
The Role of Immunotherapy in Multiple Myeloma

Abstract
:1. Introduction
2. Immune Dysregulation in Multiple Myeloma
3. Immunotherapy in Multiple Myeloma
- (1)
- Reverse tumor mediated immune paralysis
- Immunomodulatory drugs (e.g., IMiDs).
- By blocking inhibitory molecules or, alternatively, activating stimulatory molecules, these treatments are intended to enhance pre-existing anti-myeloma immune responses. (e.g., immune checkpoint inhibitors).
- Cytokines (e.g., interferon and GM-CSF, Siltuximab, an anti-IL-6 antibody and ALT-803, an interleukin 15 agonist).
- (2)
- Stimulate myeloma specific immune responses
- MM Vaccines (e.g., dendritic cell based, peptide based antibodies).
- Adoptive T Cell Transfer (focused on genetically modified T cells to target various MM antigens. (e.g., chimeric antigen receptor (CAR) T cells using BCMA and CD-19 or TCR cells engineered to target the NY-ESO-1 antigen).
- (3)
- Selectively eliminate the malignant clone.
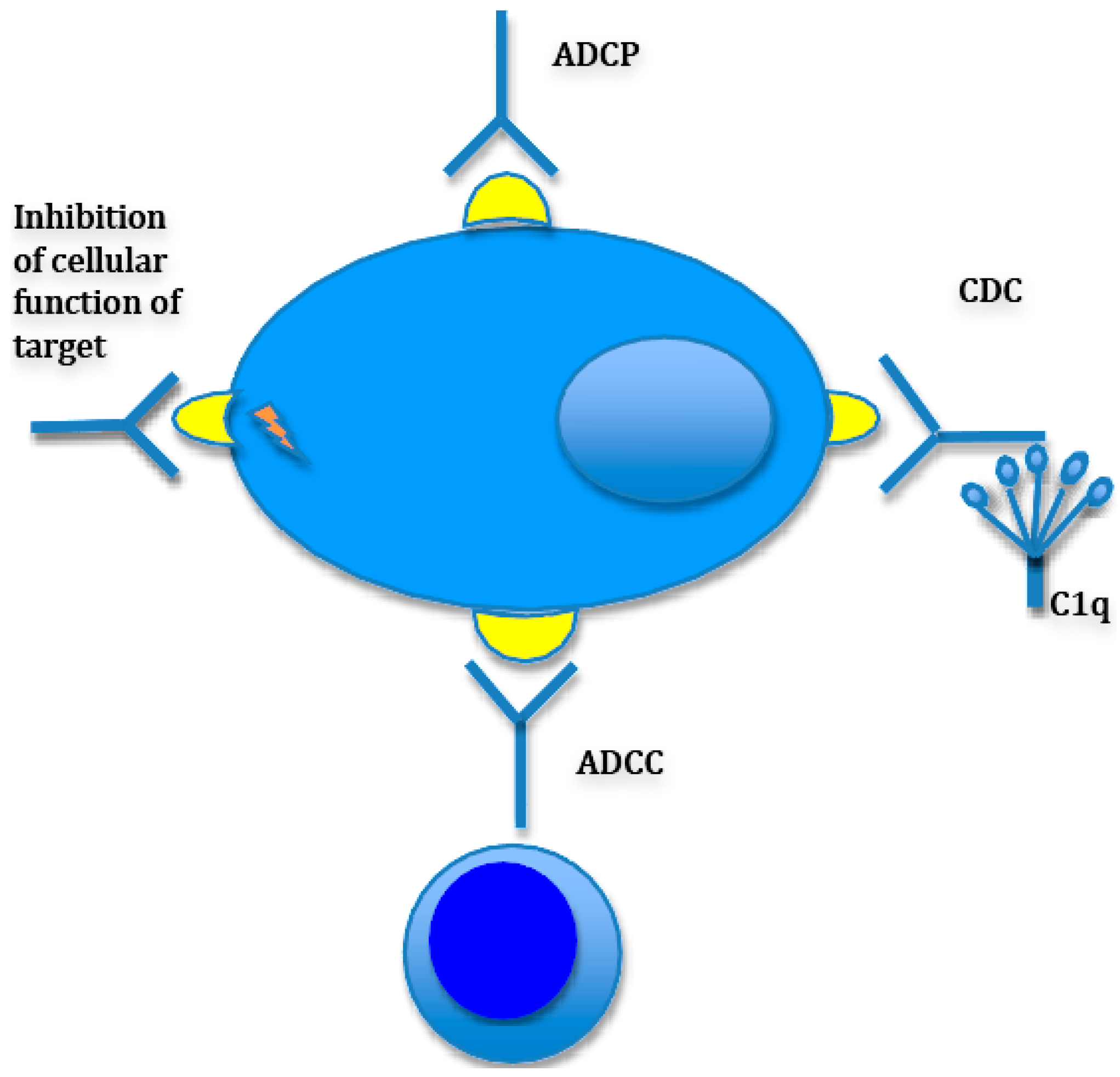
- Monoclonal antibodies (e.g., daratumumab, an antibody against CD38 and elotuzumab, an antibody targeting SLAMF7).
3.1. Immunomodulatory Drugs (IMiDs)
3.2. Vaccination Strategies

{kind=link}
| Setting | TAA Target | Platform/Adjunct Treatment | Phase | Status | Identifier |
|---|---|---|---|---|---|
| MM (ISS I,II,III) | WT1 | ASCT | Not provided | R | NCT01827137 |
| Advanced myeloma | hTERT | ASCT | I/II | U | NCT00834665 |
| High risk myeloma | MAGE-A3 and NY-ESO-1 | DTPACE with ASCT | II/III | C | NCT00090493 |
| Various malignancies | NY-ESO-1 | Resiquimod and/or Poly-ICLC | I/II | C | NCT00948961 |
| Symptomatic ISS stage I,II,III myeloma | MAGE-A3 and AS15 | After ASCT | I | ONR | NCT01380145 |
| Early stage (ISS—I) myeloma | Id (DC based ) | n/a | I | C | NCT00988312 |
| Late stage (ISS—II,III) myeloma | Id pulsed DC | Tandem auto/allo SCT | I/II | C | NCT00186316 |
| MM >12 months of therapy | Id-KLH | CD3/CD28 activated T cells ASCT | II | R | NCT01426828 |
| MM | MUC1 | GM-CSF | II | W | NCT00162500 |
| Various malignancies | MUC1 | hGM-CSF | I/II | C | NCT01232712 |
3.3. Antibody Therapies
3.3.1. Elotuzumab; Anti-CS1 (SLAMF7) Antibody
3.3.2. Daratumumab; Anti-CD38 Antibody

3.3.3. Bispecific T Cell Engagers (BiTEs)
3.4. Immune Checkpoint Inhibitors (PD-1/PD-L1 Axis)
| Setting | PD1 Antibody | IMiD | Additional Intervention | Phase | Status | Identifier |
|---|---|---|---|---|---|---|
| NDMM | Pembrolizumab | Lenalidomide | n/a | III | R | NCT02579863 |
| RRMM | Pembrolizumab | Lenalidomide | n/a | I | R | NCT02036502 |
| RRMM | Pembrolizumab | Pomalidomide | n/a | I/II | R | NCT02289222 |
| RRMM | Pembrolizumab | Pomalidomide | n/a | III | R | NCT02576977 |
| RRMM | Pidilizumab | Lenalidomide | n/a | I/II | R | NCT02077959 |
| Post ASCT | Pembrolizumab | Lenalidomide | n/a | II | R | NCT02331368 |
| RRMM | Nivolumab | n/a | Ipilimumab Lirilumab | I | R | NCT01592370 |
| Post ASCT | Pidilizumab | n/a | DC/MM | II | ONR | NCT01067287 |
| Locally advanced/metastatic solid tumors or hematological malignancies | MPDL3280A | n/a | n/a | I | R | NCT01375842 |
| MM | MPDL3280A | Lenalidomide | n/a | Ib | R | NCT02431208 |
3.5. Adoptive T Cell Therapies (ACT)
3.6. Chimeric Antigen Receptor (CAR) T Cells
3.7. TCR Transgenic T Cells
3.8. Cytokines
3.8.1. Interleukin–6 (IL-6)
3.8.2. Interleukin-15 (IL-15)
| Setting | Cytokine | Inhibitor | Additional Intervention | Phase | Status | Identifier |
|---|---|---|---|---|---|---|
| NDMM | IL-6 | CNTO 328 | VMP combination | II | C | NCT00911859 |
| MGUS, SMM, indolent MM | IL-6 | CNTO 328 | Cardiac functions | I | C | NCT01219010 |
| High risk smoldering | IL-6 | CNTO 328 | n/a | II | ONR | NCT01484275 |
| RRMM | IL-6 | CNTO 328 | VD | III | W | NCT01266811 |
| NDMM | IL-6 | CNTO 328 | VRD | Ib/II | C | NCT01531998 |
| MM, NHL, Castleman’s disease | IL-6 | CNTO 328 | n/a | I | C | NCT00412321 |
| RRMM | IL-6 | CNTO 328 | Dexamethasone | II | C | NCT00402181 |
| RRMM | IL-6 | CNTO 328 | Bortezomib | II | ONR | NCT00401843 |
| RRMM | IL-15 | ALT-803 | n/a | I/II | R | NCT02099539 |
| Post allogeneic stem cell transplantation | IL-15 | ALT-803 | n/a | I/II | R | NCT01885897 |
4. Future
Conflicts of Interest
References
- Korde, N.; Kristinsson, S.Y.; Landgren, O. Monoclonal gammopathy of undetermined significance (MGUS) and smoldering multiple myeloma (SMM): Novel biological insights and development of early treatment strategies. Blood 2011, 117, 5573–5581. [Google Scholar] [CrossRef] [PubMed]
- Raitakari, M.; Brown, R.D.; Gibson, J.; Joshua, D.E. T cells in myeloma. Hematol. Oncol. 2003, 21, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Dhodapkar, M.V.; Krasovsky, J.; Osman, K.; Geller, M.D. Vigorous premalignancy-specific effector T cell response in the bone marrow of patients with monoclonal gammopathy. J. Exp. Med. 2003, 198, 1753–1757. [Google Scholar] [CrossRef] [PubMed]
- Prabhala, R.H.; Pelluru, D.; Fulciniti, M.; Prabhala, H.K.; Nanjappa, P.; Song, W.; Pai, C.; Amin, S.; Tai, Y.T.; Richardson, P.G.; et al. Elevated IL-17 produced by TH17 cells promotes myeloma cell growth and inhibits immune function in multiple myeloma. Blood 2010, 115, 5385–5392. [Google Scholar] [CrossRef] [PubMed]
- Noonan, K.; Borrello, I. The immune microenvironment of myeloma. Cancer Microenviron. 2011, 4, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Mitsdoerffer, M.; Croxford, A.L.; Awasthi, A.; Dardalhon, V.A.; Galileos, G.; Vollmar, P.; Stritesky, G.L.; Kaplan, M.H.; Waisman, A.; et al. IL-6 controls Th17 immunity in vivo by inhibiting the conversion of conventional T cells into Foxp3+ regulatory T cells. Proc. Natl. Acad. Sci. USA 2008, 105, 18460–18465. [Google Scholar] [CrossRef] [PubMed]
- Ratta, M.; Fagnoni, F.; Curti, A.; Vescovini, R.; Sansoni, P.; Oliviero, B.; Fogli, M.; Ferri, E.; Della Cuna, G.R.; Tura, S.; et al. Dendritic cells are functionally defective in multiple myeloma: The role of interleukin-6. Blood 2002, 100, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Hideshima, T.; Akiyama, M.; Raje, N.; Richardson, P.; Chauhan, D.; Anderson, K.C. Ex vivo induction of multiple myeloma-specific cytotoxic T lymphocytes. Blood 2003, 102, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Kristinsson, S.Y.; Tang, M.; Pfeiffer, R.M.; Bjorkholm, M.; Goldin, L.R.; Blimark, C.; Mellqvist, U.H.; Wahlin, A.; Turesson, I.; Landgren, O. Monoclonal gammopathy of undetermined significance and risk of infections: A population-based study. Haematologica 2012, 97, 854–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huff, C.A.; Fuchs, E.J.; Noga, S.J.; O'Donnell, P.V.; Ambinder, R.F.; Diehl, L.; Borrello, I.; Vogelsang, G.B.; Miller, C.B.; Flinn, I.A.; et al. Long-term follow-up of T cell-depleted allogeneic bone marrow transplantation in refractory multiple myeloma: Importance of allogeneic T cells. Biol. Blood Marrow Transplant. 2003, 9, 312–319. [Google Scholar] [CrossRef]
- Tricot, G.; Vesole, D.H.; Jagannath, S.; Hilton, J.; Munshi, N.; Barlogie, B. Graft-versus-myeloma effect: Proof of principle. Blood 1996, 87, 1196–1198. [Google Scholar] [PubMed]
- Verdonck, L.F.; Lokhorst, H.M.; Dekker, A.W.; Nieuwenhuis, H.K.; Petersen, E.J. Graft-versus-myeloma effect in two cases. Lancet 1996, 347, 800–801. [Google Scholar] [CrossRef]
- Lu, G.; Middleton, R.E.; Sun, H.; Naniong, M.; Ott, C.J.; Mitsiades, C.S.; Wong, K.K.; Bradner, J.E.; Kaelin, W.G., Jr. The myeloma drug lenalidomide promotes the cereblon-dependent destruction of Ikaros proteins. Science 2014, 343, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Quach, H.; Ritchie, D.; Stewart, A.K.; Neeson, P.; Harrison, S.; Smyth, M.J.; Prince, H.M. Mechanism of action of immunomodulatory drugs (IMiDS) in multiple myeloma. Leukemia 2010, 24, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Galustian, C.; Meyer, B.; Labarthe, M.C.; Dredge, K.; Klaschka, D.; Henry, J.; Todryk, S.; Chen, R.; Muller, G.; Stirling, D.; et al. The anti-cancer agents lenalidomide and pomalidomide inhibit the proliferation and function of T regulatory cells. Cancer Immunol. Immunother. 