
Tryptophan Breakdown in Patients with HCV Infection is Influenced by IL28B Polymorphism


Abstract
:1. Introduction
2. Experimental Section
2.1. Patients

{kind=link}
{kind=link}
| Age (y) | Sex | Fibrosis status | IL28B genotype | HCV genotype | HCV LOG10 | Therapy Response * |
|---|---|---|---|---|---|---|
| 23.5 | Female | No cirrhosis | T/T | 3a | 5.36 | SVR |
| 24.5 | Female | No cirrhosis | C/T | 3a | 5.67 | SVR |
| 25.2 | Female | F0-1 | T/T | 1a | 5.50 | LFU |
| 30.1 | Male | F0-1 | T/T | 3a | 5.34 | SVR |
| 30.4 | Female | F0-1 | C/T | 1a | 5.75 | SVR |
| 30.6 | Female | F0-1 | C/C | 1b | 7.02 | Relapse |
| 31.1 | Male | F0-1 | C/C | 3a | 4.64 | SVR |
| 32.5 | Female | No cirrhosis | C/T | 3a | 5.82 | LFU |
| 34.0 | Female | F0-1 | C/T | 4 | 3.90 | SVR |
| 38.9 | Male | F2 | T/T | 4 | 5.05 | SVR |
| 39.8 | Male | F0-1 | T/T | 1a | 6.91 | No response |
| 39.9 | Female | F0-1 | C/T | 3a | 5.93 | SVR |
| 40.7 | Male | F3 | C/T | 3a | 4.72 | Relapse |
| 42.3 | Male | No cirrhosis | C/T | 3a | 5.62 | SVR |
| 45.1 | Female | No chirrhosis | C/C | 3a | 3.43 | SVR |
| 45.3 | Female | F3 | C/T | 3a | 6.53 | SVR |
| 48.0 | Male | Chirrhosis Child a | C/C | 1b | 5.36 | SVR |
| 48.2 | Male | F0-1 | C/T | 1a | 7.52 | Relapse |
| 49.1 | Male | F3 | C/T | 3a | 6.34 | Relapse |
| 54.0 | Female | F0-1 | C/C | 1a | 7.01 | Relapse |
| 54.0 | Male | F3 | C/C | 3a | 6.61 | Relapse |
| 54.8 | Male | Chirrhosis Child A | C/T | 3 | 6.48 | Relapse |
| 55.0 | Female | F3 | C/T | 1b | 6.16 | SVR |
| 60.0 | Male | Chirrhosis Child A | C/C | 2 | 6.30 | SVR |
| 70.1 | Female | F2 | C/C | 1b | 4.13 | LFU |
2.2. Laboratory Variables
2.3. Statistical Analysis
3. Results


4. Discussion
| IL28B Polymorphism | C/C | C/T | T/T | non-C/C | Χ2, p * | U, p ** |
|---|---|---|---|---|---|---|
| (n = 8) | (n = 5) | (n = 12) | (n = 17) | |||
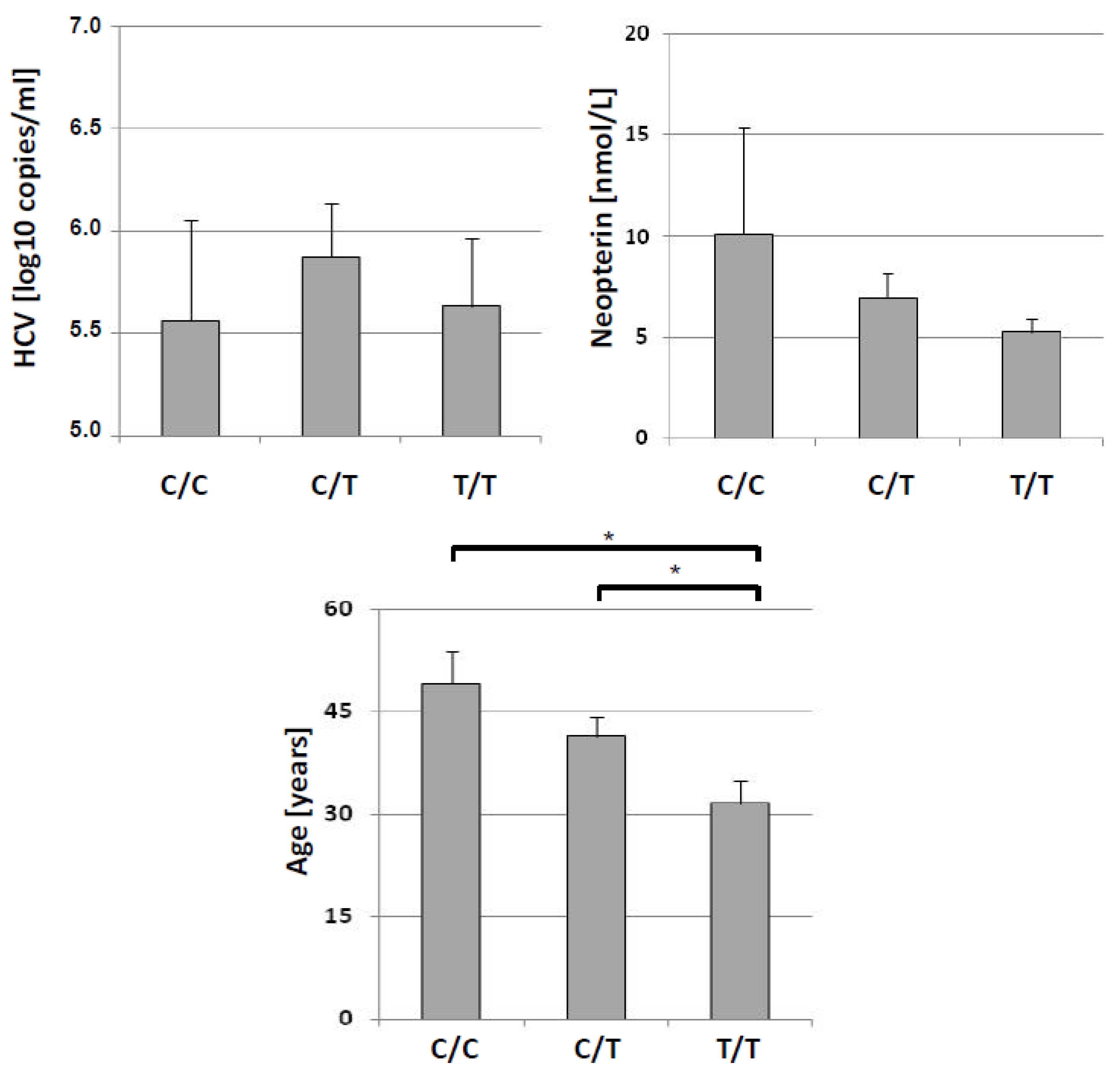
| HCV load [log 10 copies/µL] | 5.56 ± 1.38 | 5.87 ± 0.92 | 5.63 ± 0.73 | 5.80 ± 0.85 | 0.183, n.s. | 0.146, n.s. |
| Age [y] | 49.1 ± 13.6 | 41.4 ± 9.67 | 31.5 ± 7.57 | 38.5 ± 10.1 | 7.044, 0.030 | 1.806, 0.075 |
| Tryptophan [µmol/L] | 49.5 ± 10.5 | 55.8 ± 17.7 | 43.8 ± 9.54 | 52.2 ± 16.4 | 2.603, n.s. | 0.058, n.s. |
| Kynurenine [µmol/L] | 2.76 ± 1.48 | 1.92 ± 0.55 | 1.41 ± 0.44 | 1.77 ± 0.55 | 5.627, 0.060 | 1.748, 0.086 |
| Kyn/Trp [µmol/mmol] | 56.4 ± 33.7 | 35.7 ± 10.9 | 31.9 ± 4.52 | 33.7 ± 9.50 | 6.218, <0.05 | 2.447, 0.014 |
| Nitrite [µmol/L] | 6.58 ± 5.16 | 10.6 ± 16.7 | 14.32 ± 15.3 | 8.17 ± 10.1 | 0.916, n.s. | 0.233, n.s. |
| Neopterin [nmol/L] | 10.1 ± 14.8 | 6.96 ± 4.30 | 5.28 ± 1.53 | 6.47 ± 3.73 | 0.805, n.s. | 0.175, n.s. |
Limitations
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Manns, M.P.; McHutchison, J.G.; Gordon, S.C.; Rustgi, V.K.; Shiffman, M.; Reindollar, R.; Goodman, Z.D.; Koury, K.; Ling, M.; Albrecht, J.K. Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis C, a randomised trial. Lancet 2001, 358, 958–965. [Google Scholar] [PubMed]
- Witthöft, T.; Möller, B.; Wiedmann, K.H.; Mauss, S.; Link, R.; Lohmeyer, J.; Lafrenz, M.; Gelbmann, C.M.; Hüppe, D.; Niederau, C.; et al. Safety, tolerability and efficacy of peginterferon alpha-2a and ribavirin in chronic hepatitis C in clinical practice, The German Open Safety Trial. J. Viral Hepat. 2007, 14, 788–796. [Google Scholar] [PubMed]
- Ge, D.; Fellay, J.; Thompson, A.J.; Gumbs, C.E.; Urban, T.J.; Shianna, K.V.; Little, L.D.; Qiu, P.; Bertelsen, A.H.; Watson, M.; Warner, A.; et al. Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance. Nature 2009, 461, 399–401. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Nishida, N.; Sugiyama, M.; Kurosaki, M.; Matsuura, K.; Sakamoto, N.; Nakagawa, M.; Korenaga, M.; Hino, K.; Hige, S.; et al. Genome-wide association of IL28B with response to pegylated interferon-alpha and ribavirin therapy for chronic hepatitis C. Nat. Genet. 2009, 41, 1105–1109. [Google Scholar]
- Lotrich, F.E.; Loftis, J.M.; Ferrell, R.E.; Rabinovitz, M.; Hauser, P. IL28B polymorphism is associated with both side effects and clearance of hepatitis C during interferon-α therapy. J. Interferon Cytokine Res. 2011, 31, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Chen, J.Y.; Lin, T.N.; Jeng, W.J.; Huang, C.H.; Huang, C.W.; Chang, S.W.; Sheen, I.S. IL28B SNP rs12979860 is a critical predictor for on-treatment and sustained virologic response in patients with hepatitis C virus genotype-1 infection. PLoS ONE 2011, 6, e18322. [Google Scholar] [CrossRef] [PubMed]
- Rauch, A.; Kutalik, Z.; Descombes, P.; Cai, T.; Di Iulio, J.; Mueller, T.; Bochud, M.; Battegay, M.; Bernasconi, E.; Borovicka, J.; et al.; Swiss Hepatitis C Cohort Study, Swiss HIV Cohort Study Genetic variation in IL28B is associated with chronic hepatitis C and treatment failure, a genome-wide association study. Gastroenterology 2010, 138, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.L.; Thio, C.L.; Martin, M.P.; Qi, Y.; Ge, D.; O'Huigin, C.; Kidd, J.; Kidd, K.; Khakoo, S.I.; Alexander, G.; et al. Genetic variation in IL28B and spontaneous clearance of hepatitis C virus. Nature 2009, 461, 798–801. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.M.; Zeuzem, S. IL28B single nucleotide polymorphisms in the treatment of hepatitis C. J. Hepatol. 2011, 55, 692–701. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Derrick, J.; Hong, J.; Sanda, C.; Grosse, W.M.; Edenberg, H.J.; Taylor, M.; Seiwert, S.; Blatt, L.M. Global transcriptional profiling demonstrates the combination of type I and type II interferon enhances antiviral and immune responses at clinically relevant doses. J. Interferon Cytokine Res. 2005, 25, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Byrne, G.I.; Lehmann, L.K.; Kirschbaum, J.G.; Borden, E.C.; Lee, C.M.; Brown, R.R. Induction of tryptophan degradation in vitro and in vivo, a gamma-interferon-stimulated activity. J. Interferon Res. 1986, 6, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Werner, E.R.; Bitterlich, G.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Szabo, G.; Dierich, M.P.; Wachter, H. Human macrophages degrade tryptophan upon induction by interferon-gamma. Life Sci. 1987, 41, 273–280. [Google Scholar] [CrossRef]
- Werner-Felmayer, G.; Werner, E.R.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Wachter, H. Characteristics of interferon induced tryptophan metabolism in human cells in vitro. Biochim. Biophys. Acta 1989, 1012, 140–147. [Google Scholar] [CrossRef]
- Wirleitner, B.; Reider, D.; Ebner, S.; Böck, G.; Widner, B.; Jaeger, M.; Schennach, H.; Romani, N.; Fuchs, D. Monocyte-derived dendritic cells release neopterin. J. Leukocyte Biol. 2002, 72, 1148–1153. [Google Scholar] [PubMed]
- Taylor, M.W.; Feng, G.S. Relationship between interferon-gamma, indoleamine 2,3-dioxygenase, and tryptophan catabolism. FASEB J. 1991, 5, 2516–2522. [Google Scholar] [PubMed]
- Pfefferkorn, E.R. Interferon gamma blocks the growth of Toxoplasma gondii in human fibroblasts by inducing the host cells to degrade tryptophan. Proc. Natl. Acad. Sci. USA 1984, 81, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Adams, O.; Besken, K.; Oberdörfer, C.; MacKenzie, C.R.; Takikawa, O.; Däubener, W. Role of indoleamine-2,3-dioxygenase in alpha/beta and gamma interferon-mediated antiviral effects against herpes simplex virus infections. J. Virol. 2004, 78, 2632–2636. [Google Scholar] [CrossRef] [PubMed]
- Larrea, E.; Riezu-Boj, J.I.; Gil-Guerrero, L.; Casares, N.; Aldabe, R.; Sarobe, P.; Civeira, M.P.; Heeney, J.L.; Rollier, C.; Verstrepen, B.; et al. Upregulation of indoleamine 2,3-dioxygenase in hepatitis C virus infection. J. Virol. 2007, 81, 3662–3666. [Google Scholar] [CrossRef] [PubMed]
- Higashitani, K.; Kanto, T.; Kuroda, S.; Yoshio, S.; Matsubara, T.; Kakita, N.; Oze, T.; Miyazaki, M.; Sakakibara, M.; Hiramatsu, N.; et al. Association of enhanced activity of indoleamine 2,3-dioxygenase in dendritic cells with the induction of regulatory T cells in chronic hepatitis C infection. J. Gastroenterol. 2013, 48, 660–670. [Google Scholar] [CrossRef] [PubMed]
