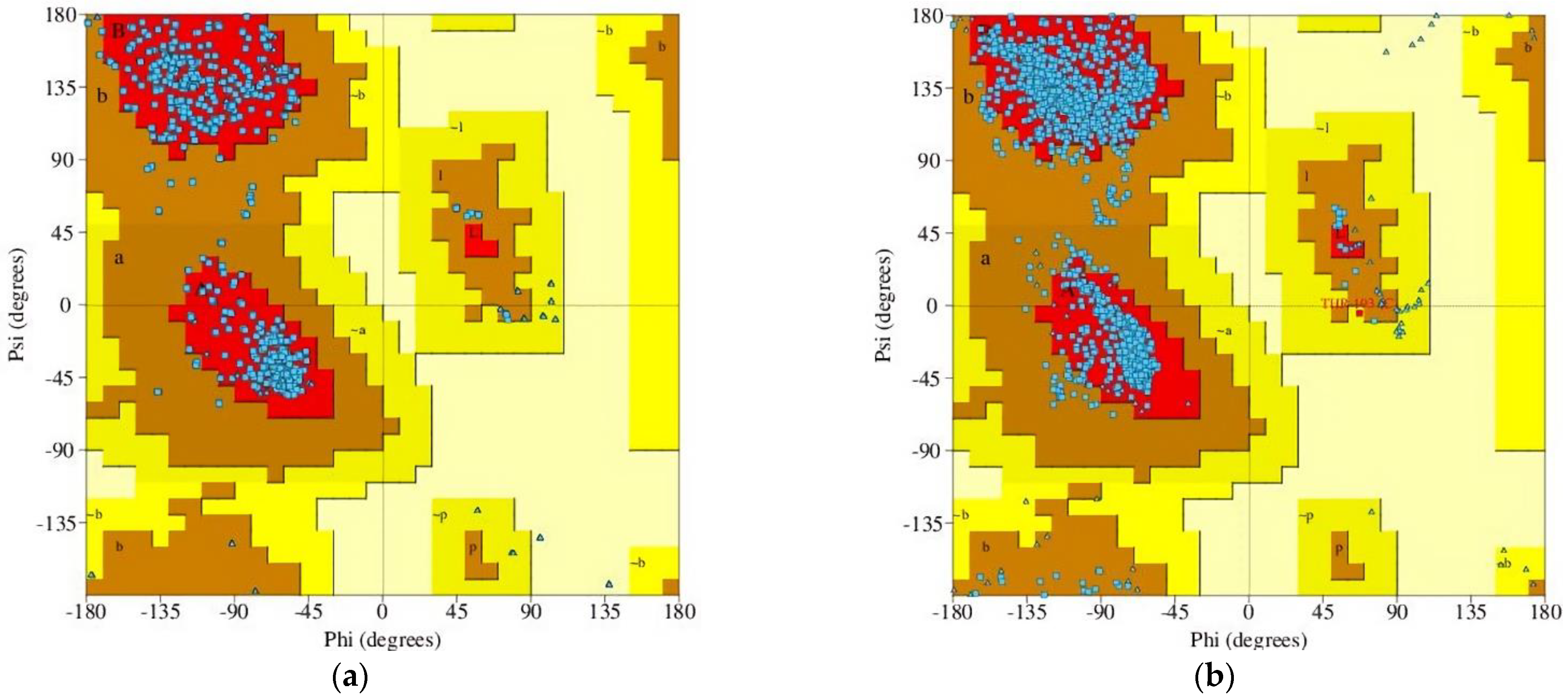
2.2.1. GABAA Receptor Homology Modeling
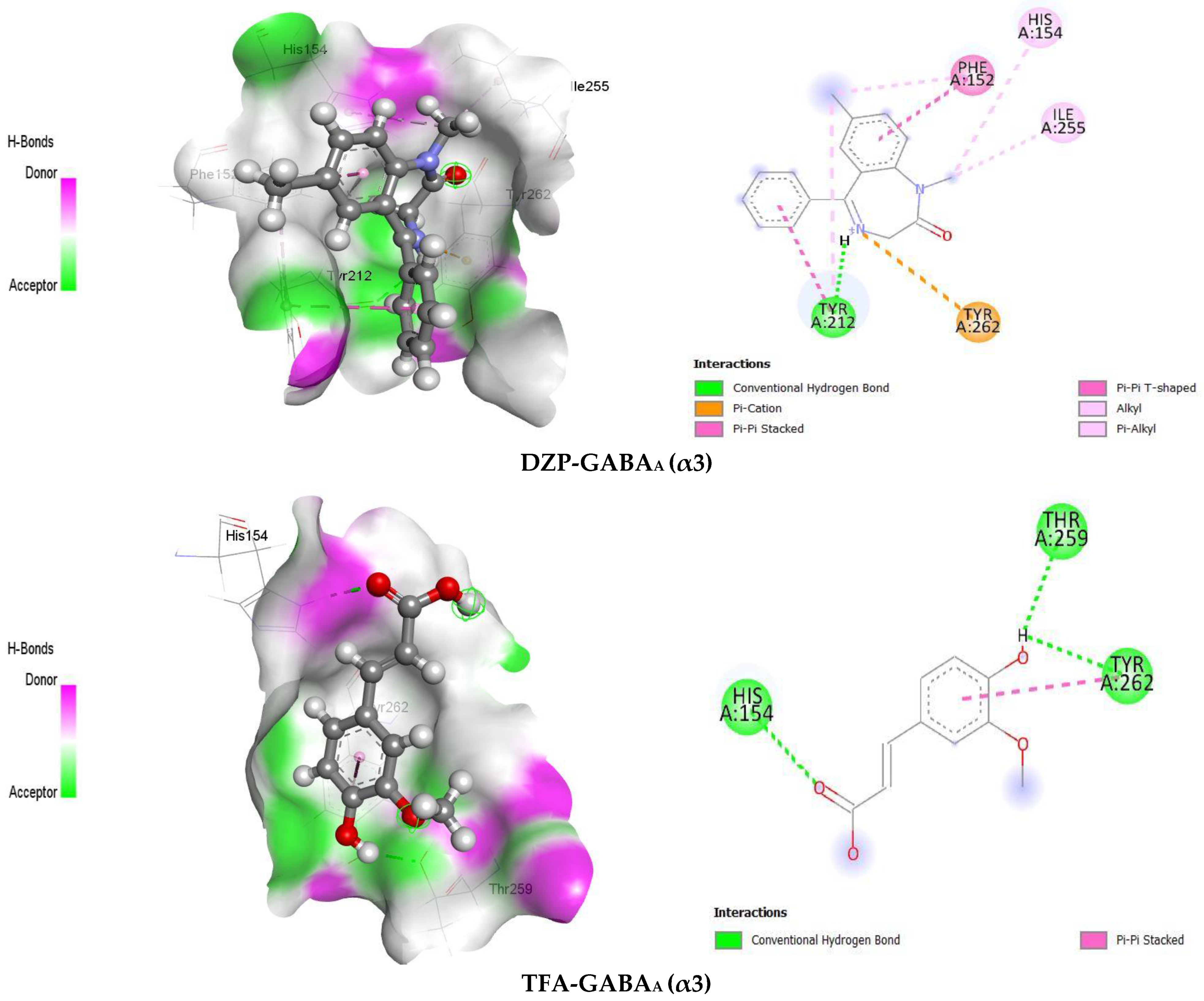
2.2.2. Molecular Docking and Visualization of Ligand–Receptor Interactions
3. Discussion
4. Materials and Methods
4.1. In Vivo Study
4.1.1. Reagents and Chemicals
4.1.2. Preparation of Test and Referral Drugs
4.1.3. Experimental Animals
4.1.4. Study Design
Open Field Test (OFT)
Hole Cross Test (HCT)
Swing Test (ST)
Dark–Light Test (DLT)
4.1.5. Statistical Analysis
4.2. In Silico Study
4.2.1. Homology Modeling and Preparation of Receptors
4.2.2. Collection and Preparation of Ligands
4.2.3. Molecular Docking and Visualization of Ligand–Receptor Interactions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tiller, J.W. Depression and anxiety. Med. J. Aust. 2013, 199, S28–S31. [Google Scholar] [CrossRef] [PubMed]
- Bhuia, M.S.; Rahaman, M.M.; Islam, T.; Bappi, M.H.; Sikder, M.I.; Hossain, K.N.; Akter, F.; Al Shamsh Prottay, A.; Rokonuzzman, M.; Gürer, E.S.; et al. Neurobiological effects of gallic acid: Current perspectives. Chin. Med. 2023, 18, 27. [Google Scholar] [CrossRef] [PubMed]
- Pahl, K.M.; Barrett, P.M.; Gullo, M.J. Examining Potential Risk Factors for Anxiety in Early Childhood. J. Anxiety Disord. 2012, 26, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Rende, R.; Waldman, I.J. Behavioral and molecular genetics and developmental psychopathology. Dev. Psychopathol. 2015, 10, 427–464. [Google Scholar]
- Khan, M.M.A.; Khan, M.N. Effects of Psychosocial and Socio-Environmental Factors on Anxiety Disorder among Adolescents in Bangladesh. Brain Behav. 2020, 10, e01899. [Google Scholar] [CrossRef]
- Haenisch, B.; Bönisch, H. Depression and Antidepressants: Insights from Knockout of Dopamine, Serotonin or Noradrenaline Re-Uptake Transporters. Pharmacol. Ther. 2011, 129, 352–368. [Google Scholar] [CrossRef] [PubMed]
- Di Giovanni, G.; Esposito, E.; Di Matteo, V. Role of Serotonin in Central Dopamine Dysfunction. CNS Neurosci. Ther. 2010, 16, 179–194. [Google Scholar] [CrossRef]
- Davies, R.D.; Gabbert, S.L.; Riggs, P.D. Anxiety Disorders in Neurologic Illness. Curr. Treat. Options Neurol. 2001, 3, 333–346. [Google Scholar] [CrossRef]
- Wang, H.Q.; Wang, Z.Z.; Chen, N.H. The receptor hypothesis and the pathogenesis of depression: Genetic bases and biological correlates. Pharmacol. Res. 2021, 167, 105542. [Google Scholar] [CrossRef]
- Möhler, H. The GABA system in anxiety and depression and its therapeutic potential. Neuropharmacology 2012, 62, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Żmudzka, E.; Sałaciak, K.; Sapa, J.; Pytka, K. Serotonin Receptors in Depression and Anxiety: Insights from Animal Studies. Life Sci. 2018, 210, 106–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, W.; Li, T.; Guo, Y.; Tian, Y.; Wang, F.; Liu, S.; Shen, H.Y.; Feng, Y.; Xiao, L. Impairment of Oligodendroglia Maturation Leads to Aberrantly Increased Cortical Glutamate and Anxiety-Like Behaviors in Juvenile Mice. Front. Cell. Neurosci. 2015, 9, 467. [Google Scholar] [CrossRef]
- McKernan, R.M.; Rosahl, T.W.; Reynolds, D.S.; Sur, C.; Wafford, K.A.; Atack, J.R.; Farrar, S.; Myers, J.; Cook, G.; Ferris, P.; et al. Sedative but not anxiolytic properties of benzodiazepines are mediated by the GABA(A) receptor alpha1 subtype. Nat. Neurosci. 2000, 3, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Löw, K.; Crestani, F.; Keist, R.; Benke, D.; Brünig, I.; Benson, J.A.; Fritschy, J.M.; Rülicke, T.; Bluethmann, H.; Möhler, H.; et al. Molecular and neuronal substrate for the selective attenuation of anxiety. Science 2000, 290, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Rowlett, J.K.; Platt, D.M.; Lelas, S.; Atack, J.R.; Dawson, G.R. Different GABAA receptor subtypes mediate the anxiolytic, abuse-related, and motor effects of benzodiazepine-like drugs in primates. Proc. Natl. Acad. Sci. USA 2005, 102, 915–920. [Google Scholar] [CrossRef]
- Kim, D.H.; Kwon, H.; Choi, J.W.; Shin, C.Y.; Cheong, J.H.; Park, S.J.; Ryu, J.H. Roles of GABAA Receptor α5 Subunit on Locomotion and Working Memory in Transient Forebrain Ischemia in Mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 102, 109962. [Google Scholar] [CrossRef]
- Behlke, L.M.; Foster, R.A.; Liu, J.; Benke, D.; Benham, R.S.; Nathanson, A.J.; Yee, B.K.; Zeilhofer, H.U.; Engin, E.; Rudolph, U. A Pharmacogenetic ‘Restriction-of-Function’ Approach Reveals Evidence for Anxiolytic-Like Actions Mediated by α5-Containing GABAA Receptors in Mice. Neuropsychopharmacology 2016, 41, 2492–2501. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y. Neuropsychological Mechanism Underlying Antidepressant Effect: A Systematic Meta-Analysis. Mol. Psychiatry 2015, 20, 311–319. [Google Scholar] [CrossRef]
- Pfeiffer, P.N.; Ganoczy, D.; Zivin, K.; Valenstein, M. Benzodiazepines and Adequacy of Initial Antidepressant Treatment for Depression. J. Clin. Psychopharmacol. 2011, 31, 360–364. [Google Scholar] [CrossRef]
- Genario, R.; Giacomini, A.; de Abreu, M.S.; Marcon, L.; Demin, K.A.; Kalueff, A.V. Sex differences in adult zebrafish anxiolytic-like responses to diazepam and melatonin. Neurosci. Lett. 2020, 714, 134548. [Google Scholar] [CrossRef] [PubMed]
- López-Muñoz, F.; Alamo, C.; García-García, P. The discovery of chlordiazepoxide and the clinical introduction of benzodiazepines: Half a century of anxiolytic drugs. J. Anxiety Disord. 2011, 25, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Balon, R.; Starcevic, V. Role of Benzodiazepines in Anxiety Disorders. Adv. Exp. Med. Biol. 2020, 1191, 367–388. [Google Scholar] [CrossRef]
- Rezaeiroshan, A.; Saeedi, M.; Morteza-Semnani, K.; Akbari, J.; Hedayatizadeh-Omran, A.; Goli, H.; Nokhodchi, A. Vesicular formation of trans-ferulic acid: An efficient approach to improve the radical scavenging and antimicrobial properties. J. Pharm. Investig. 2022, 17, 652–661. [Google Scholar] [CrossRef]
- Trombino, S.; Cassano, R.; Bloise, E.; Muzzalupo, R.; Tavano, L.; Picci, N. Synthesis and antioxidant activity evaluation of a novel cellulose hydrogel containing trans-ferulic acid. Carbohydr. Polym. 2009, 75, 184–188. [Google Scholar] [CrossRef]
- Trombino, S.; Cassano, R.; Ferrarelli, T.; Barone, E.; Picci, N.; Mancuso, C. Trans-ferulic acid-based solid lipid nanoparticles and their antioxidant effect in rat brain microsomes. Colloids Surf. B Biointerfaces 2013, 109, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Suh, J.H.; Zheng, X.; Wang, Y.; Ho, C.T. Identification and Quantification of Potential Anti-inflammatory Hydroxycinnamic Acid Amides from Wolfberry. J. Agric. Food Chem. 2017, 65, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Helmy, S.A.; El-Mofty, S.; El Gayar, A.M.; El-Sherbiny, I.M.; El-Far, Y.M. Novel doxorubicin/folate-targeted trans-ferulic acid-loaded PLGA nanoparticles combination: In-vivo superiority over standard chemotherapeutic regimen for breast cancer treatment. Biomed. Pharmacother. 2022, 145, 112376. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, E.H.M.; Abdel-Wahab, B.A.; Ali, F.E.M.; Abd El-Ghafar, O.A.M.; Kozman, M.R.; Sharkawi, S.M.Z. Trans-ferulic acid ameliorates cisplatin-induced testicular damage via suppression of TLR4, P38-MAPK, and ERK1/2 signaling pathways. Environ. Sci. Pollut. Res. Int. 2021, 28, 41948–41964. [Google Scholar] [CrossRef] [PubMed]
- Shayan-Nasr, M.; Ghaniei, A.; Eslami, M.; Zadeh-Hashem, E. Ameliorative role of trans-ferulic acid on induced oxidative toxicity of rooster semen by β-cyfluthrin during low temperature liquid storage. Poult. Sci. 2021, 100, 101308. [Google Scholar] [CrossRef]
- Chotimarkorn, C.; Ushio, H. The effect of trans-ferulic acid and gamma-oryzanol on ethanol-induced liver injury in C57BL mouse. Phytomedicine 2008, 15, 951–958. [Google Scholar] [CrossRef]
- Singh, S.S.B.; Patil, K.N. Trans-ferulic acid attenuates hyperglycemia-induced oxidative stress and modulates glucose metabolism by activating AMPK signaling pathway in vitro. J. Food Biochem. 2022, 46, e14038. [Google Scholar] [CrossRef]
- Zafeer, M.F.; Firdaus, F.; Anis, E.; Mobarak Hossain, M. Prolong treatment with Trans-ferulic acid mitigates bioenergetics loss and restores mitochondrial dynamics in streptozotocin-induced sporadic dementia of Alzheimer’s type. Neurotoxicology 2019, 73, 246–257. [Google Scholar] [CrossRef]
- Nagarajan, S.; Chellappan, D.R.; Chinnaswamy, P.; Thulasingam, S. Ferulic acid pretreatment mitigates MPTP-induced motor impairment and histopathological alterations in C57BL/6 mice. Pharm. Biol. 2015, 53, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Lorigooini, Z.; Nouri, A.; Mottaghinia, F.; Balali-Dehkordi, S.; Bijad, E.; Dehkordi, S.H.; Soltani, A.; Amini-Khoei, H. Ferulic acid through mitigation of NMDA receptor pathway exerts anxiolytic-like effect in mouse model of maternal separation stress. J. Basic Clin. Physiol. Pharmacol. 2020, 32, 1–10. [Google Scholar] [CrossRef]
- Chen, J.; Lin, D.; Zhang, C.; Li, G.; Zhang, N.; Ruan, L.; Yan, Q.; Li, J.; Yu, X.; Xie, X.; et al. Antidepressant-like effects of ferulic acid: Involvement of serotonergic and norepinergic systems. Metab. Brain Dis. 2015, 30, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.; Kaur, T.; Goel, R.K. Ferulic Acid Supplementation for Management of Depression in Epilepsy. Neurochem. Res. 2017, 42, 2940–2948. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Shi, A.M.; Wang, Q. Sedative-hypnotic and anxiolytic effects and the mechanism of action of aqueous extracts of peanut stems and leaves in mice. J. Sci. Food Agric. 2018, 98, 4885–4894. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Hayashida, H.; Murata, M.; Takamatsu, J. Effect of ferulic acid and Angelica archangelica extract on behavioral and psychological symptoms of dementia in frontotemporal lobar degeneration and dementia with Lewy bodies. Geriatr. Gerontol. Int. 2011, 11, 309–314. [Google Scholar] [CrossRef]
- Koh, P.O. Ferulic acid prevents the cerebral ischemic injury-induced decrease of Akt and Bad phosphorylation. Neurosci. Lett. 2012, 507, 156–160. [Google Scholar] [CrossRef]
- Dhaliwal, J.; Dhaliwal, N.; Akhtar, A.; Kuhad, A.; Chopra, K. Beneficial effects of ferulic acid alone and in combination with insulin in streptozotocin induced diabetic neuropathy in Sprague Dawley rats. Life Sci. 2020, 255, 117856. [Google Scholar] [CrossRef]
- Xu, Y.; Lin, D.; Yu, X.; Xie, X.; Wang, L.; Lian, L.; Fei, N.; Chen, J.; Zhu, N.; Wang, G.; et al. The antinociceptive effects of ferulic acid on neuropathic pain: Involvement of descending monoaminergic system and opioid receptors. Oncotarget 2016, 7, 20455–20468. [Google Scholar] [CrossRef]
- Salau, V.F.; Erukainure, O.L.; Ibeji, C.U.; Olasehinde, T.A.; Koorbanally, N.A.; Islam, M.S. Ferulic Acid Modulates Dysfunctional Metabolic Pathways and Purinergic Activities, While Stalling Redox Imbalance and Cholinergic Activities in Oxidative Brain Injury. Neurotoxic. Res. 2020, 37, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Moon, N.R.; Kang, S.; Kim, D.S. Ferulic acid and vinpocetine intake improves memory function by enhancing insulin sensitivity and reducing neuroinflammation and oxidative stress in type 2 diabetic animals with induced Alzheimer’s disease. J. Funct. Foods 2022, 95, 105180. [Google Scholar] [CrossRef]
- Ghobadi, M.; Arji, B.; Yadegari, M.; Esmailidehaj, M.; Homayouni Moghadam, F.; Rezvani, M.E. Ferulic Acid Ameliorates Cell Injuries, Cognitive and Motor Impairments in Cuprizone-Induced Demyelination Model of Multiple Sclerosis. Cell J. 2022, 24, 681–688. [Google Scholar] [CrossRef]
- Araki, R.; Yasubuchi, A.; Ikegaya, M.; Hojo, C.; Tachioka, H.; Kawai, K.; Omote, M.; Kita, A.; Yabe, T. Ferulic acid alleviates abnormal behaviors in isolation-reared mice via 5-HT(1A) receptor partial agonist activity. Psychopharmacology 2021, 238, 2147–2154. [Google Scholar] [CrossRef]
- Sborgi, S.M.S.; Fernandes, L.C.; Santos, A.G.; Ferro, M.M.; Miyoshi, E.J.R. Anxiolytic Activity of Ferulic Acid in the Light-Dark Test in Zebrafish. Society 2021, 10, e582101119894. [Google Scholar] [CrossRef]
- Sliwoski, G.; Kothiwale, S.; Meiler, J.; Lowe, E.W., Jr. Computational Methods in Drug Discovery. Pharmaceutics 2014, 66, 334–395. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. Biotech R&D spend jumps by more than 15. Nat. Rev. Drug Discov. 2016, 15, 447. [Google Scholar] [CrossRef]
- Schaduangrat, N.; Lampa, S.; Simeon, S.; Gleeson, M.P.; Spjuth, O.; Nantasenamat, C. Towards reproducible computational drug discovery. J. Cheminform. 2020, 12, 9. [Google Scholar] [CrossRef]
- Tibbitts, J.; Canter, D.; Graff, R.; Smith, A.; Khawli, L.A. Key Factors Influencing ADME Properties of Therapeutic Proteins: A Need for ADME Characterization in Drug Discovery and Development. mAbs 2016, 8, 229–245. [Google Scholar] [CrossRef]
- Bhuia, M.S.; Islam, T.; Rokonuzzman, M.; Shamsh Prottay, A.A.; Akter, F.; Hossain, M.I.; Chowdhury, R.; Kazi, M.A.; Khalipha, A.B.R.; Coutinho, H.D.M.; et al. Modulatory Effects of Phytol on the Antiemetic Property of Domperidone, Possibly through the D2 Receptor Interaction Pathway: In Vivo and In Silico Studies. 3 Biotech 2023, 13, 116. [Google Scholar] [CrossRef] [PubMed]
- Bhuia, M.S.; Wilairatana, P.; Ferdous, J.; Chowdhury, R.; Bappi, M.H.; Rahman, M.A.; Mubarak, M.S.; Islam, M.T. Hirsutine, an Emerging Natural Product with Promising Therapeutic Benefits: A Systematic Review. Molecules 2023, 28, 6141. [Google Scholar] [CrossRef]
- Bhuia, M.S.; Chowdhury, R.; Sonia, F.A.; Kamli, H.; Shaikh, A.; El-Nashar, H.A.S.; El-Shazly, M.; Islam, M.T. Anticancer Potential of the Plant-Derived Saponin Gracillin: A Comprehensive Review of Mechanistic Approaches. Chem. Biodivers. 2023, 8, e202300847. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, N.M.; Gesto, D.; Oliveira, E.F.; Santos-Martins, D.; Brás, N.F.; Sousa, S.F.; Fernandes, P.A.; Ramos, M.J. Receptor-Based Virtual Screening Protocol for Drug Discovery. Arch. Biochem. Biophys. 2015, 582, 56–67. [Google Scholar] [CrossRef] [PubMed]
- MacIntyre, M.M.; Zare, M.; Williams, M.T. Anxiety-Related Disorders in the Context of Racism. Curr. Psychiatry Rep. 2023, 25, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Bourin, M. Animal models for screening anxiolytic-like drugs: A perspective. Dialogues Clin. Neurosci. 2015, 17, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Craske, M.G.; Rauch, S.L.; Ursano, R.; Prenoveau, J.; Pine, D.S.; Zinbarg, R.E. What is an anxiety disorder? Depress. Anxiety 2009, 26, 1066–1085. [Google Scholar] [CrossRef]
- Steimer, T. Animal models of anxiety disorders in rats and mice: Some conceptual issues. Dialogues Clin. Neurosci. 2011, 13, 495–506. [Google Scholar] [CrossRef]
- Julian, L.J. Measures of anxiety: State-Trait Anxiety Inventory (STAI), Beck Anxiety Inventory (BAI), and Hospital Anxiety and Depression Scale-Anxiety (HADS-A). Arthritis Care Res. 2011, 63 (Suppl. S11), S467–S472. [Google Scholar] [CrossRef]
- Fuchs, E.; Flügge, G. Experimental Animal Models for the Simulation of Depression and Anxiety. Dialogues Clin. Neurosci. 2006, 8, 323–333. [Google Scholar] [CrossRef]
- Thompson, T.; Grabowski-Boase, L.; Tarantino, L.M. Prototypical anxiolytics do not reduce anxiety-like behavior in the open field in C57BL/6J mice. Pharmacol. Biochem. Behav. 2015, 133, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.S.; Ibrahim, M.; Rahman, M.M.; Islam, T.; Bhuia, M.S.; Zaman, S.; Islam, M.T. Phytochemical group test and pharmacological investigations of Persicaria barbata (L.) H. Hara. Pharm. Res. J. 2023, 2, 1–15. [Google Scholar]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to Measure Locomotor and Anxiety-Like Behavior in Mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef]
- Amaral, M.P.M.; da Silva, M.P., Jr.; da Lima, F.C.A.; Gutierrez, S.J.C.; Arcanjo, D.D.R.; de Oliveira, R.C.M. Anxiolytic/Sedative Effect of Monoterpene (−)-Borneol in Mice and In Silico Molecular Interaction with GABAA Receptor. Pharmaceutics 2023, 3, 132–141. [Google Scholar] [CrossRef]
- Aman, U.; Subhan, F.; Shahid, M.; Akbar, S.; Ahmad, N.; Ali, G.; Fawad, K.; Sewell, R.D. Passiflora incarnata attenuation of neuropathic allodynia and vulvodynia apropos GABA-ergic and opioidergic antinociceptive and behavioural mechanisms. BMC Complement. Altern. Med. 2016, 16, 77. [Google Scholar] [CrossRef]
- Islam, N.U.; Khan, I.; Rauf, A.; Muhammad, N.; Shahid, M.; Shah, M.R. Antinociceptive, muscle relaxant and sedative activities of gold nanoparticles generated by methanolic extract of Euphorbia milii. BMC Complement. Altern. Med. 2015, 15, 160. [Google Scholar] [CrossRef]
- Shastry, R.; Ullal, S.D.; Karkala, S.; Rai, S.; Gadgade, A. Anxiolytic activity of aqueous extract of Camellia sinensis in rats. Indian J. Pharmacol. 2016, 48, 681–686. [Google Scholar] [CrossRef]
- Knabl, J.; Zeilhofer, U.B.; Crestani, F.; Rudolph, U.; Zeilhofer, H.U. Genuine Antihyperalgesia by Systemic Diazepam Revealed by Experiments in GABAA Receptor Point-Mutated Mice. Pain 2009, 141, 233–238. [Google Scholar] [CrossRef]