2009, 58, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Yi, Q.; Szmania, S.; Freeman, J.; Qian, J.; Rosen, N.A.; Viswamitra, S.; Cottler-Fox, M.; Barlogie, B.; Tricot, G.; van Rhee, F. Optimizing dendritic cell-based immunotherapy in multiple myeloma: Intranodal injections of idiotype-pulsed CD40 ligand-matured vaccines led to induction of type-1 and cytotoxic T-cell immune responses in patients. Br. J. Haematol. 2010, 150, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Titzer, S.; Christensen, O.; Manzke, O.; Tesch, H.; Wolf, J.; Emmerich, B.; Carsten, C.; Diehl, V.; Bohlen, H. Vaccination of multiple myeloma patients with idiotype-pulsed dendritic cells: Immunological and clinical aspects. Br. J. Haematol. 2000, 108, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Lacy, M.Q.; Mandrekar, S.; Dispenzieri, A.; Hayman, S.; Kumar, S.; Buadi, F.; Dingli, D.; Litzow, M.; Wettstein, P.; Padley, D.; et al. Idiotype-pulsed antigen-presenting cells following autologous transplantation for multiple myeloma may be associated with prolonged survival. Am. J. Hematol. 2009, 84, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vacchelli, E.; Bravo-San Pedro, J.M.; Buque, A.; Senovilla, L.; Baracco, E.E.; Bloy, N.; Castoldi, F.; Abastado, J.P.; Agostinis, P.; et al. Classification of current anticancer immunotherapies. Oncotarget 2014, 5, 12472–12508. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Smith, R.; Daley, J.; Mimura, N.; Tai, Y.T.; Anderson, K.C.; Munshi, N.C. Myeloma-specific multiple peptides able to generate cytotoxic T lymphocytes: A potential therapeutic application in multiple myeloma and other plasma cell disorders. Clin. Cancer Res. 2012, 18, 4850–4860. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Prabhala, R.; Voskertchian, A.; Brown, A.; Maguire, C.; Richardson, P.; Dranoff, G.; Anderson, K.C.; Munshi, N.C. A multiepitope of XBP1, CD138 and CS1 peptides induces myeloma-specific cytotoxic T lymphocytes in T cells of smoldering myeloma patients. Leukemia 2015, 29, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Vasir, B.; Borges, V.; Wu, Z.; Grosman, D.; Rosenblatt, J.; Irie, M.; Anderson, K.; Kufe, D.; Avigan, D. Fusion of dendritic cells with multiple myeloma cells results in maturation and enhanced antigen presentation. Br. J. Haematol. 2005, 129, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, J.; Vasir, B.; Uhl, L.; Blotta, S.; Macnamara, C.; Somaiya, P.; Wu, Z.; Joyce, R.; Levine, J.D.; Dombagoda, D.; et al. Vaccination with dendritic cell/tumor fusion cells results in cellular and humoral antitumor immune responses in patients with multiple myeloma. Blood 2011, 117, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Rosenblatt, J.; Avivi, I.; Vasir, B.; Uhl, L.; Munshi, N.C.; Katz, T.; Dey, B.R.; Somaiya, P.; Mills, H.; Campigotto, F.; et al. Vaccination with dendritic cell/tumor fusions following autologous stem cell transplant induces immunologic and clinical responses in multiple myeloma patients. Clin. Cancer Res. 2013, 19, 3640–3648. [Google Scholar] [CrossRef] [PubMed]
