
Nociceptive TRP Channels: Sensory Detectors and Transducers in Multiple Pain Pathologies

Abstract
:1. Introduction
2. Nociceptive TRP Channels
2.1. History, Identification and Cloning
2.2. Characterization of Channel Function: Activation by Diverse Physico-Chemical Stimuli
2.3. Similarities and Differences in Channel Expression and Localization
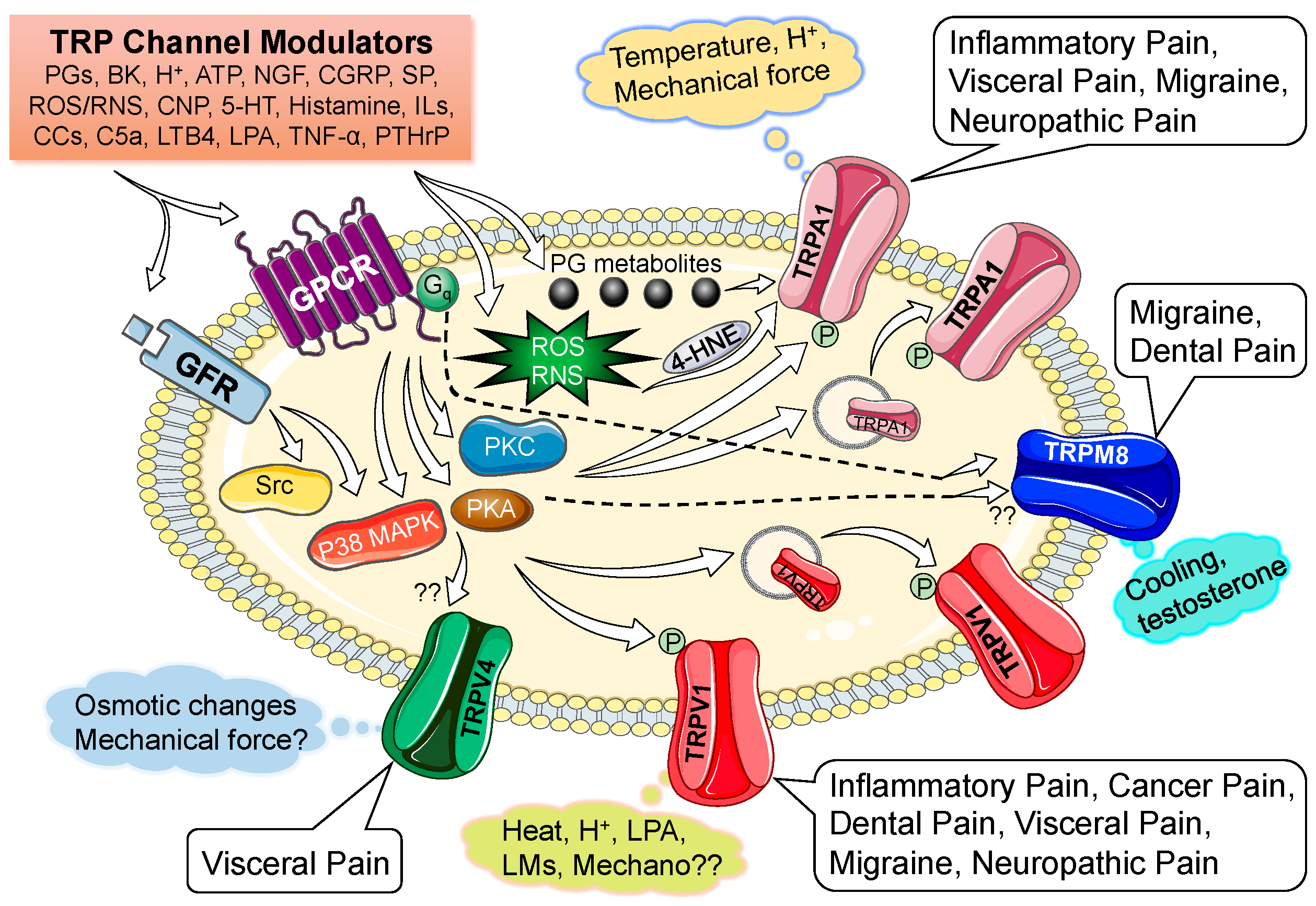
3. Modulation of Nociceptive TRP Channel Activity and Expression
4. Involvement of Nociceptive TRP Channels in Painful Pathologies
4.1. Inflammatory Pain
4.2. Neuropathic Pain
4.3. Visceral Pain
4.4. Pain Associated with Cancers and Other Pathological Conditions
4.4.1. Cancer Pain
4.4.2. Dental Pain
4.4.3. Migraine
5. Nociceptive TRP Channels in Non-Painful Pathologies and Physiological Processes
6. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Loeser, J.D.; Treede, R.D. The kyoto protocol of IASP basic pain terminology. Pain 2008, 137, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Cosens, D.J.; Manning, A. Abnormal electroretinogram from a drosophila mutant. Nature 1969, 224, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Minke, B. The history of the drosophila TRP channel: The birth of a new channel superfamily. J. Neurogenet. 2010, 24, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.J.; Sweet, T.B.; Clapham, D.E. International union of basic and clinical pharmacology. LXXVI. Current progress in the mammalian TRP ion channel family. Pharmacol. Rev. 2010, 62, 381–404. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Montell, C. Physiology, phylogeny, and functions of the TRP superfamily of cation channels. Sci. STKE 2001, 90, re1. [Google Scholar] [CrossRef] [PubMed]
- Benemei, S.; De Cesaris, F.; Fusi, C.; Rossi, E.; Lupi, C.; Geppetti, P. TRPA1 and other TRP channels in migraine. J. Headache Pain 2013, 14, 71. [Google Scholar] [CrossRef] [PubMed]
- Julius, D. TRP channels and pain. Ann. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef] [PubMed]
- Meents, J.E.; Neeb, L.; Reuter, U. TRPV1 in migraine pathophysiology. Trends Mol. Med. 2010, 16, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Sensory TRP channels: The key transducers of nociception and pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 73–118. [Google Scholar] [PubMed]
- Moran, M.M.; McAlexander, M.A.; Biro, T.; Szallasi, A. Transient receptor potential channels as therapeutic targets. Nat. Rev. Drug Discov. 2011, 10, 601–620. [Google Scholar] [CrossRef] [PubMed]
- Patapoutian, A.; Tate, S.; Woolf, C.J. Transient receptor potential channels: Targeting pain at the source. Nat. Rev. Drug Discov. 2009, 8, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Sherrington, C. The Integrative Action of the Nervous System; Oxford University Press: Oxford, UK, 1906. [Google Scholar]
- Dray, A. Mechanism of action of capsaicin-like molecules on sensory neurons. Life Sci. 1992, 51, 1759–1765. [Google Scholar] [CrossRef]
- Amann, R.; Maggi, C.A. Ruthenium red as a capsaicin antagonist. Life Sci. 1991, 49, 849–856. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [PubMed]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef]
- Caterina, M.J.; Rosen, T.A.; Tominaga, M.; Brake, A.J.; Julius, D. A capsaicin-receptor homologue with a high threshold for noxious heat. Nature 1999, 398, 436–441. [Google Scholar] [PubMed]
- Liedtke, W.; Choe, Y.; Marti-Renom, M.A.; Bell, A.M.; Denis, C.S.; Sali, A.; Hudspeth, A.J.; Friedman, J.M.; Heller, S. Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate osmoreceptor. Cell 2000, 103, 525–535. [Google Scholar] [CrossRef]
- Strotmann, R.; Harteneck, C.; Nunnenmacher, K.; Schultz, G.; Plant, T.D. OTRPC4, a nonselective cation channel that confers sensitivity to extracellular osmolarity. Nat. Cell Biol. 2000, 2, 695–702. [Google Scholar] [PubMed]
- Muller, D.; Hoenderop, J.G.; Meij, I.C.; van den Heuvel, L.P.; Knoers, N.V.; den Hollander, A.I.; Eggert, P.; Garcia-Nieto, V.; Claverie-Martin, F.; Bindels, R.J. Molecular cloning, tissue distribution, and chromosomal mapping of the human epithelial Ca2+ channel (ECAC1). Genomics 2000, 67, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.B.; Chen, X.Z.; Berger, U.V.; Vassilev, P.M.; Tsukaguchi, H.; Brown, E.M.; Hediger, M.A. Molecular cloning and characterization of a channel-like transporter mediating intestinal calcium absorption. J. Biol. Chem. 1999, 274, 22739–22746. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.; Erben, R.G.; Rump, A.; Adamski, J. Gene structure and regulation of the murine epithelial calcium channels ECAC1 and 2. Biochem. Biophys. Res. Commun. 2001, 289, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Flonta, M.L. Physiology. Cold current in thermoreceptive neurons. Nature 2001, 413, 480. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Reeve, A.J.; Andersson, D.A.; Moqrich, A.; Earley, T.J.; Hergarden, A.C.; Story, G.M.; Colley, S.; Hogenesch, J.B.; McIntyre, P.; et al. A heat-sensitive TRP channel expressed in keratinocytes. Science 2002, 296, 2046–2049. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ramsey, I.S.; Kotecha, S.A.; Moran, M.M.; Chong, J.A.; Lawson, D.; Ge, P.; Lilly, J.; Silos-Santiago, I.; Xie, Y.; et al. TRPV3 is a calcium-permeable temperature-sensitive cation channel. Nature 2002, 418, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Jaquemar, D.; Schenker, T.; Trueb, B. An ankyrin-like protein with transmembrane domains is specifically lost after oncogenic transformation of human fibroblasts. J. Biol. Chem. 1999, 274, 7325–7333. [Google Scholar] [CrossRef] [PubMed]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef]
- Grimm, C.; Kraft, R.; Sauerbruch, S.; Schultz, G.; Harteneck, C. Molecular and functional characterization of the melastatin-related cation channel TRPM3. J. Biol. Chem. 2003, 278, 21493–21501. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.F.; Loch, S.; Lambert, S.; Straub, I.; Mannebach, S.; Mathar, I.; Dufer, M.; Lis, A.; Flockerzi, V.; Philipp, S.E.; et al. Transient receptor potential M3 channels are ionotropic steroid receptors in pancreatic beta cells. Nat. Cell Biol. 2008, 10, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Appendino, G.; Nilius, B. Pharmacology of vanilloid transient receptor potential cation channels. Mol. Pharmacol. 2009, 75, 1262–1279. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 is a nociceptor channel involved in the detection of noxious heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Jordt, S.E.; Tominaga, M.; Julius, D. Acid potentiation of the capsaicin receptor determined by a key extracellular site. Proc. Natl. Acad. Sci. USA 2000, 97, 8134–8139. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, A.; Uzzell, V.; Dubin, A.E.; Mathur, J.; Petrus, M.; Bandell, M.; Patapoutian, A. TRPV1 is activated by both acidic and basic ph. J. Neurosci. 2009, 29, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Bisogno, T.; Trevisani, M.; Al-Hayani, A.; De Petrocellis, L.; Fezza, F.; Tognetto, M.; Petros, T.J.; Krey, J.F.; Chu, C.J.; et al. An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc. Natl. Acad. Sci. USA 2002, 99, 8400–8405. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.W.; Cho, H.; Kwak, J.; Lee, S.Y.; Kang, C.J.; Jung, J.; Cho, S.; Min, K.H.; Suh, Y.G.; Kim, D.; et al. Direct activation of capsaicin receptors by products of lipoxygenases: Endogenous capsaicin-like substances. Proc. Natl. Acad. Sci. USA 2000, 97, 6155–6160. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, P.M.; Petersson, J.; Andersson, D.A.; Chuang, H.; Sorgard, M.; Di Marzo, V.; Julius, D.; Hogestatt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [PubMed]
- Kozai, D.; Ogawa, N.; Mori, Y. Redox regulation of transient receptor potential channels. Antioxid. Redox Ssignal. 2014, 21, 971–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, M.-K.; Guler, A.D.; Caterina, M.J. TRPV1 shows dynamic ionic selectivity during agonist stimulation. Nat. Neurosci. 2008, 11, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, D.P.; Wang, S.Y.; Wang, G.K.; Nau, C. A tyrosine residue in TM6 of the vanilloid receptor TRPV1 involved in desensitization and calcium permeability of capsaicin-activated currents. Mol. Cell. Neurosci. 2003, 23, 314–324. [Google Scholar] [CrossRef]
- Koplas, P.A.; Rosenberg, R.L.; Oxford, G.S. The role of calcium in the desensitization of capsaicin responses in rat dorsal root ganglion neurons. J. Neurosci. 1997, 17, 3525–3537. [Google Scholar] [PubMed]
- Loo, L.; Shepherd, A.J.; Mickle, A.D.; Lorca, R.A.; Shutov, L.P.; Usachev, Y.M.; Mohapatra, D.P. The C-type natriuretic peptide induces thermal hyperalgesia through a noncanonical gbetagamma-dependent modulation of TRPV1 channel. J. Neurosci. 2012, 32, 11942–11955. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Zhu, M.X. TRPV3. In Handbook of Experimental Pharmacology; Springer: Berlin, Germany, 2014; Volume 222, pp. 273–291. [Google Scholar]
- Bourinet, E.; Altier, C.; Hildebrand, M.E.; Trang, T.; Salter, M.W.; Zamponi, G.W. Calcium-permeable ion channels in pain signaling. Physiol. Rev. 2014, 94, 81–140. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.C.; Horn, N.A.; Huynh, T.; Kelava, L.; Lansman, J.B. Evidence TRPV4 contributes to mechanosensitive ion channels in mouse skeletal muscle fibers. Channels 2012, 6, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Haber, N.; Joseph, E.; Dina, O.A.; Liedtke, W.; Levine, J.D. TRPV4 mediates pain-related behavior induced by mild hypertonic stimuli in the presence of inflammatory mediator. Pain 2005, 118, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Haber, N.; Yeh, J.J.; Boyd, A.E.; Parada, C.A.; Chen, X.; Reichling, D.B.; Levine, J.D. Hypotonicity induces TRPV4-mediated nociception in rat. Neuron 2003, 39, 497–511. [Google Scholar] [CrossRef]
- Yudin, Y.; Rohacs, T. Regulation of TRPM8 channel activity. Mol. Cell. Endocrinol. 2012, 353, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Asuthkar, S.; Elustondo, P.A.; Demirkhanyan, L.; Sun, X.; Baskaran, P.; Velpula, K.K.; Thyagarajan, B.; Pavlov, E.V.; Zakharian, E. The TRPM8 protein is a testosterone receptor: I. Biochemical evidence for direct TRPM8-testosterone interactions. J. Biol. Chem. 2015, 290, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Asuthkar, S.; Demirkhanyan, L.; Sun, X.; Elustondo, P.A.; Krishnan, V.; Baskaran, P.; Velpula, K.K.; Thyagarajan, B.; Pavlov, E.V.; Zakharian, E. The TRPM8 protein is a testosterone receptor: II. Functional evidence for an ionotropic effect of testosterone on TRPM8. J. Biol. Chem. 2015, 290, 2670–2688. [Google Scholar] [CrossRef] [PubMed]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Karashima, Y.; Talavera, K.; Everaerts, W.; Janssens, A.; Kwan, K.Y.; Vennekens, R.; Nilius, B.; Voets, T. TRPA1 acts as a cold sensor in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 1273–1278. [Google Scholar] [CrossRef] [PubMed]
- Dunham, J.; Leith, J.; Lumb, B.; Donaldson, L. Transient receptor potential channel A1 and noxious cold responses in rat cutaneous nociceptors. Neuroscience 2010, 165, 1412–1419. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kang, D.; Xu, J.; Lake, M.; Hogan, J.O.; Sun, C.; Walter, K.; Yao, B.; Kim, D. Species differences and molecular determinant of TRPA1 cold sensitivity. Nat. Commun. 2013, 4, 2501. [Google Scholar] [CrossRef] [PubMed]
- Moparthi, L.; Kichko, T.I.; Eberhardt, M.; Hogestatt, E.D.; Kjellbom, P.; Johanson, U.; Reeh, P.W.; Leffler, A.; Filipovic, M.R.; Zygmunt, P.M. Human TRPA1 is a heat sensor displaying intrinsic u-shaped thermosensitivity. Sci. Rep. 2016, 6, 28763. [Google Scholar] [CrossRef] [PubMed]
- Jabba, S.; Goyal, R.; Sosa-Pagan, J.O.; Moldenhauer, H.; Wu, J.; Kalmeta, B.; Bandell, M.; Latorre, R.; Patapoutian, A.; Grandl, J. Directionality of temperature activation in mouse TRPA1 ion channel can be inverted by single-point mutations in ankyrin repeat six. Neuron 2014, 82, 1017–1031. [Google Scholar] [CrossRef] [PubMed]
- De la Roche, J.; Eberhardt, M.J.; Klinger, A.B.; Stanslowsky, N.; Wegner, F.; Koppert, W.; Reeh, P.W.; Lampert, A.; Fischer, M.J.M.; Leffler, A. The molecular basis for species-specific activation of human TRPA1 protein by protons involves poorly conserved residues within transmembrane domains 5 and 6. J. Biol. Chem. 2013, 288, 20280–20292. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Chang, R.B.; Liman, E.R. TRPA1 is a component of the nociceptive response to CO2. J. Neurosci. 2010, 30, 12958–12963. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Chang, R.B.; Allgood, S.D.; Silver, W.L.; Liman, E.R. A TRPA1-dependent mechanism for the pungent sensation of weak acids. J. Gen. Physiol. 2011, 137, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 2007, 445, 541–545. [Google Scholar] [PubMed]
- Macpherson, L.J.; Xiao, B.; Kwan, K.Y.; Petrus, M.J.; Dubin, A.E.; Hwang, S.; Cravatt, B.; Corey, D.P.; Patapoutian, A. An ion channel essential for sensing chemical damage. J. Neurosci. 2007, 27, 11412–11415. [Google Scholar] [CrossRef] [PubMed]
- Viana, F. TRPA1 channels: Molecular sentinels of cellular stress and tissue damage. J. Physiol. 2016, 594, 4151–4169. [Google Scholar] [PubMed]
- Trevisani, M.; Siemens, J.; Materazzi, S.; Bautista, D.M.; Nassini, R.; Campi, B.; Imamachi, N.; Andrè, E.; Patacchini, R.; Cottrell, G.S.; et al. 4-hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc. Natl. Acad. Sci. USA 2007, 104, 13519–13524. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Dubin, A.E.; Petrus, M.J.; Patapoutian, A. TRPV1 and TRPA1 mediate peripheral nitric oxide-induced nociception in mice. PLoS ONE 2009, 4, e7596. [Google Scholar] [CrossRef] [PubMed]
- Pozsgai, G.; Hajna, Z.; Bagoly, T.; Boros, M.; Kemény, Á.; Materazzi, S.; Nassini, R.; Helyes, Z.; Szolcsányi, J.; Pintér, E. The role of transient receptor potential ankyrin 1 (TRPA1) receptor activation in hydrogen-sulphide-induced CGRP-release and vasodilation. Eur. J. Pharmacol. 2012, 689, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, M.; Dux, M.; Namer, B.; Miljkovic, J.; Cordasic, N.; Will, C.; Kichko, T.I.; de la Roche, J.; Fischer, M.; Suarez, S.A.; et al. H2S and no cooperatively regulate vascular tone by activating a neuroendocrine HNO-TRPA1-CGRP signalling pathway. Nat. Commun. 2014, 5, 4381. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Mizuno, Y.; Kozai, D.; Yamamoto, S.; Kiyonaka, S.; Shibata, T.; Uchida, K.; Mori, Y. Molecular characterization of TRPA1 channel activation by cysteine-reactive inflammatory mediators. Channels 2008, 2, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Orengo, L.; Dhaka, A.; Heuermann, R.J.; Young, T.J.; Montana, M.C.; Cavanaugh, E.J.; Kim, D.; Story, G.M. Cutaneous nociception evoked by 15-delta PGJ2 via activation of ion channel TRPA1. Mol. Pain 2008, 4, 30. [Google Scholar] [CrossRef] [PubMed]
- Zurborg, S.; Yurgionas, B.; Jira, J.A.; Caspani, O.; Heppenstall, P.A. Direct activation of the ion channel TRPA1 by Ca2+. Nat. Neurosci. 2007, 10, 277–279. [Google Scholar] [CrossRef] [PubMed]
- Brierley, S.M.; Castro, J.; Harrington, A.M.; Hughes, P.A.; Page, A.J.; Rychkov, G.Y.; Blackshaw, L.A. TRPA1 contributes to specific mechanically activated currents and sensory neuron mechanical hypersensitivity. J. Physiol. 2011, 589, 3575–3593. [Google Scholar] [CrossRef] [PubMed]
- Vilceanu, D.; Stucky, C.L. TRPA1 mediates mechanical currents in the plasma membrane of mouse sensory neurons. PLoS ONE 2010, 5, e12177. [Google Scholar] [CrossRef] [PubMed]
- Brierley, S.M.; Hughes, P.A.; Page, A.J.; Kwan, K.Y.; Martin, C.M.; O’Donnell, T.A.; Cooper, N.J.; Harrington, A.M.; Adam, B.; Liebregts, T.; et al. The ion channel TRPA1 is required for normal mechanosensation and is modulated by algesic stimuli. Gastroenterology 2009, 137, 2084–2095 e2083. [Google Scholar] [CrossRef] [PubMed]
- Karashima, Y.; Prenen, J.; Talavera, K.; Janssens, A.; Voets, T.; Nilius, B. Agonist-induced changes in Ca2+ permeation through the nociceptor cation channel TRPA1. Biophys. J. 2010, 98, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, A.J.; Mohapatra, D.P. Tissue preparation and immunostaining of mouse sensory nerve fibers innervating skin and limb bones. J. Vis. Exp. 2012, 59, e3485. [Google Scholar] [CrossRef] [PubMed]
- Le Pichon, C.E.; Chesler, A.T. The functional and anatomical dissection of somatosensory subpopulations using mouse genetics. Front. Neuroanat. 2014, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Haber, N.; Dina, O.A.; Joseph, E.K.; Reichling, D.; Levine, J.D. A transient receptor potential vanilloid 4-dependent mechanism of hyperalgesia is engaged by concerted action of inflammatory mediators. J. Neurosci. 2006, 26, 3864–3874. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.; Kerby, A.; Aubdool, A.A.; Power, A.R.; Grover, S.; Gentry, C.; Grant, A.D. 4alpha-phorbol 12,13-didecanoate activates cultured mouse dorsal root ganglia neurons independently of TRPV4. Br. J. Pharmacol. 2013, 168, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Flegel, C.; Schobel, N.; Altmuller, J.; Becker, C.; Tannapfel, A.; Hatt, H.; Gisselmann, G. RNA-seq analysis of human trigeminal and dorsal root ganglia with a focus on chemoreceptors. PLoS ONE 2015, 10, e0128951. [Google Scholar] [CrossRef] [PubMed]
- Goswami, S.C.; Mishra, S.K.; Maric, D.; Kaszas, K.; Gonnella, G.L.; Clokie, S.J.; Kominsky, H.D.; Gross, J.R.; Keller, J.M.; Mannes, A.J.; et al. Molecular signatures of mouse TRPV1-lineage neurons revealed by RNA-seq transcriptome analysis. J. Pain 2014, 15, 1338–1359. [Google Scholar] [CrossRef] [PubMed]
- Sapio, M.R.; Goswami, S.C.; Gross, J.R.; Mannes, A.J.; Iadarola, M.J. Transcriptomic analyses of genes and tissues in inherited sensory neuropathies. Exp. Neurol. 2016, 283, 375–395. [Google Scholar] [CrossRef] [PubMed]
- Ferrandiz-Huertas, C.; Mathivanan, S.; Wolf, C.J.; Devesa, I.; Ferrer-Montiel, A. Trafficking of thermo-TRP channels. Membranes 2014, 4, 525–564. [Google Scholar] [CrossRef] [PubMed]
- Mickle, A.D.; Shepherd, A.J.; Loo, L.; Mohapatra, D.P. Induction of thermal and mechanical hypersensitivity by parathyroid hormone-related peptide through upregulation of TRPV1 function and trafficking. Pain 2015, 156, 1620–1636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, J.; McNaughton, P.A. NGF rapidly increases membrane expression of TRPV1 heat-gated ion channels. EMBO J. 2005, 24, 4211–4223. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Dubin, A.E.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Nociceptive signals induce trafficking of TRPA1 to the plasma membrane. Neuron 2009, 64, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Wang, J.; Steinhoff, M.; Dolly, J.O. TNFalpha induces co-trafficking of TRPV1/TRPA1 in VAMP1-containing vesicles to the plasmalemma via Munc18-1/syntaxin1/SNAP-25 mediated fusion. Sci. Rep. 2016, 6, 21226. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Morsy, N.; Winston, J.; Pasricha, P.J.; Garrett, K.; Akbarali, H.I. Modulation of TRPV1 by nonreceptor tyrosine kinase, c-Src kinase. Am. J. Physiol. Cell Physiol. 2004, 287, C558–C563. [Google Scholar] [CrossRef] [PubMed]
- Bhave, G.; Hu, H.-J.; Glauner, K.S.; Zhu, W.; Wang, H.; Brasier, D.J.; Oxford, G.S.; Gereau, R.W. Protein kinase c phosphorylation sensitizes but does not activate the capsaicin receptor transient receptor potential vanilloid 1 (TRPV1). Proc. Natl. Acad. Sci. USA 2003, 100, 12480–12485. [Google Scholar] [CrossRef] [PubMed]
- Bhave, G.; Zhu, W.; Wang, H.; Brasier, D.J.; Oxford, G.S.; Gereau Iv, R.W. Camp-dependent protein kinase regulates desensitization of the capsaicin receptor (VR1) by direct phosphorylation. Neuron 2002, 35, 721–731. [Google Scholar] [CrossRef]
- Jung, J.; Shin, J.S.; Lee, S.Y.; Hwang, S.W.; Koo, J.; Cho, H.; Oh, U. Phosphorylation of vanilloid receptor 1 by Ca2+/calmodulin-dependent kinase II regulates its vanilloid binding. J. Biol. Chem. 2004, 279, 7048–7054. [Google Scholar] [CrossRef] [PubMed]
- Numazaki, M.; Tominaga, T.; Toyooka, H.; Tominaga, M. Direct phosphorylation of capsaicin receptor VR1 by protein kinase cepsilon and identification of two target serine residues. J. Biol. Chem. 2002, 277, 13375–13378. [Google Scholar] [CrossRef] [PubMed]
- Vellani, V.; Mapplebeck, S.; Moriondo, A.; Davis, J.B.; McNaughton, P.A. Protein kinase c activation potentiates gating of the vanilloid receptor VR1 by capsaicin, protons, heat and anandamide. J. Physiol. 2001, 534, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Pareek, T.K.; Keller, J.; Kesavapany, S.; Agarwal, N.; Kuner, R.; Pant, H.C.; Iadarola, M.J.; Brady, R.O.; Kulkarni, A.B. Cyclin-dependent kinase 5 modulates nociceptive signaling through direct phosphorylation of transient receptor potential vanilloid 1. Proc. Natl. Acad. Sci. USA 2007, 104, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, D.P.; Nau, C. Desensitization of capsaicin-activated currents in the vanilloid receptor TRPV1 is decreased by the cyclic AMP-dependent protein kinase pathway. J. Biol. Chem. 2003, 278, 50080–50090. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, D.P.; Nau, C. Regulation of Ca2+-dependent desensitization in the vanilloid receptor TRPV1 by calcineurin and camp-dependent protein kinase. J. Biol. Chem. 2005, 280, 13424–13432. [Google Scholar] [CrossRef] [PubMed]
- Spahn, V.; Stein, C.; Zollner, C. Modulation of transient receptor vanilloid 1 activity by transient receptor potential ankyrin 1. Mol. Pharmacol. 2014, 85, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Chuang, H.H.; Prescott, E.D.; Kong, H.; Shields, S.; Jordt, S.E.; Basbaum, A.I.; Chao, M.V.; Julius, D. Bradykinin and nerve growth factor release the capsaicin receptor from ptdins(4,5)p2-mediated inhibition. Nature 2001, 411, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Lukacs, V.; Yudin, Y.; Hammond, G.R.; Sharma, E.; Fukami, K.; Rohacs, T. Distinctive changes in plasma membrane phosphoinositides underlie differential regulation of TRPV1 in nociceptive neurons. J. Neurosci. 2013, 33, 11451–11463. [Google Scholar] [CrossRef] [PubMed]
- Hanack, C.; Moroni, M.; Lima, W.C.; Wende, H.; Kirchner, M.; Adelfinger, L.; Schrenk-Siemens, K.; Tappe-Theodor, A.; Wetzel, C.; Kuich, P.H.; et al. GABA blocks pathological but not acute TRPV1 pain signals. Cell 2015, 160, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Dai, Y.; Fukuoka, T.; Yamanaka, H.; Kobayashi, K.; Obata, K.; Cui, X.; Tominaga, M.; Noguchi, K. Phospholipase C and protein kinase A mediate bradykinin sensitization of TRPA1: A molecular mechanism of inflammatory pain. Brain 2008, 131, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Cavanaugh, E.J.; Simkin, D. Inhibition of transient receptor potential A1 channel by phosphatidylinositol-4,5-bisphosphate. Am. J. Physiol. Cell Physiol. 2008, 295, C92–C99. [Google Scholar] [CrossRef] [PubMed]
- Gregus, A.M.; Doolen, S.; Dumlao, D.S.; Buczynski, M.W.; Takasusuki, T.; Fitzsimmons, B.L.; Hua, X.-Y.; Taylor, B.K.; Dennis, E.A.; Yaksh, T.L. Spinal 12-lipoxygenase-derived hepoxilin a3 contributes to inflammatory hyperalgesia via activation of TRPV1 and TRPA1 receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 6721–6726. [Google Scholar] [CrossRef] [PubMed]
- Daniels, R.L.; Takashima, Y.; McKemy, D.D. Activity of the neuronal cold sensor TRPM8 is regulated by phospholipase C via the phospholipid phosphoinositol 4,5-bisphosphate. J. Biol. Chem. 2009, 284, 1570–1582. [Google Scholar] [CrossRef] [PubMed]
- Abe, J.; Hosokawa, H.; Sawada, Y.; Matsumura, K.; Kobayashi, S. Ca2+-dependent PKC activation mediates menthol-induced desensitization of transient receptor potential M8. Neurosci. Lett. 2006, 397, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, L.S.; Raisinghani, M.; Pingle, S.C.; Long, C.; Pimentel, F. Downregulation of transient receptor potential melastatin 8 by protein kinase C-mediated dephosphorylation. J. Neurosci. 2005, 25, 11322–11329. [Google Scholar] [CrossRef] [PubMed]
- Yudin, Y.; Lukacs, V.; Cao, C.; Rohacs, T. Decrease in phosphatidylinositol 4,5-bisphosphate levels mediates desensitization of the cold sensor TRPM8 channels. J. Physiol. 2011, 589, 6007–6027. [Google Scholar] [CrossRef] [PubMed]
- De Petrocellis, L.; Starowicz, K.; Moriello, A.S.; Vivese, M.; Orlando, P.; Di Marzo, V. Regulation of transient receptor potential channels of melastatin type 8 (TRPM8): Effect of cAMP, cannabinoid CB(1) receptors and endovanilloids. Exp. Cell Res. 2007, 313, 1911–1920. [Google Scholar] [CrossRef] [PubMed]
- Linte, R.M.; Ciobanu, C.; Reid, G.; Babes, A. Desensitization of cold- and menthol-sensitive rat dorsal root ganglion neurones by inflammatory mediators. Exp. Br. Res. 2007, 178, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Bavencoffe, A.; Gkika, D.; Kondratskyi, A.; Beck, B.; Borowiec, A.S.; Bidaux, G.; Busserolles, J.; Eschalier, A.; Shuba, Y.; Skryma, R.; et al. The transient receptor potential channel trpm8 is inhibited via the alpha 2a adrenoreceptor signaling pathway. J. Biol. Chem. 2010, 285, 9410–9419. [Google Scholar] [CrossRef] [PubMed]
- Sarria, I.; Gu, J. Menthol response and adaptation in nociceptive-like and nonnociceptive-like neurons: Role of protein kinases. Mol. Pain 2010, 6, 47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mak, S.; Li, L.; Parra, A.; Denlinger, B.; Belmonte, C.; McNaughton, P.A. Direct inhibition of the cold-activated TRPM8 ion channel by Galpha-q. Nat. Cell Biol. 2012, 14, 851–858. [Google Scholar] [CrossRef] [PubMed]
- De Petrocellis, L.; Vellani, V.; Schiano-Moriello, A.; Marini, P.; Magherini, P.C.; Orlando, P.; Di Marzo, V. Plant-derived cannabinoids modulate the activity of transient receptor potential channels of ankyrin type-1 and melastatin type-8. J. Pharmacol. Exp. Ther. 2008, 325, 1007–1015. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.A.; Nash, M.; Bevan, S. Modulation of the cold-activated channel TRPM8 by lysophospholipids and polyunsaturated fatty acids. J. Neurosci. 2007, 27, 3347–3355. [Google Scholar] [CrossRef] [PubMed]
- Vanden Abeele, F.; Zholos, A.; Bidaux, G.; Shuba, Y.; Thebault, S.; Beck, B.; Flourakis, M.; Panchin, Y.; Skryma, R.; Prevarskaya, N. Ca2+-independent phospholipase A2-dependent gating of TRPM8 by lysophospholipids. J. Biol. Chem. 2006, 281, 40174–40182. [Google Scholar] [CrossRef] [PubMed]
- Rohacs, T.; Lopes, C.M.; Michailidis, I.; Logothetis, D.E. Pi(4,5)p2 regulates the activation and desensitization of TRPM8 channels through the TRP domain. Nat. Neurosci. 2005, 8, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Qin, F. Functional control of cold- and menthol-sensitive TRPM8 ion channels by phosphatidylinositol 4,5-bisphosphate. J. Neurosci. 2005, 25, 1674–1681. [Google Scholar] [CrossRef] [PubMed]
- Rohacs, T. Regulation of transient receptor potential channels by the phospholipase C pathway. Adv. Biol. Regul. 2013, 53, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Stokes, A.J.; Shimoda, L.M.; Koblan-Huberson, M.; Adra, C.N.; Turner, H. A TRPV2-PKA signaling module for transduction of physical stimuli in mast cells. J. Exp. Med. 2004, 200, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Haber, N.; Dina, O.A.; Joseph, E.K.; Reichling, D.B.; Levine, J.D. Interaction of transient receptor potential vanilloid 4, integrin, and src tyrosine kinase in mechanical hyperalgesia. J. Neurosci. 2008, 28, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Denadai-Souza, A.; Martin, L.; de Paula, M.A.; de Avellar, M.C.; Muscara, M.N.; Vergnolle, N.; Cenac, N. Role of transient receptor potential vanilloid 4 in rat joint inflammation. Arthritis Rheum. 2012, 64, 1848–1858. [Google Scholar] [CrossRef] [PubMed]
- Wegierski, T.; Lewandrowski, U.; Muller, B.; Sickmann, A.; Walz, G. Tyrosine phosphorylation modulates the activity of TRPV4 in response to defined stimuli. J. Biol. Chem. 2009, 284, 2923–2933. [Google Scholar] [CrossRef] [PubMed]
- Melemedjian, O.K.; Tillu, D.V.; Moy, J.K.; Asiedu, M.N.; Mandell, E.K.; Ghosh, S.; Dussor, G.; Price, T.J. Local translation and retrograde axonal transport of CREB regulates IL-6-induced nociceptive plasticity. Mol. Pain 2014, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Melemedjian, O.K.; Asiedu, M.N.; Tillu, D.V.; Peebles, K.A.; Yan, J.; Ertz, N.; Dussor, G.O.; Price, T.J. IL-6- and NGF-induced rapid control of protein synthesis and nociceptive plasticity via convergent signaling to the eif4f complex. J. Neurosci. 2010, 30, 15113–15123. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Eskander, M.A.; Ruparel, S.; Green, D.P.; Chen, P.B.; Por, E.D.; Jeske, N.A.; Gao, X.; Flores, E.R.; Hargreaves, K.M. Persistent nociception triggered by nerve growth factor (NGF) is mediated by TRPV1 and oxidative mechanisms. J. Neurosci. 2015, 35, 8593–8603. [Google Scholar] [CrossRef] [PubMed]
- Hillery, C.A.; Kerstein, P.C.; Vilceanu, D.; Barabas, M.E.; Retherford, D.; Brandow, A.M.; Wandersee, N.J.; Stucky, C.L. Transient receptor potential vanilloid 1 mediates pain in mice with severe sickle cell disease. Blood 2011, 118, 3376–3383. [Google Scholar] [CrossRef] [PubMed]
- Ro, J.Y.; Lee, J.S.; Zhang, Y. Activation of TRPV1 and TRPA1 leads to muscle nociception and mechanical hyperalgesia. Pain 2009, 144, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Shutov, L.P.; Warwick, C.A.; Shi, X.; Gnanasekaran, A.; Shepherd, A.J.; Mohapatra, D.P.; Woodruff, T.M.; Clark, J.D.; Usachev, Y.M. The complement system component C5a produces thermal hyperalgesia via macrophage-to-sensory neuron signaling that requires NGF and TRPV1. J. Neurosci. 2016, 36, 5055–5070. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.; Helyes, Z.; Sandor, K.; Bite, A.; Pinter, E.; Nemeth, J.; Banvolgyi, A.; Bolcskei, K.; Elekes, K.; Szolcsanyi, J. Role of transient receptor potential vanilloid 1 receptors in adjuvant-induced chronic arthritis: In vivo study using gene-deficient mice. J. Pharmacol. Exp. Ther. 2005, 314, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Walder, R.Y.; Radhakrishnan, R.; Loo, L.; Rasmussen, L.A.; Mohapatra, D.P.; Wilson, S.P.; Sluka, K.A. TRPV1 is important for mechanical and heat sensitivity in uninjured animals and development of heat hypersensitivity after muscle inflammation. Pain 2012, 153, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Ueda, T.; Shibata, Y.; Kumamoto, N.; Ugawa, S. The role of TRPV1 channels in carrageenan-induced mechanical hyperalgesia in mice. Neuroreport 2015, 26, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Eid, S.R.; Crown, E.D.; Moore, E.L.; Liang, H.A.; Choong, K.C.; Dima, S.; Henze, D.A.; Kane, S.A.; Urban, M.O. HC-030031, a TRPA1 selective antagonist, attenuates inflammatory- and neuropathy-induced mechanical hypersensitivity. Mol. Pain 2008, 4, 48. [Google Scholar] [CrossRef] [PubMed]
- Petrus, M.; Peier, A.M.; Bandell, M.; Hwang, S.W.; Huynh, T.; Olney, N.; Jegla, T.; Patapoutian, A. A role of TRPA1 in mechanical hyperalgesia is revealed by pharmacological inhibition. Mol. Pain 2007, 3, 40. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, D.S.; Meotti, F.C.; Andrade, E.L.; Leal, P.C.; Motta, E.M.; Calixto, J.B. The involvement of the transient receptor potential a1 (TRPA1) in the maintenance of mechanical and cold hyperalgesia in persistent inflammation. Pain 2010, 148, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Katsura, H.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Tominaga, M.; Noguchi, K. TRPA1 induced in sensory neurons contributes to cold hyperalgesia after inflammation and nerve injury. J. Clin. Investig. 2005, 115, 2393–2401. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Escalera, J.; Balakrishna, S.; Fan, L.; Caceres, A.I.; Robinson, E.; Sui, A.; McKay, M.C.; McAlexander, M.A.; Herrick, C.A.; et al. TRPA1 controls inflammation and pruritogen responses in allergic contact dermatitis. FASEB J. 2013, 27, 3549–3563. [Google Scholar] [CrossRef] [PubMed]
- Than, J.Y.-X.L.; Li, L.; Hasan, R.; Zhang, X. Excitation and modulation of TRPA1, TRPV1, and TRPM8 channel-expressing sensory neurons by the pruritogen chloroquine. J. Biol. Chem. 2013, 288, 12818–12827. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; Nelson, A.M.; Batia, L.; Morita, T.; Estandian, D.; Owens, D.M.; Lumpkin, E.A.; Bautista, D.M. The ion channel TRPA1 is required for chronic itch. J. Neurosci. 2013, 33, 9283–9294. [Google Scholar] [CrossRef] [PubMed]
- Todaka, H.; Taniguchi, J.; Satoh, J.; Mizuno, A.; Suzuki, M. Warm temperature-sensitive transient receptor potential vanilloid 4 (TRPV4) plays an essential role in thermal hyperalgesia. J. Biol. Chem. 2004, 279, 35133–35138. [Google Scholar] [CrossRef] [PubMed]
- Barton, N.J.; McQueen, D.S.; Thomson, D.; Gauldie, S.D.; Wilson, A.W.; Salter, D.M.; Chessell, I.P. Attenuation of experimental arthritis in TRPV1r knockout mice. Exp. Mol. Pathol. 2006, 81, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.; Chapman, R.J.; Woodhams, S.; Sagar, D.R.; Turner, J.; Burston, J.J.; Bullock, C.; Paton, K.; Huang, J.; Wong, A.; et al. Increased function of pronociceptive TRPV1 at the level of the joint in a rat model of osteoarthritis pain. Ann. Rheum. Dis. 2013. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Russell, F.A.; Spina, D.; McDougall, J.J.; Graepel, R.; Gentry, C.; Staniland, A.A.; Mountford, D.M.; Keeble, J.E.; Malcangio, M.; et al. A distinct role for transient receptor potential ankyrin 1, in addition to transient receptor potential vanilloid 1, in tumor necrosis factor α–induced inflammatory hyperalgesia and freund’s complete adjuvant–induced monarthritis. Arthritis Rheum. 2011, 63, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; De Wilde, G.; Doherty, S.A.; Lories, R.J.; Vaughn, F.L.; Laslett, L.L.; Maciewicz, R.A.; Soni, A.; Hart, D.J.; Zhang, W.; et al. The ile585val trpv1 variant is involved in risk of painful knee osteoarthritis. Ann. Rheumatic Dis. 2011, 70, 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Treanor, J.J.; Garami, A.; Fang, L.; Surapaneni, S.; Akrami, A.; Alvarez, F.; Bak, A.; Darling, M.; Gore, A.; et al. Pharmacological blockade of the vanilloid receptor TRPV1 elicits marked hyperthermia in humans. Pain 2008, 136, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Kym, P.R.; Kort, M.E.; Hutchins, C.W. Analgesic potential of TRPV1 antagonists. Biochem. Pharmacol. 2009, 78, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Kort, M.E.; Kym, P.R. TRPV1 antagonists: Clinical setbacks and prospects for future development. Prog. Med. Chem. 2012, 51, 57–70. [Google Scholar] [PubMed]
- Wong, G.Y.; Gavva, N.R. Therapeutic potential of vanilloid receptor TRPV1 agonists and antagonists as analgesics: Recent advances and setbacks. Brain Res. Rev. 2009, 60, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Rowbotham, M.C.; Nothaft, W.; Duan, W.R.; Wang, Y.; Faltynek, C.; McGaraughty, S.; Chu, K.L.; Svensson, P. Oral and cutaneous thermosensory profile of selective TRPV1 inhibition by ABT-102 in a randomixed healthy volunteer trial. Pain 2011, 152, 1192–1200. [Google Scholar] [CrossRef] [PubMed]
- A Safety, Tolerability and Pharmacokinetic Study of ABT-102 in Healthy Subjects. Clinical Trials.gov 2010. Available online: https://clinicaltrials.gov/ct2/show/NCT00854659?term=ABT102&rank=1 (accessed on 28 October 2016).
- SB-705498 Dental Pain Study after Tooth Extraction. Clinical Trials.gov 2011. Available online: https://clinicaltrials.gov/ct2/show/NCT00281684?term=SB-705498&rank=8 (accessed on 28 October 2016).
- SB-705498 Rectal Pain Study. Clinical Trials.gov 2012. Available online: https://clinicaltrials.gov/ct2/show/NCT00461682?term=SB-705498&rank=10 (accessed on 28 October 2016).
- A Study to Evaluate the Safety and Efficacy of an Investigational Drug in the Treatment of Postoperative Dental Pain (MK-2295-005). Clinical Trials.gov 2015. Available online: https://clinicaltrials.gov/ct2/show/NCT00387140?term=MK2295&rank=1 (accessed on 28 October 2016).
- Esophageal Hypersensitivity Study in Healthy Volunteers. Clinical Trials.gov 2009. Available online: https://clinicaltrials.gov/ct2/show/NCT00711048?term=AZD1386&rank=11 (accessed on 28 October 2016).
- Krarup, A.L.; Ny, L.; Astrand, M.; Bajor, A.; Hvid-Jensen, F.; Hansen, M.B.; Simren, M.; Funch-Jensen, P.; Drewes, A.M. Randomised clinical trial: The efficacy of a transient receptor potnetial vanilloid 1 antagonist AZD1386 in human oesophageal pain. Aliment. Pharmacol. Ther. 2011, 33, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Study to Investigate the Analgesic Efficacy of a Single Dose of AZD1386. Clinical Trials.gov 2016. Available online: https://clinicaltrials.gov/ct2/show/NCT00672646?term=azd1386&rank=2 (accessed on 28 October 2016).
- Quiding, H.; Jonzon, B.; Svensson, O.; Webster, L.; Reimfelt, A.; Karin, A.; Karlsten, R.; Segerdahl, M. TRPV1 antagonistic analgesic effect: A randomized study of AZD1386 in pain after third molar extraction. Pain 2013, 154, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Study to Evaluate Efficacy, Safety, Tolerability and Pharmacokinetics of AZD1386 in Patients with Peripheral Neuropathic Pain (AVANT). Clinical Trials.gov 2009. Available online: https://clinicaltrials.gov/ct2/show/NCT00672646?term=azd1386&rank=2 (accessed on 28 October 2016).
- Study to Evaluate the Efficacy, Safety, Tolerability and Pharmacokinetics of AZD1386 in Patients with Osteoarthritis (OA) of the Knee (OA19). Clinical Trials.gov 2012. Available online: https://clinicaltrials.gov/ct2/show/NCT00672646?term=azd1386&rank=2 (accessed on 28 October 2016).
- NEO6860, a TRPV1 Antagonist, First in Human Study. Clinical Trials.gov 2016. Available online: https://clinicaltrials.gov/ct2/show/NCT02337543?term=TRPV1&rank=17 (accessed on 28 October 2016).
- A Proof-of-Concept Study Assessing NEO6860 in Osteoarthritis Pain. Clinical Trials.gov 2016. Available online: https://clinicaltrials.gov/ct2/show/NCT02712957?term=TRPV1&rank=20 (accessed on 28 October 2016).
- Study of NGX-4010 for the Treatment of Postherpetic Neuralgia. Clinical Trials.gov 2008. Available online: https://www.clinicaltrials.gov/ct2/show/NCT00115310?term=ngx+4010&rank=3 (accessed on 28 October 2016).
- Mou, J.; Paillard, F.; Trumbull, B.; Trudeau, J.; Stoker, M.; Katz, N.P. Qutenza (capsaicin) 8% patch onset and duration of response and effects of multiple treatments in neuropathic pain patients. Clin. J. Pain 2014, 30, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Study of NGX-4010 for the Treatment of Painful HIV-Associated Neuropathy. Clinical Trials.gov 2011. Available online: https://www.clinicaltrials.gov/ct2/show/results/NCT00321672?term=ngx+4010&rank=1 (accessed on 28 October 2016).
- Simpson, D.M.; Brown, S.; Tobias, J.K.; Vanhove, G.F.; NGX-4010 C107 Study Group. NGX-4010, a capsaicin 8% dermal patch, for the treatment of HIV-associated distal sensory polyneuropathy: Results of a 52-week open-label study. Clin. J. Pain 2014, 30, 134–142. [Google Scholar] [PubMed]
- Brown, D.C.; Agnello, K.; Iadarola, M.J. Intrathecal resiniferatoxin in a dog model: Efficacy in bone cancer pain. Pain 2015, 156, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Resiniferatoxin to Treat Severe Pain Associated with Advanced Cancer. Clinical Trials.gov 2016. Available online: https://clinicaltrials.gov/ct2/show/NCT00804154?term=Resiniferatoxin&rank=1 (accessed on 28 October 2016).
- Periganglionic Resiniferatoxin for the Treatment of Intractable Pain due to Cancer-Induced Bone Pain. Clinical Trials.gov 2016. Available online: https://clinicaltrials.gov/ct2/show/NCT02522611?term= Resiniferatoxin&rank=2 (accessed on 28 October 2016).
- Evaluating the Safety and Efficacy of Civamide (Zucapsaicin) in Osteoarthritis (OA) of the Knee(s). Clinical Trials.gov 2011. Available online: https://clinicaltrials.gov/ct2/show/NCT00995306?term=zucapsaicin&rank=2 (accessed on 28 October 2016).
- Evaluation of Civamide (Zucapsaicin) in Treatment of Postherpetic Neuroalgia and Post-Incisional Neuralgia. Clinical Trials.gov 2014. Available online: https://clinicaltrials.gov/ct2/show/NCT00845923? term=zucapsaicin&rank=4 (accessed on 28 October 2016).
- A phase III Study of Civamide Nasal Solution (Zucapsaicin) for the Treatment of Episodic Cluster Headache. Clinical Trials.gov 2011. Available online: https://clinicaltrials.gov/ct2/show/NCT00033839? term=zucapsaicin&rank=8 (accessed on 28 October 2016).
- A clinical trial to study the effects of GRC 17536 in patients with painful diabetic peripheral neuropathy (painful extremities due to peripheral nerve damage in diabetic patients). Clinical Trials.gov 2014. Available online: https://clinicaltrials.gov/ct2/show/NCT01726413?term=GRC-17536&rank=2 (accessed on 28 October 2016).
- Efficacy and Safety of SAR292833 Administration for 4 Weeks in Patients with Chronic Peripheral Neuropathic Pain (Alchemilla). Clinical Trials.gov 2016. Available online: https://clinicaltrials.gov/ct2/show/NCT01463397?term=NCT01463397&rank=1 (accessed on 28 October 2016).
- Winchester, W.J.; Gore, K.; Glatt, S.; Petit, W.; Gardiner, J.C.; Conlon, K.; Postlethwaite, M.; Saintot, P.P.; Roberts, S.; Gosset, J.R.; et al. Inhibition of TRPM8 channels reduces pain in the cold pressor test in humans. J. Pharmacol. Exp. Ther. 2014, 351, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Single dose, Dose Escalation Healthy Volunteers Study of PF-05105679 (Single Dose). Clinical Trials.gov 2011. Available online: https://clinicaltrials.gov/ct2/show/NCT01393652?term=PF-05105679&rank=1 (accessed on 28 October 2016).
- Kaneko, Y.; Szallasi, A. Transient receptor potnetial (TRP) channels: A clinical perspective. Br. J. Pharmacol. 2014, 171, 2474–2507. [Google Scholar] [CrossRef] [PubMed]
- Reilly, R.M.; McDonald, H.A.; Puttfarcken, P.S.; Joshi, S.K.; Lewis, L.; Pai, M.; Franklin, P.H.; Segreti, J.A.; Neelands, T.R.; Han, P.; et al. Pharmacology of modality-specific transient receptor potential vanilloid-1 antagonists that do not alter body temperature. J. Pharmacol. Exp. Ther. 2012, 342, 416–428. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Hwang, S. Emerging roles of TRPA1 in sensation of oxidative stress and its implications in defense and danger. Arch. Pharm. Res. 2013, 36, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Old, E.A.; Nadkarni, S.; Grist, J.; Gentry, C.; Bevan, S.; Kim, K.-W.; Mogg, A.J.; Perretti, M.; Malcangio, M. Monocytes expressing CX3CR1 orchestrate the development of vincristine-induced pain. J. Clin. Investig. 2014, 124, 2023–2036. [Google Scholar] [CrossRef] [PubMed]
- Barrière, D.A.; Rieusset, J.; Chanteranne, D.; Busserolles, J.; Chauvin, M.-A.; Chapuis, L.; Salles, J.; Dubray, C.; Morio, B. Paclitaxel therapy potentiates cold hyperalgesia in streptozotocin-induced diabetic rats through enhanced mitochondrial reactive oxygen species production and TRPA1 sensitization. Pain 2012, 153, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Nassini, R.; Gees, M.; Harrison, S.; De Siena, G.; Materazzi, S.; Moretto, N.; Failli, P.; Preti, D.; Marchetti, N.; Cavazzini, A.; et al. Oxaliplatin elicits mechanical and cold allodynia in rodents via TRPA1 receptor stimulation. Pain 2011, 152, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Katsura, H.; Obata, K.; Mizushima, T.; Yamanaka, H.; Kobayashi, K.; Dai, Y.; Fukuoka, T.; Tokunaga, A.; Sakagami, M.; Noguchi, K. Antisense knock down of TRPA1, but not TRPM8, alleviates cold hyperalgesia after spinal nerve ligation in rats. Exp. Neurol. 2006, 200, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Peña, C.; Viana, F. Targeting TRPM8 for pain relief. Open Pain J. 2013, 6, 154–164. [Google Scholar] [CrossRef]
- Colburn, R.W.; Lubin, M.L.; Stone, D.J., Jr.; Wang, Y.; Lawrence, D.; D’Andrea, M.R.; Brandt, M.R.; Liu, Y.; Flores, C.M.; Qin, N. Attenuated cold sensitivity in TRPM8 null mice. Neuron 2007, 54, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Rong, W.; Hillsley, K.; Davis, J.B.; Hicks, G.; Winchester, W.J.; Grundy, D. Jejunal afferent nerve sensitivity in wild-type and TRPV11 knockout mice. J. Physiol. 2004, 560, 867–881. [Google Scholar] [CrossRef] [PubMed]
- Kimball, E.S.; Wallace, N.H.; Schneider, C.R.; D’Andrea, M.R.; Hornby, P.J. Vanilloid receptor 1 antagonists attenuate disease severity in dextran sulphate sodium-induced colitis in mice. Neurogastroenterol. Motil. 2004, 16, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.L.; Facer, P.; Davis, J.B.; Smith, G.D.; Egerton, J.; Bountra, C.; Williams, N.S.; Anand, P. Sensory fibres expressing capsaicin receptor TRPV1 in patients with rectal hypersensitivity and faecal urgency. Lancet 2003, 361, 385–391. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Wang, P.; Merriam, F.V.; Bjorling, D.E. Lack of TRPV1 inhibits cystitis-induced increased mechanical sensitivity in mice. Pain 2008, 139, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.C., 3rd; Xu, L.; Gebhart, G.F. The mechanosensitivity of mouse colon afferent fibers and their sensitization by inflammatory mediators require transient receptor potential vanilloid 1 and acid-sensing ion channel 3. J. Neurosci. 2005, 25, 10981–10989. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.C., 3rd; Otsuka, E.; Wagstrom, E.; Jensen, C.S.; Price, M.P.; Gebhart, G.F. Short-term sensitization of colon mechanoreceptors is associated with long-term hypersensitivity to colon distention in the mouse. Gastroenterology 2007, 133, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Massa, F.; Sibaev, A.; Marsicano, G.; Blaudzun, H.; Storr, M.; Lutz, B. Vanilloid receptor (TRPV1)-deficient mice show increased susceptibility to dinitrobenzene sulfonic acid induced colitis. J. Mol. Med. 2006, 84, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Storr, M. TRPV1 in colitis: Is it a good or a bad receptor?—A viewpoint. Neurogastroenterol. Motil. 2007, 19, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Dang, K.; Bielefeldt, K.; Gebhart, G.F. Differential responses of bladder lumbosacral and thoracolumbar dorsal root ganglion neurons to purinergic agonists, protons, and capsaicin. J. Neurosci. 2005, 25, 3973–3984. [Google Scholar] [CrossRef] [PubMed]
- Cruz, F.; Guimaraes, M.; Silva, C.; Reis, M. Suppression of bladder hyperreflexia by intravesical resiniferatoxin. Lancet 1997, 350, 640–641. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Sikand, P. TRPV1: A target for next generation analgesics. Curr. Neuropharmacol. 2008, 6, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Dinis, P.; Charrua, A.; Avelino, A.; Cruz, F. Intravesical resiniferatoxin decreases spinal c-fos expression and increases bladder volume to reflex micturition in rats with chronic inflamed urinary bladders. BJU Int. 2004, 94, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Ceppa, E.; Cattaruzza, F.; Lyo, V.; Amadesi, S.; Pelayo, J.C.; Poole, D.P.; Vaksman, N.; Liedtke, W.; Cohen, D.M.; Grady, E.F.; et al. Transient receptor potential ion channels V4 and A1 contribute to pancreatitis pain in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G556–G571. [Google Scholar] [CrossRef] [PubMed]
- D’Aldebert, E.; Cenac, N.; Rousset, P.; Martin, L.; Rolland, C.; Chapman, K.; Selves, J.; Alric, L.; Vinel, J.P.; Vergnolle, N. Transient receptor potential vanilloid 4 activated inflammatory signals by intestinal epithelial cells and colitis in mice. Gastroenterology 2011, 140, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Une, Y.; Miyano, K.; Abe, H.; Hisaoka, K.; Morioka, N.; Nakata, Y. Activation of transient receptor potential ankyrin 1 evokes nociception through substance p release from primary sensory neurons. J. Neurochem. 2012, 120, 1036–1047. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Zuo, X.; Zhen, Y.; Yu, Y.; Gao, L. Transient receptor potential ankyrin-1 participates in visceral hyperalgesia following experimental colitis. Neurosci. Lett. 2008, 440, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Terada, Y.; Fujimura, M.; Nishimura, S.; Tsubota, M.; Sekiguchi, F.; Nishikawa, H.; Kawabata, A. Contribution of TRPA1 as a downstream signal of proteinase-activated receptor-2 to pancreatic pain. J. Pharmacol. Sci. 2013, 123, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, V.; Alpizar, Y.A.; Luis, E.; Tajada, S.; Denlinger, B.; Fajardo, O.; Manenschijn, J.-A.; Fernández-Peña, C.; Talavera, A.; Kichko, T.; et al. TRPA1 channels mediate acute neurogenic inflammation and pain produced by bacterial endotoxins. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Bessac, B.F.; Sivula, M.; von Hehn, C.A.; Escalera, J.; Cohn, L.; Jordt, S.-E. TRPA1 is a major oxidant sensor in murine airway sensory neurons. J. Clin. Investig. 2008, 118, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.A.; Gentry, C.; Alenmyr, L.; Killander, D.; Lewis, S.E.; Andersson, A.; Bucher, B.; Galzi, J.-L.; Sterner, O.; Bevan, S.; et al. TRPA1 mediates spinal antinociception induced by acetaminophen and the cannabinoid δ9-tetrahydrocannabiorcol. Nat. Commun. 2011, 2, 551. [Google Scholar] [CrossRef] [PubMed]
- Van Zanten, S.V. Review: Fibre, antispasmodics, and peppermint oil are all effective for irritable bowel syndrome. Evid.-Based Med. 2009, 14, 84. [Google Scholar] [CrossRef] [PubMed]
- Lashinger, E.S.; Steiginga, M.S.; Hieble, J.P.; Leon, L.A.; Gardner, S.D.; Nagilla, R.; Davenport, E.A.; Hoffman, B.E.; Laping, N.J.; Su, X. AMTB, a TRPM8 channel blocker: Evidence in rats for activity in overactive bladder and painful bladder syndrome. Am. J. Physiol. Renal Physiol. 2008, 295, F803–F810. [Google Scholar] [CrossRef] [PubMed]
- Mantyh, P. Bone cancer pain: Causes, consequences, and therapeutic opportunities. Pain 2013, 154, S54–S62. [Google Scholar] [CrossRef] [PubMed]
- Ghilardi, J.R.; Rohrich, H.; Lindsay, T.H.; Sevcik, M.A.; Schwei, M.J.; Kubota, K.; Halvorson, K.G.; Poblete, J.; Chaplan, S.R.; Dubin, A.E.; et al. Selective blockade of the capsaicin receptor TRPV1 attenuates bone cancer pain. J. Neurosci. 2005, 25, 3126–3131. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Posadas, A.; Picazo-Juarez, G.; Llorente, I.; Jara-Oseguera, A.; Morales-Lazaro, S.; Escalante-Alcalde, D.; Islas, L.D.; Rosenbaum, T. Lysophosphatidic acid directly activates TRPV1 through a c-terminal binding site. Nat. Chem. Biol. 2012, 8, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.L.; Zhang, Y.Q.; Zhao, Z.Q. Involvement of lysophosphatidic acid in bone cancer pain by potentiation of TRPV1 via PKC-epsilon pathway in dorsal root ganglion neurons. Mol. Pain 2010, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Pan, H.L.; Li, T.T.; Zhang, Y.Q.; Wei, J.Y.; Zhao, Z.Q. The sensitization of peripheral c-fibers to lysophosphatidic acid in bone cancer pain. Life Sci. 2010, 87, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.C.; Iadarola, M.J.; Perkowski, S.Z.; Erin, H.; Shofer, F.; Laszlo, K.J.; Olah, Z.; Mannes, A.J. Physiologic and antinociceptive effects of intrathecal resiniferatoxin in a canine bone cancer model. Anesthesiology 2005, 103, 1052–1059. [Google Scholar] [CrossRef] [PubMed]
- Park, C.K.; Kim, M.S.; Fang, Z.; Li, H.Y.; Jung, S.J.; Choi, S.Y.; Lee, S.J.; Park, K.; Kim, J.S.; Oh, S.B. Functional expression of thermo-transient receptor potential channels in dental primary afferent neurons: Implication for tooth pain. J. Biol. Chem. 2006, 281, 17304–17311. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Chung, G.; Jo, H.J.; Kim, Y.S.; Bae, Y.C.; Jung, S.J.; Kim, J.S.; Oh, S.B. Characterization of dental nociceptive neurons. J. Dent. Res. 2011, 90, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Chung, G.; Oh, S.B. TRP channels in dental pain. Open Pain J. 2013, 6, 31–36. [Google Scholar] [CrossRef]
- Chung, M.K.; Lee, J.; Duraes, G.; Ro, J.Y. Lipopolysaccharide-induced pulpitis up-regulates TRPV1 in trigeminal ganglia. J. Dent. Res. 2011, 90, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Haas, E.T.; Rowland, K.; Gautam, M. Tooth injury increases expression of the cold sensitive trp channel TRPA1 in trigeminal neurons. Arch. Oral Biol. 2011, 56, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Meents, J.E.; Hoffmann, J.; Chaplan, S.R.; Neeb, L.; Schuh-Hofer, S.; Wickenden, A.; Reuter, U. Two TRPV1 receptor antagonists are effective in two different experimental models of migraine. J. Headache Pain 2015, 16, 57. [Google Scholar] [CrossRef] [PubMed]
- Chasman, D.I.; Schurks, M.; Anttila, V.; de Vries, B.; Schminke, U.; Launer, L.J.; Terwindt, G.M.; van den Maagdenberg, A.M.; Fendrich, K.; Volzke, H.; et al. Genome-wide association study reveals three susceptibility loci for common migraine in the general population. Nat. Genet. 2011, 43, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Edelmayer, R.M.; Le, L.N.; Yan, J.; Wei, X.; Nassini, R.; Materazzi, S.; Preti, D.; Appendino, G.; Geppetti, P.; Dodick, D.W.; et al. Activation of TRPA1 on dural afferents: A potential mechanism of headache pain. Pain 2012, 153, 1949–1958. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, G.; Benemei, S.; Materazzi, S.; De Logu, F.; De Siena, G.; Fusi, C.; Fortes Rossato, M.; Coppi, E.; Marone, I.M.; Ferreira, J.; et al. TRPA1 mediates trigeminal neuropathic pain in mice downstream of monocytes/macrophages and oxidative stress. Brain 2016, 139, 1361–1377. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, D.J.; Lee, H.; Lo, L.; Shields, S.D.; Zylka, M.J.; Basbaum, A.I.; Anderson, D.J. Distinct subsets of unmyelinated primary sensory fibers mediate behavioral responses to noxious thermal and mechanical stimuli. Proc. Natl. Acad. Sci. USA 2009, 106, 9075–9080. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.G.; Zhuo, M. No requirement of TRPV1 in long-term potentiation or long-term depression in the anterior cingulate cortex. Mol. Brain 2014, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Geppetti, P.; Materazzi, S.; Nicoletti, P. The transient receptor potential vanilloid 1: Role in airway inflammation and disease. Eur. J. Pharmacol. 2006, 533, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; McLeod, R.L.; Hey, J.A. TRPV1 receptor: A target for the treatment of pain, cough, airway disease and urinary incontinence. Drug News Perspect. 2005, 18, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Gunthorpe, M.J.; Szallasi, A. Peripheral TRPV1 receptors as targets for drug development: New molecules and mechanisms. Curr. Pharm. Des. 2008, 14, 32–41. [Google Scholar] [PubMed]
- Bhattacharya, A.; Scott, B.P.; Nasser, N.; Ao, H.; Maher, M.P.; Dubin, A.E.; Swanson, D.M.; Shankley, N.P.; Wickenden, A.D.; Chaplan, S.R. Pharmacology and antitussive efficacy of 4-(3-trifluoromethyl-pyridin-2-yl)-piperazine-1-carboxylic acid (5-trifluoromethyl-pyridin-2-yl)-amide (jnj17203212), a transient receptor potential vanilloid 1 antagonist in guinea pigs. J. Pharmacol. Exp. Ther. 2007, 323, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Razavi, R.; Chan, Y.; Afifiyan, F.N.; Liu, X.J.; Wan, X.; Yantha, J.; Tsui, H.; Tang, L.; Tsai, S.; Santamaria, P.; et al. TRPV1+ sensory neurons control beta cell stress and islet inflammation in autoimmune diabetes. Cell 2006, 127, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Park, U.; Vastani, N.; Guan, Y.; Raja, S.N.; Koltzenburg, M.; Caterina, M.J. TRP vanilloid 2 knock-out mice are susceptible to perinatal lethality but display normal thermal and mechanical nociception. J. Neurosci. 2011, 31, 11425–11436. [Google Scholar] [CrossRef] [PubMed]
- Link, T.M.; Park, U.; Vonakis, B.M.; Raben, D.M.; Soloski, M.J.; Caterina, M.J. TRPV2 has a pivotal role in macrophage particle binding and phagocytosis. Nat. Immunol. 2010, 11, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Dhaka, A.; Murray, A.N.; Mathur, J.; Earley, T.J.; Petrus, M.J.; Patapoutian, A. TRPM8 is required for cold sensation in mice. Neuron 2007, 54, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D.M.; Siemens, J.; Glazer, J.M.; Tsuruda, P.R.; Basbaum, A.I.; Stucky, C.L.; Jordt, S.E.; Julius, D. The menthol receptor TRPM8 is the principal detector of environmental cold. Nature 2007, 448, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Noel, J.; Zimmermann, K.; Busserolles, J.; Deval, E.; Alloui, A.; Diochot, S.; Guy, N.; Borsotto, M.; Reeh, P.; Eschalier, A.; et al. The mechano-activated K+ channels TRAAK and TREK-1 control both warm and cold perception. EMBO J. 2009, 28, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Corey, D.P.; Garcia-Anoveros, J.; Holt, J.R.; Kwan, K.Y.; Lin, S.Y.; Vollrath, M.A.; Amalfitano, A.; Cheung, E.L.; Derfler, B.H.; Duggan, A.; et al. TRPA1 is a candidate for the mechanosensitive transduction channel of vertebrate hair cells. Nature 2004, 432, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Nagata, K.; Duggan, A.; Kumar, G.; Garcia-Anoveros, J. Nociceptor and hair cell transducer properties of TRPA1, a channel for pain and hearing. J. Neurosci. 2005, 25, 4052–4061. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Allchorne, A.J.; Vollrath, M.A.; Christensen, A.P.; Zhang, D.S.; Woolf, C.J.; Corey, D.P. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron 2006, 50, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, M.Z.P.; Jordt, S.E. TRPA1: A sensory channel of many talents. In TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades; Liedtke, W.B., Heller, S., Eds.; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Zygmunt, P.M.; Hogestatt, E.D. TRPA1. In Handbook of Experimental Pharmacology; Springer: Berlin, Germany, 2014; Volume 222, pp. 583–630. [Google Scholar]


{kind=link}
| Agonist/Antagonist (Producer/Company) | Channel (Mode of Action) | Current Clinical Use/Trial Status | Specific Painful Pathology | References |
|---|---|---|---|---|
| AMG517 (Amgen) | TRPV1 (channel blocker) | Phase Ib/Phase II-terminated | Dental pain | [147,148,149,150] |
| ABT102 (Abbott) | TRPV1 (channel blocker) | Phase I—completed; Phase II—unknown | Healthy volunteers | [148,149,150,151,152] |
| GRC 6211 (Lilly/Glenmark) | TRPV1 (channel blocker) | Phase I-Phase II | Dental pain | [148,149,150] |
| SB-705498 (GlaxoSmithKline) | TRPV1 (channel blocker) | Phase II—completed | Dental pain/Toothache | [149,150,153] |
| Phase II—terminated | Rectal pain | [154] | ||
| MK-2295 (Merck-Neurogen) | TRPV1 (channel blocker) | Phase II—completed | Postoperative dental pain | [149,150,155] |
| AZD1386 (Astra-Zeneca) | TRPV1 (channel blocker) | Phase I—completed | Esophageal pain | [149,150,156,157] |
| Phase II—completed | Dental pain | [149,158,159] | ||
| Phase II—terminated | Osteoarthritis, knee pain | [149,150,160] | ||
| Phase II—terminated | Neuropathic pain | [161] | ||
| NEO06860 (Neomed Institute) | TRPV1 (blockade of channel activation by capsaicin, but not by heat and protons) | Phase I—completed Phase II—ongoing | Osteoarthritis, knee pain | [162,163] |
| NGX 4010 (NeurogesX; Acorda Therapeutics/ Astellas Pharma) | TRPV1 (agonist; capsaicin transdermal patch) | Phase III—completed (launched for clinical use in PHN) | PHN-associated neuropathic pain | [164,165] |
| Phase III—completed | HIV-associated neuropathic pain | [165,166,167] | ||
| Resiniferatoxin (NIDCR-NIH) | TRPV1 (potent agonist) | Phase I—ongoing | Advanced cancer pain | [168,169,170] |
| Zucapsaicin (Sanofi-Aventis; Winston Pharmaceuticals) | TRPV1 (agonist; nasal and topical capsaicin patch & cream) | Phase III—completed (launched for clinical use in osteoarthritis) | Osteoarthritis, knee pain | [171] |
| Phase III—completed | Episodic cluster headache | [172] | ||
| Phase II—completed | PHN-associated neuropathic pain | [173] | ||
| GRC-17536 (Glenmark) | TRPA1 (channel locker) | Phase II—completed | Diabetic peripheral neuropathic pain | [174] |
| SAR292833 (Sanofi) | TRPV3 (channel blocker) | