Relationships between Cargo, Cell Penetrating Peptides and Cell Type for Uptake of Non-Covalent Complexes into Live Cells


 and
and
Abstract
:1. Introduction

2. Results and Discussion
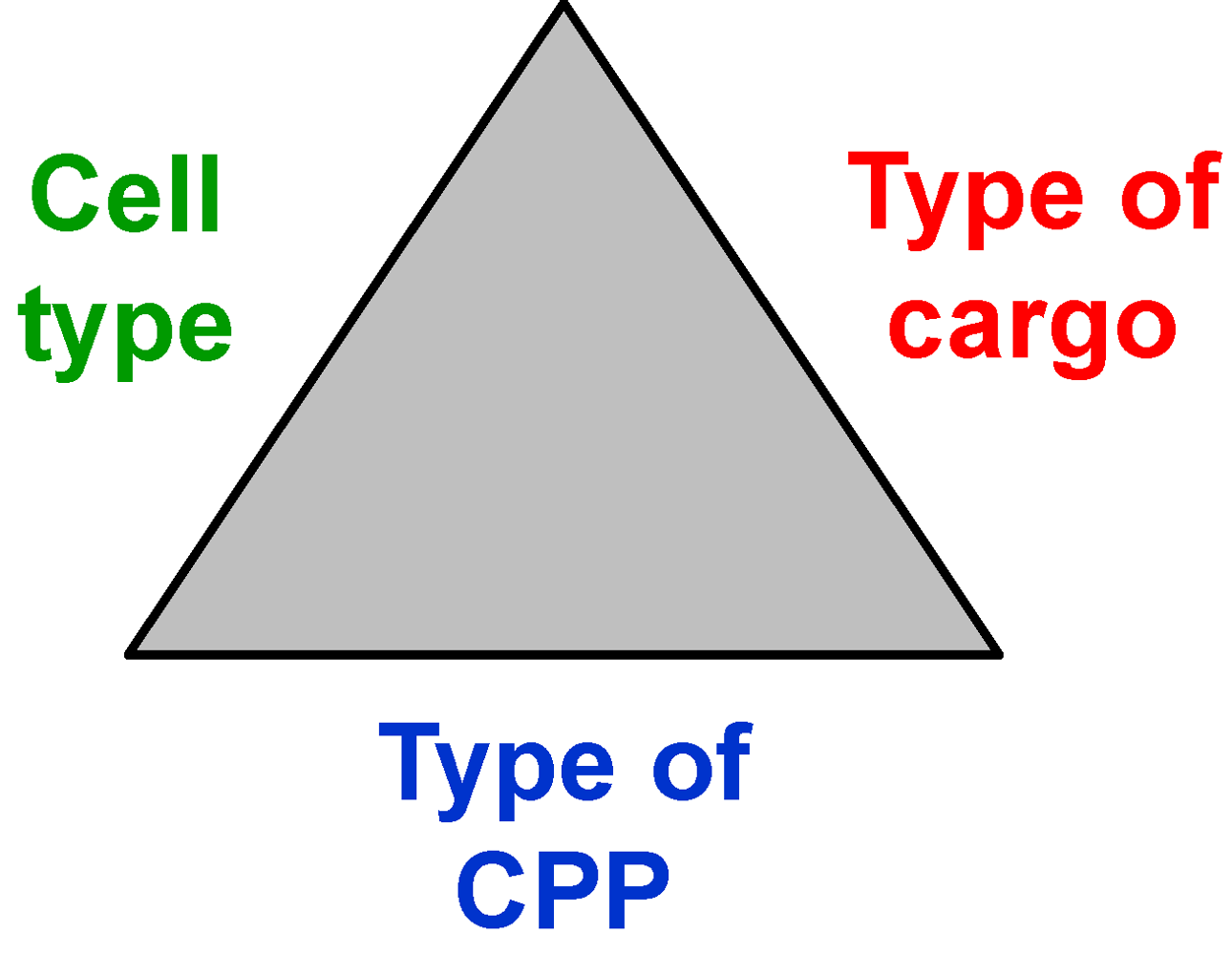
2.1. Cell Penetrating Peptides, Cargoes and Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amphiphilic-cationic peptides (transcription factors and -activators) | ||
|---|---|---|
| Penetratin (Antennapedia) | RQIKIWFQNRRMKWKK | 8 positive charges, MW 2247 Da |
| HIV-TAT (47-57) | YGRKKRRQRRR-amide | 9 positive charges, MW 1560 Da |
| MPG peptides | ||
| MPGα | Ac-GALFLAFLAAALSLMGLWSQPKKKRKV-NH-CH2-CH2-SH | 5 positive charges, MW 3047 Da |
| MPGβ | Ac-GALFLGFLGAAGSTMGAWSQPKKKRKV-NH-CH2-CH2-SH | 5 positive charges, MW 2910 Da |
| CAD-2 (des-acetyl, Lys19-CADY) | GLWRALWRLLRSLWRLLWKA-NH-CH2-CH2-SH | 6 positive charges, MW 2653 Da |
| Cell penetrating pentapeptide | ||
| CPPP-2 | KLPVM | 2 positive charges, MW 605 Da |
| Histone | ||
| Calf thymus histone, type II-AS | more than 100 amino acids | many positive charges, resulting from Arg and Lys residues (more than 20) |
| Proteins | |
| β-Galactosidase | MW = 540 kDa present study and [24] |
| Bovine serum albumin, ATTO488-labeled | MW = 68 kDa present study and [24] |
| Antibodies | |
| Antibody anti PI3-kinase-γ, monoclonal, unlabeled | MW ≈ 150 kDa not shown |
| Antibody anti actin, monoclonal, ATTO488-labeled | MW ≈ 150 kDa not shown |
| Antibody polyclonal, goat anti mouse, FITC-labeled | MW ≈ 150 kDa [24] |
| Peptides | |
| Backbone cyclic phosphotyrosine octapeptides | MW ≈ 1.1 kDa only functionally characterized [25] |
| Nucleotides | |
| Deoxy nucleoside triphosphate, ATTO-labeled ATTO488-dUTP | MW = 1 kDA |
| 4 negative charges [26] | |
| Adhesion cell lines | |
| HeLa | Human cervix carcinoma |
| COS-7 | African green monkey kidney |
| NIH-3T3 | Swiss mouse embryo |
| Suspension cell lines | |
| Jurkat | Human T cell leukemia |
| NB-4 | Human acute promyelocytic leukemia |
| Kasumi-1 | Human acute myeloid leukemia |
| Protozoa | |
| Leishmania tarentolae | Lizard protozoa |
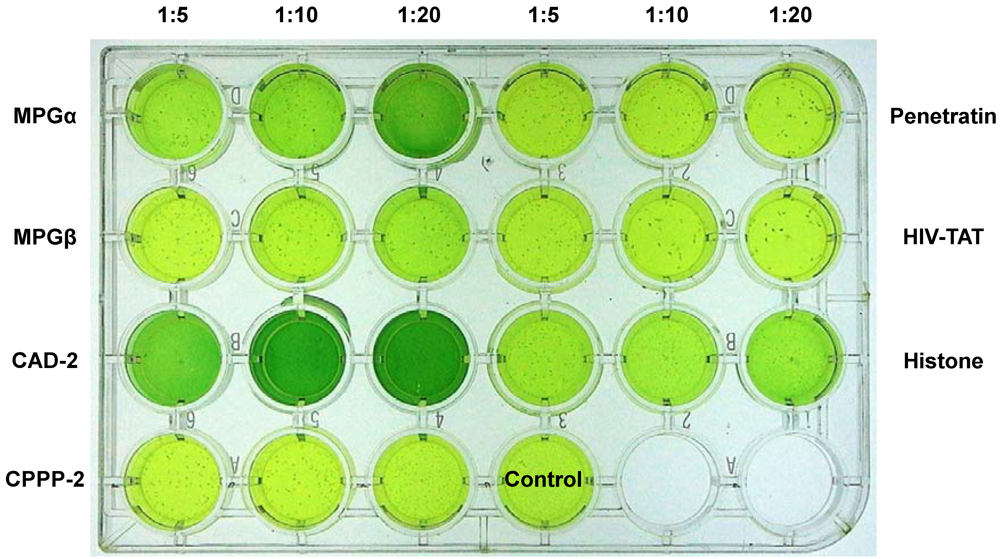
2.2. Proteolytic Activities of Certain Cells and Stabilities of CPPs


2.3. Transport Efficiencies of CPPs
2.3.1.Dependence on Cell Line


2.3.2. Dependency on the Type of Cargo
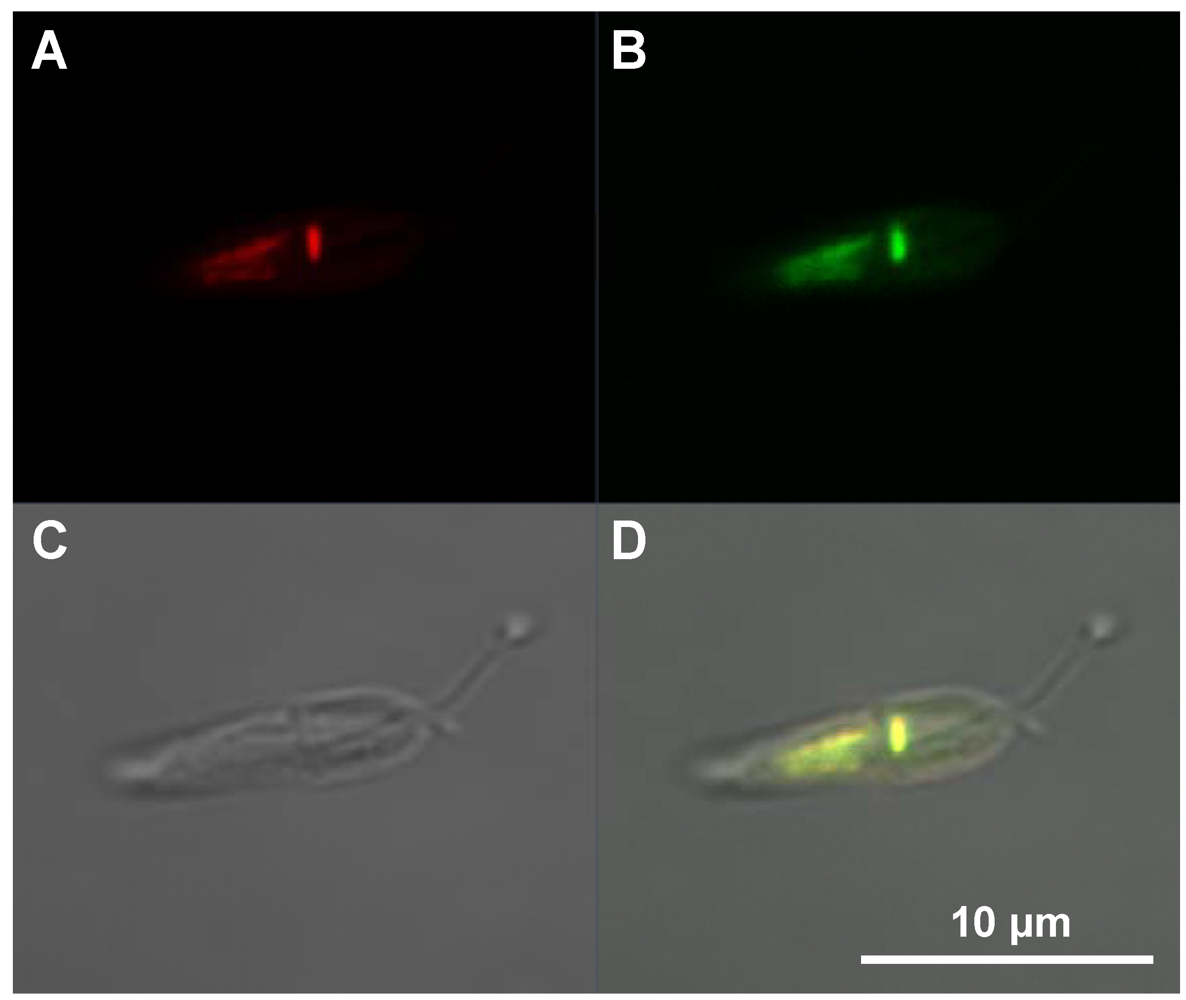
2.4. Intracellular Localization

2.5. Quantification of Transport Efficiencies
| Amount of added complex per 1.6 mL serum-free medium | Internalized amount (0.3 x 106 HeLa cells per well) | |
|---|---|---|
| amol per cell | Intracellular concentration in µM | |
| ATTO488-deoxy uridine triphosphate into HeLa cells [26] | ||
| 1 µg + JBS-Nucleoducin, charge by charge 1:4 | 1.1 | 0.1 |
| 5 µg + JBS-Nucleoducin, charge by charge 1:4 | 2.1 | |
| ATTO488-labeled bovine serum albumin into HeLa cells [24] | ||
| 10 µg + JBS-Proteoducin, molar ratio 1:10 | 20 | |
| 25 µg + JBS-Proteoducin, molar ratio 1:10 | 50 | 4.3 |
| FITC-antibody (secondary) into HeLa cells [24] | ||
| 5 µg + JBS-Proteoducin, molar ratio 1:10 | 0.4 | |
| 10 µg + JBS-Proteoducin, molar ratio 1:10 | 2.2 | |
| 25 µg + JBS-Proteoducin, molar ratio 1:10 | 4.3 | 0.6 |
| ATTO488-labeled bovine serum albumin into Leishmania tarentolae | ||
| 1 µg + MPGα, molar ratio 1:10 | 0.8 x 10−2 | 0.04 |
| 5 µg + MPGα, molar ratio 1:10 | 0.7 x 10−2 | 0.1 |
| 10 µg + MPGα, molar ratio 1:10 | 3.3 x 10−2 | 0.2 |
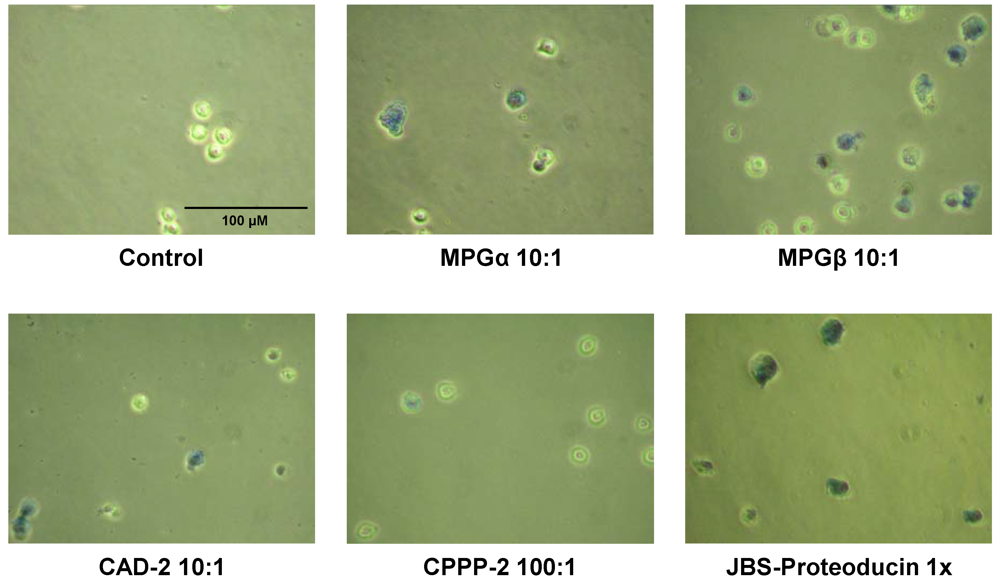
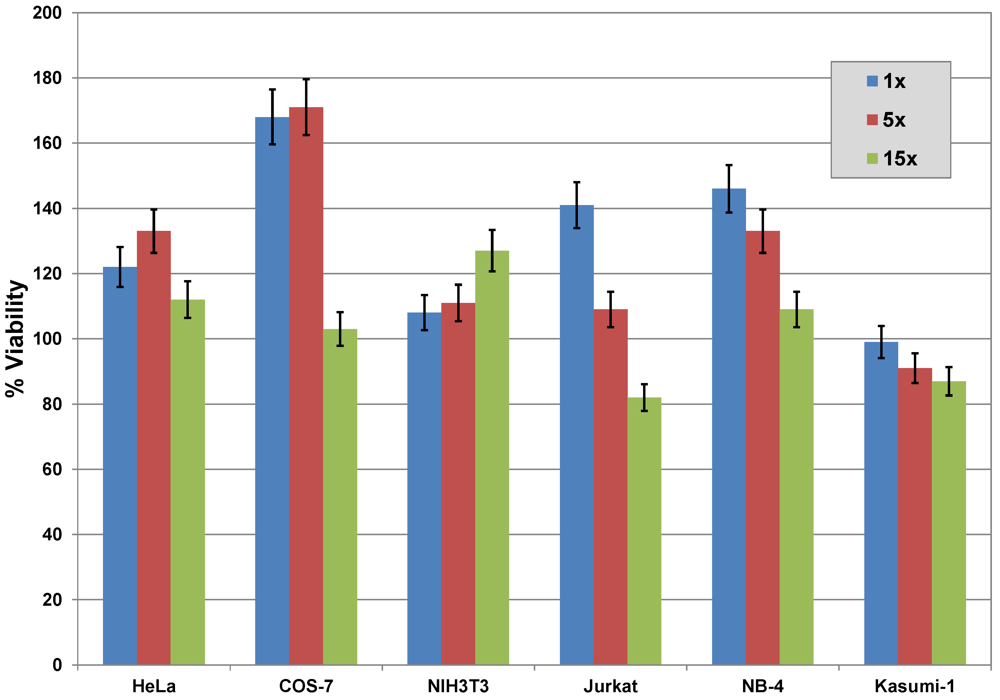
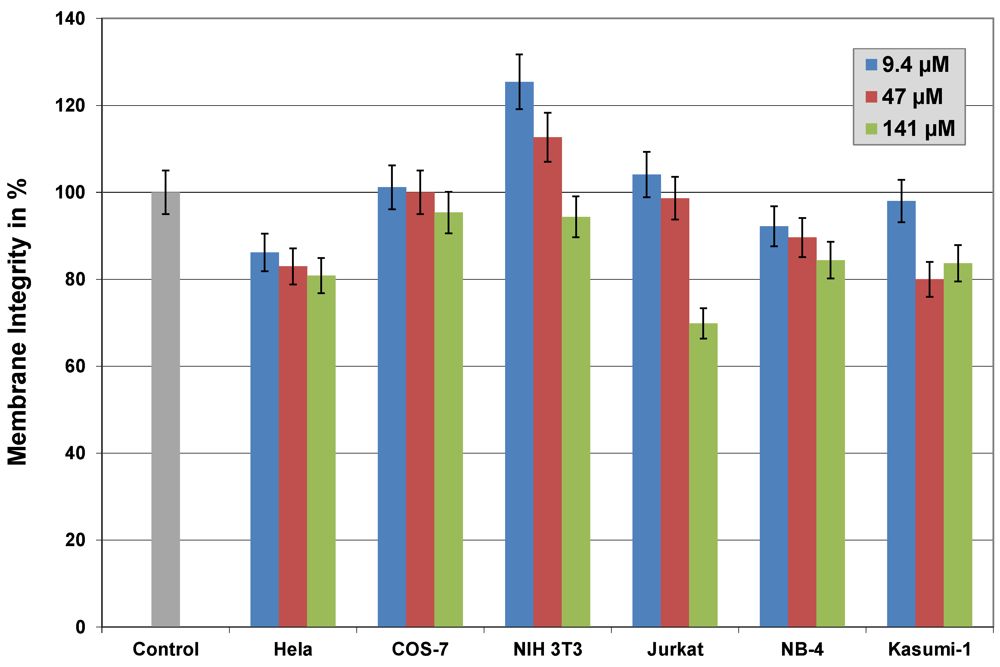
2.6. Cytotoxicity


| Amphiphilic-cationic peptides (transcription factors and -activators) | |
| Penetratin (Antennapedia) | ≤ 10 µM |
| HIV-TAT (47-57) | ≥ 30 µM |
| MPG peptides | |
| MPGα | ≤ 5 µM |
| MPGβ | ≤ 5 µM |
| CAD-2 | ≤ 25 µM |
| Cell penetrating pentapeptide | |
| CPPP-2 | ≥ 100 µM |
| Histone | |
| Calf thymus histone, type II-AS | ≈ 5 µM |
| Cocktails | |
| JBS-Proteoducin | ≥ 50-fold of recommended amount |
| JBS-Nucleoducin | ≥ 30-fold of recommended amount |
3. Experimental
3.1. Materials
3.2. Equipment
3.3. Methods
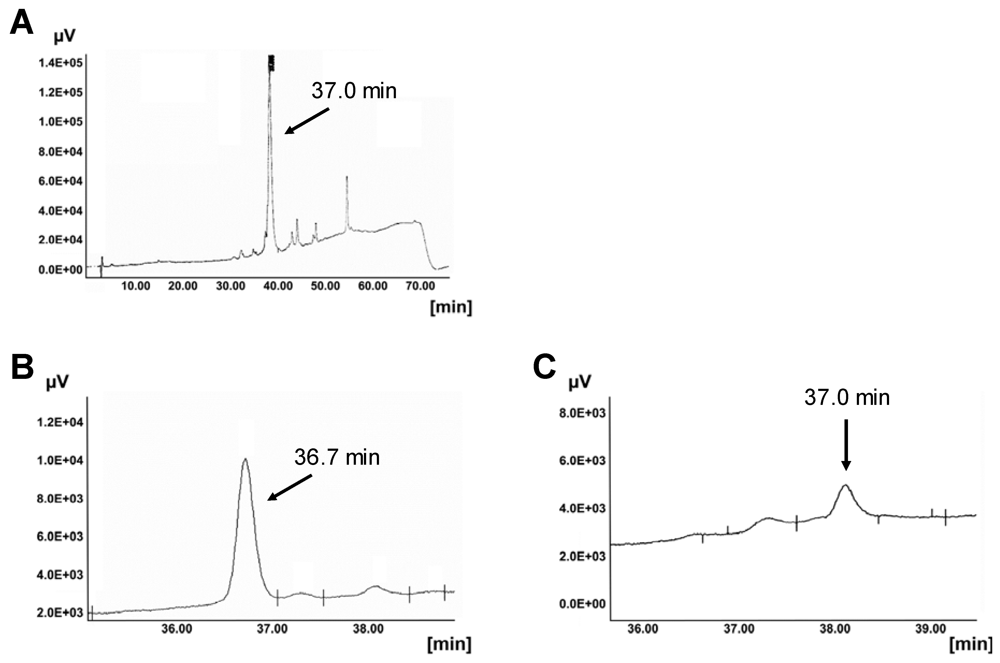
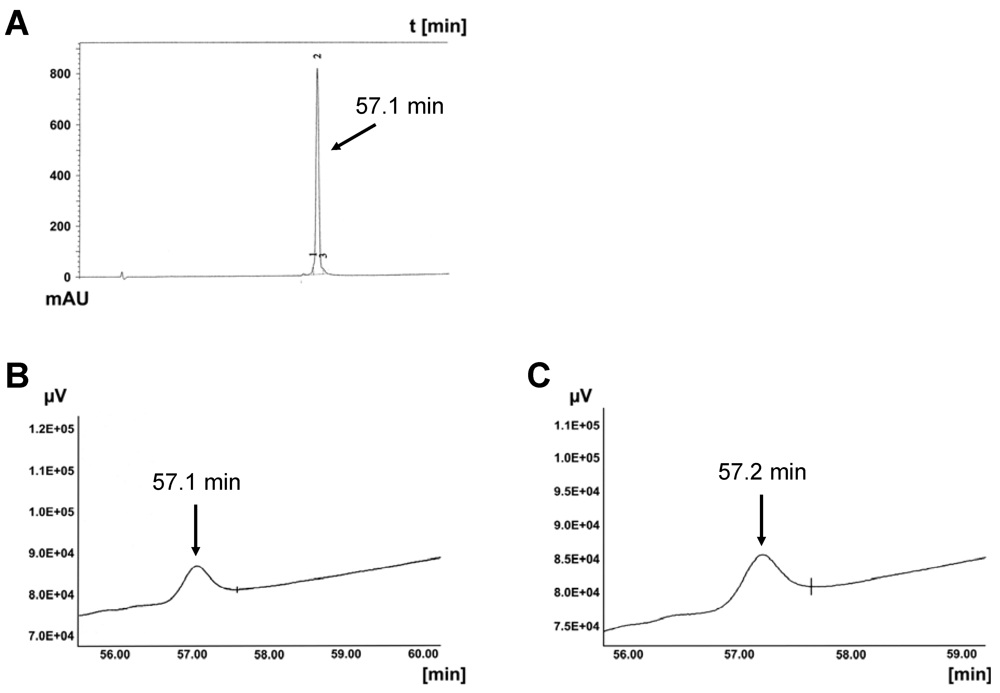
3.3.1. Degradation of CPPs by Proteases
3.3.2. Preparation of Stock Solutions
3.3.3. Complexing Cargoes with Peptides and Proteins
3.3.4. Internalization
3.3.4.1. Internalization of β-Galactosidase and Antibodies into Jurkat Cells
3.3.4.2. Cultivation and Transduction of Leishmania tarentolae
3.3.5. Staining of Internalized β-Galactosidase
3.3.6. Confocal Fluorescence Microscopy
3.3.7. Calculation of Intracellular Cargo Concentrations
3.3.8. Cell Viability Assay
3.3.9. Membrane Integrity Assay
3.3.10. Cytotoxicity Test for Leishmania tarentolae
3.3.11. Mycoplasma Test
4. Conclusions
Acknowledgements
References
- Heitz, F.; Morris, M.C.; Divita, G. Twenty years of cell-penetrating peptides: from molecular mechanism to therapeutics. Br. J. Pharmacol. 2009, 157, 195–206. [Google Scholar]
- Deshayes, S.; Morris, M.; Heitz, F.; Divita, G. Delivery of proteins and nucleic acids using a non-covalent peptide-based strategy. Advanced Drug Del. Reviews 2008, 60, 537–547. [Google Scholar]
- Lindgren, M.; Langel, U. Classes and prediction of cell-penetrating peptides. In Cell penetrating peptides Methods and Protocols. Methods in Molecular Biology; Langel, U., Ed.; Springer: Amsterdam, The Netherlands, 2011; Volume 683, pp. 3–19. [Google Scholar]
- Dupont, E.; Prochiantz, A.; Joliot, A. Penetratin story: An overview. Cell penetrating peptides. In Cell penetrating peptides Methods and Protocols. Methods in Molecular Biology; Langel, U., Ed.; Springer: Amsterdam, The Netherlands, 2011; Volume 683, pp. 21–29. [Google Scholar]
- Deshayes, S.; Plenat, Th.; Aldrian-Herrada, G.; Divita, G.; Le Grimellec, Ch.; Heitz, F. Primary amphipathic cell penetrating peptides: Structural requirements and interactions with model membranes. Biochemistry 2004, 43, 7698–7706. [Google Scholar]
- Kurzawa, L.; Pellerano, M.; Morris, M.C. PEP and CADY-mediated delivery of fluorescent peptides and proteins into living cells. Biochim. Biophys. Acta 2010, 1798, 2274–2285. [Google Scholar]
- Crombez, L.; Aldrian-Herrada, G.; Konate, K.; Nguyen, Qu.N.; McMaster, G.K.; Brasseur, R.; Heitz, F.; Divita, G. A new potent secondary amphipathic cell-penetrating peptide for siRNA delivery into mammalian cells. Mol. Therapy 2009, 17, 95–103. [Google Scholar]
- Gomez, J.A.; Gama, V.; Yoshida, T.; Sun, W.; Hayes, P.; Leskov, K.; Boothman, D.; Matsuyama, S. Bax-inhibiting peptides derived from Ku70 and cell-penetrating pentapeptides. Biochem. Soc. Trans. 2007, 35, 797–801. [Google Scholar]
- Gomez, J.; Matsuyama, S. Cell-penetrating penta-peptides and BAX-inhibiting peptides: Protocol for their application. In Cell penetrating peptides Methods and Protocols. Methods in Molecular Biology; Langel, U., Ed.; Springer: Amsterdam, The Netherland, 2011; Volume 683, pp. 465–471. [Google Scholar]
- Breitling, R.; Klingner, S.; Callewaert, N.; Pietrucha, R.; Geyer, A.; Ehrlich, G.; Hartung, R.; Mueller, A.; Contreras, R.; Beverley, St.M.; Alexandrov, K. Non-pathogenic trypanosomatid protozoa as a platform for protein research and production. Protein Expr. Purific. 2002, 25, 209–218. [Google Scholar]
- Aamand, H.L.; Norden, B.; Fant, K. Functionalization with C-terminal cysteine enhances transfection efficiency of cell-penetrating peptides through dimer formation. Biochem. Biophys. Res. Comm. 2012, 418, 469–474. [Google Scholar]
- Richard, J.P.; Melikov, K.; Brooks, H.; Prevot, P.; Lebleu, B.; Chernomordik, L.V. Cellular uptake of unconjugated TAT peptide involves clathrin-dependent endocytosis and heparan sulfate receptors. J. Biol. Chem. 2005, 280, 15300–15306. [Google Scholar]
- Ignatovich, I.A.; Dishe, E.B.; Pavlotskaya, A.V.; Akifiev, B.N.; Burov, S.V.; Orlov, S.V.; Perevozchikov, A.P. Complexes of plasmid DNA with basic domain 47–57 of HIV-1 TAT protein are transferred to mammalian cells by endocytosis-mediated pathways. J. Biol. Chem. 2003, 43, 42625–42636. [Google Scholar]
- Jeang, K.T.; Xiao, H.; Rich, E.A. Multifaced activities of the HIV-1 transactivator of transcription, TAT. J. Biol. Chem. 1999, 274, 28837–28840. [Google Scholar]
- Gros, E.; Deshayes, S.; Morris, M.C.; Aldrian-Herrada, G.; Depollier, J.; Heitz, F.; Divita, G. A non-covalent peptide-based strategy for protein and peptide nucleic acid transduction. Biochim. Biophys. Acta 2006, 1758, 384–393. [Google Scholar]
- Chaloin, L.; Vidal, P.; Heitz, A.; Van Mau, N.; Mery, J.; Divita, G.; Heitz, F. Conformations of primary amphipathic carrier peptides in membrane mimicking environments. Biochemistry 1997, 36, 11179–11187. [Google Scholar] [CrossRef]
- Deshayes, S.; Heitz, A.; Morris, M.C.; Charnet, P.; Divita, G.; Heitz, F. Insight into mechanism of internalization of cell-penetrating carrier peptide Pep-1 through conformational analysis. Biochemistry 2004, 43, 1449–1457. [Google Scholar]
- Morris, M.C.; Gros, E.; Aldrian-Herrada, G.; Choob, M.; Archdeacon, J.; Heitz, F.; Divita, G. A non-covalent peptide-based carrier for in vivo delivery of DNA mimics. Nucleic Acid Res. 2007, 35 No7, e49. [Google Scholar]
- Mäe, M.; EL Andaloussi, S.; Lundin, P.; Oskolkov, N.; Johansson, H.J.; Guterstam, P.; Langel, U. A stearylated CPP for delivery of splice correcting oligonucleotides using a non-covalent co-incubation strategy. J. Controlled Rel. 2009, 134, 221–227. [Google Scholar]
- Oskolkov, N.; Arukuusk, P.; Copolovici, D.-M.; Lindberg, St.; Margus, H.; Padar, K.; Pooga, M.; Langel, U. NickFects, phosphorylated derivatives of transportan 10 for cellular delivery of oligonucleotides. Int. J. Pept. Res. Therapeutics 2011, 17, 147–157. [Google Scholar] [CrossRef]
- Ezzat, K.; EL Andaloussi, S.; Zaghloul, E.M.; Lehto, T.; Lindberg, St.; Moreno, P.M.D.; Viola, J.R.; Magdy, T.; Abdo, R.; Guterstam, P.; Sillard, R.; Hammond, S.M.; Wood, M.J.A.; Arzumanov, A.A.; Gait, M.J.; Smith, C.I.E.; Hällbrink, M.; Langel, U. PepFect 14, a novel cell-penetrating peptide for oligonucleotide delivery in solution and as solid formulation. Nucleic Acid Res. 2011, 39, 5284–5298. [Google Scholar]
- Hariton-Gazal, E.; Rosenbluh, J.; Graessmann, A.; Gilon, C.; Loyter, A. Direct translocation of histone molecules across cell membranes. J. Cell Sci. 2003, 116, 4577–4586. [Google Scholar] [CrossRef]
- Singh, R.K.; Liang, D.; Gajjalaiavari, U.R.; Kabaj, M.-H.M.; Paik, J.; Gunjan, A. Excess histone levels mediate cytotoxicity via multiple mechanisms. Cell Cycle 2010, 9, 4236–4244. [Google Scholar] [CrossRef]
- Mussbach, F.; Franke, M.; Zoch, A.; Schaefer, B.; Reissmann, S. Transduction of peptides and proteins into live cells by cell penetrating peptides. J. Cell. Biochem. 2011, 112, 3824–3833. [Google Scholar] [CrossRef]
- Zoda, M.S.; Zacharias, M.; Mussbach, F.; Schaefer, B.; Reissmann, S. Assembly and stimulatory activity of backbone to side chain cyclic octapeptide-ligands for the N-terminal SH2-domain of the protein-tyrosine phosphatase SHP-1. In Proceedings of the 31st European Peptide Symposium September 5-9, 2010, Copenhagen, Denmark; pp. 210–211.
- Mussbach, F.; Pietrucha, R.; Schaefer, B.; Reissmann, S. Internalization of nucleoside phosphates into live cells by complex formation with different cell penetrating peptides and JBS-Nucleoducin. In Cell penetrating peptides Methods and Protocols. Methods in Molecular Biology; Langel, U., Ed.; Springer: Amsterdam, The Netherland, 2011; Volume 683, pp. 33–40. [Google Scholar]
- LEXSY-Eukaryotic protein expression in Leishmania tarentolae. Available online: http://www.jenabioscience.com/cms/en/1/browse/1838, accessed on 4 February 2013.
- Keller, A.-A.; Breitling, R.; Hemmerich, P.; Braun, M.; Schaefer, B.; Lorkowski, S.; Reissmann, S. Transduction of proteins into Leishmania tarentolae by formation of non-covalent complexes with cell-penetrating peptides. In preparation.
- Wu, R.P.; Youngblood, D.S.; Hassinger, J.N.; Lovejoy, C.E.; Nelson, M.H.; Iversen, P.L.; Moulton, H.M. Cell-penetrating peptides as transporters for morpholino oligomers: effects of amino acid composition on intracellular delivery and cytotoxicity. Nucleic Acids Res. 2007, 35, 5182–5191. [Google Scholar] [CrossRef]
- Niles, A.L.; Moravec, R.A.; Hesselberth, P.E.; Scurria, M.A.; Daily, W.J.; Riss, T.L. A homogeneous assay to measure live and dead cells in the same sample by detecting different protease markers. Anal. Biochem. 2007, 366, 197–206. [Google Scholar] [CrossRef]
- Brand, P.; Lenser, T.; Hemmerich, P. Assembly dynamics of PML nuclear bodies in living cells. PMC Biophysics 2010, 3. [Google Scholar] [CrossRef]
- Deshayes, S.; Konate, K.; Aldrian, G.; Crombez, L.; Heitz, F.; Divita, G. Structural polymorphism of non-covalent peptide-based delivery systems: Highway to cellular uptake. Biochim. Biophys. Acta 2010, 1798, 2304–2314. [Google Scholar] [CrossRef]
- Nakase, I.; Tadokoro, A.; Kawabata, N.; Takeuchi, T.; Katoh, H.; Hiramoto, K.; Negishi, M.; Nomizu, M.; Sugiura, Y.; Futaki, S. Interaction of arginine-rich peptides with membrane-associated proteoglycans is crucial for induction of actin organization and macropinocytosis. Biochemistry 2007, 46, 492–501. [Google Scholar]
- Foerg, C.; Ziegler, U.; Fernandez-Carneado, J.; Giralt, E.; Renner, R.; Beck-Sickinger, A.; Merkle, H.P. Decoding the entry of two novel cell-penetrating peptides in HeLa-cells: Lipid raft-mediated endocytosis and endosomal escape. Biochemistry 2005, 44, 72–81. [Google Scholar]
- Madani, F.; Lindberg, S.; Langel, U.; Futaki, S.; Graeslund, A. Mechanism of cellular uptake of cell-penetrating peptides. J. Biophysics 2011, 1–10. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Keller, A.-A.; Mussbach, F.; Breitling, R.; Hemmerich, P.; Schaefer, B.; Lorkowski, S.; Reissmann, S. Relationships between Cargo, Cell Penetrating Peptides and Cell Type for Uptake of Non-Covalent Complexes into Live Cells. Pharmaceuticals 2013, 6, 184-203. https://doi.org/10.3390/ph6020184
Keller A-A, Mussbach F, Breitling R, Hemmerich P, Schaefer B, Lorkowski S, Reissmann S. Relationships between Cargo, Cell Penetrating Peptides and Cell Type for Uptake of Non-Covalent Complexes into Live Cells. Pharmaceuticals. 2013; 6(2):184-203. https://doi.org/10.3390/ph6020184
Chicago/Turabian StyleKeller, Andrea-Anneliese, Franziska Mussbach, Reinhard Breitling, Peter Hemmerich, Buerk Schaefer, Stefan Lorkowski, and Siegmund Reissmann. 2013. "Relationships between Cargo, Cell Penetrating Peptides and Cell Type for Uptake of Non-Covalent Complexes into Live Cells" Pharmaceuticals 6, no. 2: 184-203. https://doi.org/10.3390/ph6020184
APA StyleKeller, A. -A., Mussbach, F., Breitling, R., Hemmerich, P., Schaefer, B., Lorkowski, S., & Reissmann, S. (2013). Relationships between Cargo, Cell Penetrating Peptides and Cell Type for Uptake of Non-Covalent Complexes into Live Cells. Pharmaceuticals, 6(2), 184-203. https://doi.org/10.3390/ph6020184