- Hsi, E.D.; Steinle, R.; Balasa, B.; Szmania, S.; Draksharapu, A.; Shum, B.P.; Huseni, M.; Powers, D.; Nanisetti, A.; Zhang, Y.; et al. CS1, a potential new therapeutic antibody target for the treatment of multiple myeloma. Clin. Cancer Res. 2008, 14, 2775–2784. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.M.; Bakan, C.E.; Swartzel, G.D.; Hofmeister, C.C.; Efebera, Y.A.; Kwon, H.; Starling, G.C.; Ciarlariello, D.; Bhaskar, S.; Briercheck, E.L.; et al. Elotuzumab directly enhances NK cell cytotoxicity against myeloma via CS1 ligation: Evidence for augmented NK cell function complementing ADCC. Cancer Immunol. Immunother. 2013, 62, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.T.; Dillon, M.; Song, W.; Leiba, M.; Li, X.F.; Burger, P.; Lee, A.I.; Podar, K.; Hideshima, T.; Rice, A.G.; et al. Anti-CS1 humanized monoclonal antibody HuLuc63 inhibits myeloma cell adhesion and induces antibody-dependent cellular cytotoxicity in the bone marrow milieu. Blood 2008, 112, 1329–1337. [Google Scholar] [CrossRef] [PubMed]
- Zonder, J.A.; Mohrbacher, A.F.; Singhal, S.; van Rhee, F.; Bensinger, W.I.; Ding, H.; Fry, J.; Afar, D.E.; Singhal, A.K. A phase 1, multicenter, open-label, dose escalation study of elotuzumab in patients with advanced multiple myeloma. Blood 2012, 120, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Lonial, S.; Vij, R.; Harousseau, J.L.; Facon, T.; Moreau, P.; Mazumder, A.; Kaufman, J.L.; Leleu, X.; Tsao, L.C.; Westland, C.; et al. Elotuzumab in combination with lenalidomide and low-dose dexamethasone in relapsed or refractory multiple myeloma. J. Clin. Oncol. 2012, 30, 1953–1959. [Google Scholar] [CrossRef] [PubMed]
- Lonial, S.; Dimopoulos, M.; Palumbo, A.; White, D.; Grosicki, S.; Spicka, I.; Walter-Croneck, A.; Moreau, P.; Mateos, M.V.; Magen, H.; et al. ELOQUENT-2 Investigators Elotuzumab Therapy for Relapsed or Refractory Multiple Myeloma. N. Engl. J. Med. 2015, 373, 621–631. [Google Scholar] [CrossRef] [PubMed]
- De Weers, M.; Tai, Y.T.; van der Veer, M.S.; Bakker, J.M.; Vink, T.; Jacobs, D.C.; Oomen, L.A.; Peipp, M.; Valerius, T.; Slootstra, J.W.; et al. Daratumumab, a novel therapeutic human CD38 monoclonal antibody, induces killing of multiple myeloma and other hematological tumors. J. Immunol. 2011, 186, 1840–1848. [Google Scholar] [CrossRef] [PubMed]