- Bernal-Morales, B.; Cueto-Escobedo, J.; Guillén-Ruiz, G.; Rodríguez-Landa, J.F.; Contreras, C.M. A Fatty Acids Mixture Reduces Anxiety-Like Behaviors in Infant Rats Mediated by GABA(A) Receptors. BioMed Res. Int. 2017, 2017, 8798546. [Google Scholar] [CrossRef]
- Reimus, P.W.; Dangelmayr, M.A.; Clay, J.T.; Chamberlain, K.R. Uranium Natural Attenuation Downgradient of an In Situ Recovery Mine Inferred from a Cross-Hole Field Test. Environ. Sci. Technol. 2019, 53, 7483–7493. [Google Scholar] [CrossRef]
- Islam, M.T.; Martins, N.; Imran, M.; Hameed, A.; Ali, S.W.; Salehi, B.; Ahmad, I.; Hussain, A.; Sharifi-Rad, J. Anxiolytic-like effects of Moringa oleifera in Swiss mice. Cell. Mol. Biol. 2020, 66, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Hossain, R.; Ahmed, T.; Rahaman, M.M.; Al-Khafaji, K.; Khan, R.A.; Sarkar, C.; Bappi, M.H.; de Andrade, E.M.; Araújo, I.M.; et al. Anxiolytic-like Effect of Quercetin Possibly through GABA Receptor Interaction Pathway: In Vivo and In Silico Studies. Molecules 2022, 27, 7149. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.; Rana, S.; Islam, M.; Kumer, A.; Hassan, M.; Biswas, T.K.; Atikullah, M. Evaluation of anxiolytic and sedative-like activities of methanolic extract of Euphorbia hirta leaves in mice. Pharm. J. Pharm. 2019, 10, 283–297. [Google Scholar] [CrossRef]
- Arrant, A.E.; Schramm-Sapyta, N.L.; Kuhn, C.M. Use of the light/dark test for anxiety in adult and adolescent male rats. Behav. Brain Res. 2013, 256, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Riaz, T.A.; Ayatollahi, S.A.; Sharifi-Rad, J. Anxiolytic-like effect of Urena lobata (L.) in Swiss albino mice. Cureus 2021, 7, e1–e6. [Google Scholar] [CrossRef]
- Sandeep, K.; Rajmeet, S.; Poonam, A. Pharmacological studies on the protective effect of ferulic acid in monosodium glutamate-induced anxiety and motor incoordination. J. Clin. Pharm. 2018, 7, 208–214. [Google Scholar] [CrossRef]
- Nuss, P. Anxiety disorders and GABA neurotransmission: A disturbance of modulation. Neuropsychiatr. Dis. Treat. 2015, 11, 165–175. [Google Scholar] [CrossRef]
- Nutt, D.J.; Malizia, A.L. New insights into the role of the GABA(A)-benzodiazepine receptor in psychiatric disorder. Br. J. Psychiatry 2001, 179, 390–396. [Google Scholar] [CrossRef]
- Calcaterra, N.E.; Barrow, J.C. Classics in chemical neuroscience: Diazepam (valium). ACS Chem. Neurosci. 2014, 5, 253–260. [Google Scholar] [CrossRef]
- Andrade, J.C.; Monteiro, Á.B.; Andrade, H.H.N.; Gonzaga, T.; Silva, P.R.; Alves, D.N.; Castro, R.D.; Maia, M.S.; Scotti, M.T.; Sousa, D.P.; et al. Involvement of GABA(A) Receptors in the Anxiolytic-Like Effect of Hydroxycitronellal. BioMed Res. Int. 2021, 2021, 9929805. [Google Scholar] [CrossRef]
- Sollozo-Dupont, I.; Estrada-Camarena, E.; Carro-Juárez, M.; López-Rubalcava, C. GABAA/benzodiazepine receptor complex mediates the anxiolytic-like effect of Montanoa tomentosa. J. Ethnopharmacol. 2015, 162, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Tallarida, R.J. Quantitative methods for assessing drug synergism. Genes Cancer 2011, 2, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Lerche, P.; Muir, W.W., III. Pain management in horses and cattle. In Handbook of Veterinary Pain Management; Elsevier: Amsterdam, The Netherlands, 2009; pp. 437–466. [Google Scholar]
- Wu, M.; Sirota, M.; Butte, A.J.; Chen, B. Characteristics of drug combination therapy in oncology by analyzing clinical trial data on ClinicalTrials.gov. Pac. Symp. Biocomput. 2015, 2014, 68–79. [Google Scholar]
- Jin, W.; Stokes, J.M.; Eastman, R.T.; Itkin, Z.; Zakharov, A.V.; Collins, J.J.; Jaakkola, T.S.; Barzilay, R. Deep learning identifies synergistic drug combinations for treating COVID-19. Proc. Natl. Acad. Sci. USA 2021, 118, e2105070118. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Sanderson, P.E.; Zheng, W. Drug combination therapy increases successful drug repositioning. Drug Discov. Today 2016, 21, 1189–1195. [Google Scholar] [CrossRef]
- Brogi, S.; Ramalho, T.C.; Kuca, K.; Medina-Franco, J.L.; Valko, M. Editorial: In silico Methods for Drug Design and Discovery. Front. Chem. 2020, 8, 612. [Google Scholar] [CrossRef]
- Pinzi, L.; Rastelli, G. Molecular Docking: Shifting Paradigms in Drug Discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef]
- Schmidtke, P.; Luque, F.J.; Murray, J.B.; Barril, X. Shielded Hydrogen Bonds as Structural Determinants of Binding Kinetics: Application in Drug Design. J. Am. Chem. Soc. 2011, 133, 18903–18910. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar] [CrossRef]
- Subhan, N.; Alam, M.A.; Ahmed, F.; Shahid, I.J.; Nahar, L.; Sarker, S.D. Bioactivity of Excoecaria agallocha. Res. Bull. Develop. 2008, 18, 521–526. [Google Scholar] [CrossRef]
- Islam, M.T.; Freitas, R.; Oliveira, G.; Guha, B. Neuropharmacological screenings of hydroalcoholic fractions of Urena lobata L. World J. Pharm. Pharm. Sci. 2014, 3, 62–71. [Google Scholar]
- Gong, Z.-H.; Li, Y.-F.; Zhao, N.; Yang, H.-J.; Su, R.-B.; Luo, Z.-P.; Li, J. Anxiolytic effect of agmatine in rats and mice. Eur. J. Pharmacol. 2006, 550, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.W.; Prlić, A.; Altunkaya, A.; Bi, C.; Bradley, A.R.; Christie, C.H.; Costanzo, L.D.; Duarte, J.M.; Dutta, S.; Feng, Z.; et al. The RCSB Protein Data Bank: Integrative view of protein, gene and 3D structural information. Nucleic Acids Res. 2016, 27, gkw1000. [Google Scholar] [CrossRef]
- Bordoli, L.; Kiefer, F.; Arnold, K.; Benkert, P.; Battey, J.; Schwede, T. Protein structure homology modeling using SWISS-MODEL workspace. Nat. Protoc. 2009, 4, 1–13. [Google Scholar] [CrossRef]
- McGarvey, P.B.; Nightingale, A.; Luo, J.; Huang, H.; Martin, M.J.; Wu, C.; UniProt Consortium. UniProt genomic mapping for deciphering functional effects of missense variants. Nucleic Acids Res. 2019, 40, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Boratyn, G.M.; Schäffer, A.A.; Agarwala, R.; Altschul, S.F.; Lipman, D.J.; Madden, T.L. Domain Enhanced Lookup Time Accelerated BLAST. Biol. Direct 2012, 7, 12. [Google Scholar] [CrossRef]
- Shi, M.; Wang, L.; Li, P.; Liu, J.; Chen, L.; Xu, D. Dasatinib–SIK2 Binding Elucidated by Homology Modeling, Molecular Docking, and Dynamics Simulations. ACS Omega 2021, 6, 11025–11038. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Jabłońska, J.; Pravda, L.; Vařeková, R.S.; Thornton, J.M. PDBsum: Structural summaries of PDB entries. Protein Sci. 2018, 27, 129–134. [Google Scholar] [CrossRef]
- Selvam, K.; Senbagam, D.; Selvankumar, T.; Sudhakar, C.; Kamala-Kannan, S.; Senthilkumar, B.; Govarthanan, M. Cellulase Enzyme: Homology Modeling, Binding Site Identification and Molecular Docking. J. Mol. Struct. 2017, 1150, 61–67. [Google Scholar] [CrossRef]
- Gundampati, R.K.; Chikati, R.; Kumari, M.; Sharma, A.; Pratyush, D.D.; Jagannadham, M.V.; Kumar, C.S.; Debnath Das, M. Protein-Protein Docking on Molecular Models of Aspergillus niger RNase and Human Actin: Novel Target for Anticancer Therapeutics. J. Mol. Model. 2012, 18, 653–662. [Google Scholar] [CrossRef]
- Lill, M.A.; Danielson, M.L. Computer-aided drug design platform using PyMOL. J. Comput. Aided Mol. Des. 2011, 25, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, S.; Vijayakumar, S.; Manogar, P.; Maniam, G.P.; Govindan, N. Homology modeling and molecular docking studies on type II diabetes complications reduced PPARγ receptor with various ligand molecules. Biomed. Pharmacother. 2017, 92, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Bhuia, M.S.; Kamli, H.; Islam, T.; Sonia, F.A.; Kazi, M.A.; Siam, M.S.H.; Rahman, N.; Bappi, M.H.; Mia, M.N.; Hossen, M.M.; et al. Antiemetic Activity of Trans-Ferulic Acid Possibly through Muscarinic Receptors Interaction Pathway: In Vivo and In Silico Study. Results Chem. 2023, 6, 101014. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Abdelrahman, A.H.; Badr, E.A.; Almansour, N.M.; Alzahrani, O.R.; Ahmed, M.N.; Soliman, M.E.; Naeem, M.A.; Shawky, A.M.; Sidhom, P.A. Naturally occurring plant-based anticancerous candidates as prospective ABCG2 inhibitors: An in silico drug discovery study. J. Mol. Graph. Model. 2022, 26, 3255–3277. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligands | Receptors | Docking Scores (kcal/mol) | Amino Acid Residues | |
|---|---|---|---|---|
| HB | Others | |||
| DZP | GABAA (α2) | −5.9 | SER134, GLY131, PHE128 | MET158, PHE127, LEU160 |
| TFA | GABAA (α2) | −5.8 | SER186, THR234, SER232 | TYR237 |
| DZP | GABAA (α3) | −7.0 | TYR212 | TYR262, TYR212, PHE152, ILE255, HIS154 |
| TFA | GABAA (α3) | −5.3 | HIS154, THR259, TYR262 | TYR262 |
| Treatment Groups | Description (R/A) | Dose (mg/kg) |
|---|---|---|
| Gr.-I: NC | Vehicle: Distilled water (p.o) | 10 |
| Gr.-II: DZP | Standard: Diazepam (agonist) (p.o) | 2 |
| Gr.-III: TFA | Lower dose (p.o) | 25 |
| Gr.-IV: TFA | Medium dose (p.o) | 50 |
| Gr.-V: TFA | Higher dose (p.o) | 75 |
| Gr.-VI: TFA + DZP | Test + Standard combination (p.o) | 50 + 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhuia, M.S.; Rokonuzzman, M.; Hossain, M.I.; Ansari, S.A.; Ansari, I.A.; Islam, T.; Al Hasan, M.S.; Mubarak, M.S.; Islam, M.T. Anxiolytic-like Effects by trans-Ferulic Acid Possibly Occur through GABAergic Interaction Pathways. Pharmaceuticals 2023, 16, 1271. https://doi.org/10.3390/ph16091271
Bhuia MS, Rokonuzzman M, Hossain MI, Ansari SA, Ansari IA, Islam T, Al Hasan MS, Mubarak MS, Islam MT. Anxiolytic-like Effects by trans-Ferulic Acid Possibly Occur through GABAergic Interaction Pathways. Pharmaceuticals. 2023; 16(9):1271. https://doi.org/10.3390/ph16091271
Chicago/Turabian StyleBhuia, Md. Shimul, Md. Rokonuzzman, Md. Imran Hossain, Siddique Akber Ansari, Irfan Aamer Ansari, Tawhida Islam, Md. Sakib Al Hasan, Mohammad S. Mubarak, and Muhammad Torequl Islam. 2023. "Anxiolytic-like Effects by trans-Ferulic Acid Possibly Occur through GABAergic Interaction Pathways" Pharmaceuticals 16, no. 9: 1271. https://doi.org/10.3390/ph16091271
APA StyleBhuia, M. S., Rokonuzzman, M., Hossain, M. I., Ansari, S. A., Ansari, I. A., Islam, T., Al Hasan, M. S., Mubarak, M. S., & Islam, M. T. (2023). Anxiolytic-like Effects by trans-Ferulic Acid Possibly Occur through GABAergic Interaction Pathways. Pharmaceuticals, 16(9), 1271. https://doi.org/10.3390/ph16091271