- Hevel, J.M.; White, K.A.; Marletta, M.A. Identification of the inducible murine macrophage nitric oxide synthase. Identification as a flavoprotein. J. Biol. Chem. 1991, 266, 22789–22791. [Google Scholar] [PubMed]
- Werner, E.R.; Werner-Felmayer, G.; Fuchs, D.; Hausen, A.; Reibnegger, G.; Schmidt, K.; Weiss, G.; Wachter, H. Tetrahydrobiopterin biosynthetic activities in human macrophages.; fibroblasts.; THP-1 and T 24 cells. GTP-cyclohydrolase I is stimulated by interferon-gamma, 6-pyruvoyl tetrahydropterin synthase and sepiapterin reductase are constitutively present. J. Biol. Chem. 1990, 265, 3189–3192. [Google Scholar] [PubMed]
- Murr, C.; Widner, B.; Wirleitner, B.; Fuchs, D. Neopterin as a marker for immune system activation. Curr. Drug Metabol. 2002, 3, 175–187. [Google Scholar] [CrossRef]
- Widner, B.; Werner, E.R.; Schennach, H.; Wachter, H.; Fuchs, D. Simultaneous measurement of serum tryptophan and kynurenine by HPLC. Clin. Chem. 1997, 43, 2424–2426. [Google Scholar] [PubMed]
- Fuchs, D.; Möller, A.A.; Reibnegger, G.; Stöckle, E.; Werner, E.R.; Wachter, H. Decreased serum tryptophan in patients with HIV-1 infection correlates with increased serum neopterin and with neurologic/psychiatric symptoms. J. Acquir. Immune Defic. Syndr. 1990, 3, 873–876. [Google Scholar] [PubMed]
- Mayersbach, P.; Augustin, R.; Schennach, H.; Schönitzer, D.; Werner, E.R.; Wachter, H.; Reibnegger, G. Commercial enzyme-linked immunosorbent assay for neopterin detection in blood donations compared with RIA and HPLC. Clin. Chem. 1994, 40, 265–266. [Google Scholar] [PubMed]
- Giustarini, D.; Dalle-Donne, I.; Colombo, R.; Milzani, A.; Rossi, R. Adaptation of the Griess reaction for detection of nitrite in human plasma. Free Radic. Res. 2004, 38, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Geisler, S.; Mayersbach, P.; Becker, K.; Schennach, H.; Fuchs, D.; Gostner, J.M. Serum tryptophan, kynurenine, phenylalanine, tyrosine and neopterin concentrations in 100 healthy blood donors. Pteridines 2015, 26, 31–36. [Google Scholar] [CrossRef]
- Muir, A.J. IL28B in the era of direct-acting antivirals for hepatitis C. J. Clin. Gastroenterol. 2013, 47, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Munn, D.H.; Mellor, A.L. Indoleamine 2,3 dioxygenase and metabolic control of immune responses. Trends Immunol. 2013, 34, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guillemin, G.J. Kynurenine pathway metabolites in humans, disease and healthy states. Int. J. Tryptophan Res. 2009, 2, 1–19. [Google Scholar] [PubMed]
- Widner, B.; Laich, A.; Sperner-Unterweger, B.; Ledochowski, M.; Fuchs, D. Neopterin production, tryptophan degradation, and mental depression—What is the link? Brain Behav. Immun. 2002, 16, 590–595. [Google Scholar] [CrossRef]
- Raison, C.L.; Broadwell, S.D.; Borisov, A.S.; Manatunga, A.K.; Capuron, L.; Woolwine, B.J.; Jacobson, I.M.; Nemeroff, C.B.; Miller, A.H. Depressive symptoms and viral clearance in patients receiving interferon-alpha and ribavirin for hepatitis C. Brain Behav. Immun. 2005, 19, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Zignego, A.L.; Cozzi, A.; Carpenedo, R.; Giannini, C.; Rosselli, M.; Biagioli, T.; Aldinucci, A.; Laffi, G.; Moroni, F. HCV patients, psychopathology and tryptophan metabolism, analysis of the effects of pegylated interferon plus ribavirin treatment. Dig. Liver Dis. 2007, 39, S107–S111. [Google Scholar] [CrossRef]
- Schroecksnadel, K.; Wirleitner, B.; Winkler, C.; Fuchs, D. Monitoring tryptophan metabolism in chronic immune activation. Clin. Chim. Acta 2006, 364, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, D.; Norkrans, G.; Wejstal, R.; Reibnegger, G.; Weiss, G.; Weiland, O.; Schvarcz, R.; Fryden, A.; Wachter, H. Changes of serum neopterin, β2-microglobulin and interferon-gamma in patients with chronic hepatitis C treated with interferon-alpha 2b. Eur. J. Med. 1992, 1, 196–200. [Google Scholar] [PubMed]
- Beinhardt, S.; Aberle, J.H.; Strasser, M.; Dulic-Lakovic, E.; Maieron, A.; Kreil, A.; Rutter, K.; Staettermayer, A.F.; Datz, C.; Scherzer, T.M.; et al. Serum level of IP-10 increases predictive value of IL28B polymorphisms for spontaneous clearance of acute HCV infection. Gastroenterology 2012, 142, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Payer, B.A.; Reiberger, T.; Aberle, J.; Ferenci, P.; Holzmann, H.; Rieger, A.; Peck-Radosavljevic, M.; Vienna HIV-HCV study group. IL28B and interferon-gamma inducible protein 10 for prediction of rapid virologic response and sustained virologic response in HIV-HCV-coinfected patients. Eur. J. Clin. Invest. 2012, 42, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.S.; Grisham, M.B. Methods to detect nitric oxide and its metabolites in biological samples. Free Radic. Biol. Med. 2007, 43, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.R.; Mohr, D.; Stocker, R. Nitric oxide inhibits indoleamine 2,3-dioxygenase activity in interferon-gamma primed mononuclear phagocytes. J. Biol. Chem. 1994, 269, 14457–14464. [Google Scholar] [PubMed]
- Fattovich, G.; Covolo, L.; Bibert, S.; Askarieh, G.; Lagging, M.; Clément, S.; Malerba, G.; Pasino, M.; Guido, M.; Puoti, M.; et al.; ITAHEC Study Group IL28B polymorphisms, IP-10 and viral load predict virological response to therapy in chronic hepatitis C. Aliment. Pharmacol. Ther. 2011, 33, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Dill, M.T.; Duong, F.H.; Vogt, J.E.; Bibert, S.; Bochud, P.Y.; Terracciano, L.; Papassotiropoulos, A.; Roth, V.; Heim, M.H. Interferon-induced gene expression is a stronger predictor of treatment response than IL28B genotype in patients with hepatitis C. Gastroenterology. 2011, 140, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Oxenkrug, G.F.; Requintina, P.J.; Mikolich, D.L.; Ruthazer, R.; Viveiros, K.; Lee, H.; Summergrad, P. Neopterin as a marker of response to antiviral therapy in hepatitis C virus patients. Hepat. Res. Treat. 2012, 2012, 619609. [Google Scholar] [CrossRef] [PubMed]
- Lagging, M.; Askarieh, G.; Negro, F.; Bibert, S.; Söderholm, J.; Westin, J.; Lindh, M.; Romero, A.; Missale, G.; Ferrari, C.; et al.; DITTO-HCV Study Group Response prediction in chronic hepatitis C by assessment of IP-10 and IL28B-related single nucleotide polymorphisms. PLoS ONE 2011, 6, e17232. [Google Scholar] [CrossRef] [PubMed]
- Grebely, J.; Page, K.; Sacks-Davis, R.; van der Loeff, M.S.; Rice, T.M.; Bruneau, J.; Morris, M.D.; Hajarizadeh, B.; Amin, J.; Cox, A.L.; et al.; InC3 Study Group The effects of female sex, viral genotype, and IL28B genotype on spontaneous clearance of acute hepatitis C virus infection. Hepatology 2014, 59, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Frick, B.; Schroecksnadel, K.; Neurauter, G.; Leblhuber, F.; Fuchs, D. Increasing production of homocysteine and neopterin and degradation of tryptophan with older age. Clin. Biochem. 2004, 37, 684–687. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoller, H.; Jenal, A.; Staettermayer, A.F.; Schroecksnadel, S.; Ferenci, P.; Fuchs, D. Tryptophan Breakdown in Patients with HCV Infection is Influenced by IL28B Polymorphism. Pharmaceuticals 2015, 8, 337-350. https://doi.org/10.3390/ph8020337
Zoller H, Jenal A, Staettermayer AF, Schroecksnadel S, Ferenci P, Fuchs D. Tryptophan Breakdown in Patients with HCV Infection is Influenced by IL28B Polymorphism. Pharmaceuticals. 2015; 8(2):337-350. https://doi.org/10.3390/ph8020337
Chicago/Turabian StyleZoller, Heinz, Annina Jenal, Albert F. Staettermayer, Sebastian Schroecksnadel, Peter Ferenci, and Dietmar Fuchs. 2015. "Tryptophan Breakdown in Patients with HCV Infection is Influenced by IL28B Polymorphism" Pharmaceuticals 8, no. 2: 337-350. https://doi.org/10.3390/ph8020337
APA StyleZoller, H., Jenal, A., Staettermayer, A. F., Schroecksnadel, S., Ferenci, P., & Fuchs, D. (2015). Tryptophan Breakdown in Patients with HCV Infection is Influenced by IL28B Polymorphism. Pharmaceuticals, 8(2), 337-350. https://doi.org/10.3390/ph8020337