- Lokhorst, H.M.; Plesner, T.; Laubach, J.P.; Nahi, H.; Gimsing, P.; Hansson, M.; Minnema, M.C.; Lassen, U.; Krejcik, J.; Palumbo, A.; et al. Targeting CD38 with daratumumab monotherapy in multiple myeloma. N. Engl. J. Med. 2015, 373, 1207–1219. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, B.P.; Raman, V.S.; Erickson, L.D.; Cook, W.J.; Weaver, L.K.; Ahonen, C.; Lin, L.L.; Mantchev, G.T.; Bram, R.J.; Noelle, R.J. BCMA is essential for the survival of long-lived bone marrow plasma cells. J. Exp. Med. 2004, 199, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Tassone, P.; Goldmacher, V.S.; Neri, P.; Gozzini, A.; Shammas, M.A.; Whiteman, K.R.; Hylander-Gans, L.L.; Carrasco, D.R.; Hideshima, T.; Shringarpure, R.; et al. Cytotoxic activity of the maytansinoid immunoconjugate B-B4-DM1 against CD138+ multiple myeloma cells. Blood 2004, 104, 3688–3696. [Google Scholar] [CrossRef] [PubMed]
- Kontermann, R.E.; Brinkmann, U. Bispecific antibodies. Drug Discov. Today 2015, 20, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Honemann, D.; Kufer, P.; Rimpler, M.M.; Chatterjee, M.; Friedl, S.; Riecher, F.; Bommert, K.; Dorken, B.; Bargou, R.C. A novel recombinant bispecific single-chain antibody, bscWue-1 × CD3, induces T-cell-mediated cytotoxicity towards human multiple myeloma cells. Leukemia 2004, 18, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Chen, D.; Zong, Y.; Ye, S.; Tang, J.; Meng, H.; An, G.; Zhang, X.; Yang, L. Immunotherapy based on bispecific T-cell engager with hIgG1 Fc sequence as a new therapeutic strategy in multiple myeloma. Cancer Sci. 2015, 106, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Bhutani, D.; Lum, L.G. Activated T cells armed with bispecific antibodies kill tumor targets. Curr. Opin. Hematol. 2015, 22, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Tai, Y.T.; Anderson, K.C. Targeting B-cell maturation antigen in multiple myeloma. Immunotherapy 2015, 7, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.S.; Stojanov, P.; Polak, P.; Kryukov, G.V.; Cibulskis, K.; Sivachenko, A.; Carter, S.L.; Stewart, C.; Mermel, C.H.; Roberts, S.A.; et al. Mutational heterogeneity in cancer and the search for new cancer-associated genes. Nature 2013, 499, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.M., Jr.; Bakan, C.E.; Mishra, A.; Hofmeister, C.C.; Efebera, Y.; Becknell, B.; Baiocchi, R.A.; Zhang, J.; Yu, J.; Smith, M.K.; et al. The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: A therapeutic target for CT-011, a novel monoclonal anti-PD-1 antibody. Blood 2010, 116, 2286–2294. [Google Scholar] [CrossRef] [PubMed]
- Lesokhin, A.; Ansell, S.; Armand, P.; Scott, E.; Halwani, A.; Gutierrez, M.; Millenson, M.; Cohen, A.; Schuster, S.; Lebovic, D.; et al. Preliminary results of a phase I study of Nivolumab (BMS-936558) in patients with relapsed or refractory lymphoid malignancies. Blood 2014, 124, 291. [Google Scholar]
- Berger, R.; Rotem-Yehudar, R.; Slama, G.; Landes, S.; Kneller, A.; Leiba, M.; Koren-Michowitz, M.; Shimoni, A.; Nagler, A. Phase I safety and pharmacokinetic study of CT-011, a humanized antibody interacting with PD-1, in patients with advanced hematologic malignancies. Clin. Cancer Res. 2008, 14, 3044–3051. [Google Scholar] [CrossRef] [PubMed]
- Badros, A.; Kocoglu, M.; Ma, N.; Rapoport, A.; Lederer, E.; Philip, S.; Lesho, P.D.C.; Hardy, N.; Yared, J.; Goloubeva, O.; Singh, Z. A phase II study of anti PD-1 antibody pembrolizumab, pomalidomide and dexamethasone in patients with relapsed/refractory multiple myeloma (RRMM). Blood 2015, 126, 506. [Google Scholar]
- Yousef, S.; Marvin, J.; Steinbach, M.; Langemo, A.; Kovacsovics, T.; Binder, M.; Kroger, N.; Luetkens, T.; Atanackovic, D. Immunomodulatory molecule PD-L1 is expressed on malignant plasma cells and myeloma-propagating pre-plasma cells in the bone marrow of multiple myeloma patients. Blood Cancer J. 2015, 5, e285. [Google Scholar] [CrossRef] [PubMed]
- Raitakari, M.; Brown, R.D.; Sze, D.; Yuen, E.; Barrow, L.; Nelson, M.; Pope, B.; Esdale, W.; Gibson, J.; Joshua, D.E. T-cell expansions in patients with multiple myeloma have a phenotype of cytotoxic T cells. Br. J. Haematol. 2000, 110, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Porrata, L.F.; Gertz, M.A.; Inwards, D.J.; Litzow, M.R.; Lacy, M.Q.; Tefferi, A.; Gastineau, D.A.; Dispenzieri, A.; Ansell, S.M.; Micallef, I.N.; et al. Early lymphocyte recovery predicts superior survival after autologous hematopoietic stem cell transplantation in multiple myeloma or non-Hodgkin lymphoma. Blood 2001, 98, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, A.P.; Aqui, N.A.; Stadtmauer, E.A.; Vogl, D.T.; Xu, Y.Y.; Kalos, M.; Cai, L.; Fang, H.B.; Weiss, B.M.; Badros, A.; et al. Combination immunotherapy after ASCT for multiple myeloma using MAGE-A3/Poly-ICLC immunizations followed by adoptive transfer of vaccine-primed and costimulated autologous T cells. Clin. Cancer Res. 2014, 20, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- Noonan, K.A.; Huff, C.A.; Davis, J.; Lemas, M.V.; Fiorino, S.; Bitzan, J.; Ferguson, A.; Emerling, A.; Luznik, L.; Matsui, W.; et al. Adoptive transfer of activated marrow-infiltrating lymphocytes induces measurable antitumor immunity in the bone marrow in multiple myeloma. Sci. Transl. Med. 2015, 7, 288ra78. [Google Scholar] [CrossRef] [PubMed]
- Garfall, A.L.; Maus, M.V.; Hwang, W.T.; Lacey, S.F.; Mahnke, Y.D.; Melenhorst, J.J.; Zheng, Z.; Vogl, D.T.; Cohen, A.D.; Weiss, B.M.; et al. Chimeric antigen receptor T cells against CD19 for multiple myeloma. N. Engl. J. Med. 2015, 373, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, R.O.; Evbuomwan, M.O.; Pittaluga, S.; Rose, J.J.; Raffeld, M.; Yang, S.; Gress, R.E.; Hakim, F.T.; Kochenderfer, J.N. B-cell maturation antigen is a promising target for adoptive T-cell therapy of multiple myeloma. Clin. Cancer Res. 2013, 19, 2048–2060. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, A.P.; Stadtmauer, E.A.; Binder-Scholl, G.K.; Goloubeva, O.; Vogl, D.T.; Lacey, S.F.; Badros, A.Z.; Garfall, A.; Weiss, B.; Finklestein, J.; et al. NY-ESO-1-specific TCR-engineered T cells mediate sustained antigen-specific antitumor effects in myeloma. Nat. Med. 2015, 21, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Van Oers, M.H.; van Zaanen, H.C.; Lokhorst, H.M. Interleukin-6, a new target for therapy in multiple myeloma? Ann. Hematol. 1993, 66, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Van Zaanen, H.C.; Koopmans, R.P.; Aarden, L.A.; Rensink, H.J.; Stouthard, J.M.; Warnaar, S.O.; Lokhorst, H.M.; van Oers, M.H. Endogenous interleukin 6 production in multiple myeloma patients treated with chimeric monoclonal anti-IL6 antibodies indicates the existence of a positive feed-back loop. J. Clin. Investig. 1996, 98, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, R.Z.; Gercheva, L.; Williams, C.; Sutherland, H.; Robak, T.; Masszi, T.; Goranova-Marinova, V.; Dimopoulos, M.A.; Cavenagh, J.D.; Spicka, I.; et al. A phase 2, randomized, double-blind, placebo-controlled study of siltuximab (anti-IL-6 mAb) and bortezomib versus bortezomib alone in patients with relapsed or refractory multiple myeloma. Am. J. Hematol. 2015, 90, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.C.; Jeng, E.K.; Rhode, P.R. The IL-15-based superagonist ALT-803 promotes the antigen-independent conversion of memory CD8 T cells into innate-like effector cells with antitumor activity. Oncoimmunology 2013, 2, e26442. [Google Scholar] [CrossRef] [PubMed]
- Paiva, B.; van Dongen, J.J.; Orfao, A. New criteria for response assessment: Role of minimal residual disease in multiple myeloma. Blood 2015, 125, 3059–3068. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocoglu, M.; Badros, A. The Role of Immunotherapy in Multiple Myeloma. Pharmaceuticals 2016, 9, 3. https://doi.org/10.3390/ph9010003
Kocoglu M, Badros A. The Role of Immunotherapy in Multiple Myeloma. Pharmaceuticals. 2016; 9(1):3. https://doi.org/10.3390/ph9010003
Chicago/Turabian StyleKocoglu, Mehmet, and Ashraf Badros. 2016. "The Role of Immunotherapy in Multiple Myeloma" Pharmaceuticals 9, no. 1: 3. https://doi.org/10.3390/ph9010003
APA StyleKocoglu, M., & Badros, A. (2016). The Role of Immunotherapy in Multiple Myeloma. Pharmaceuticals, 9(1), 3. https://doi.org/10.3390/ph9010003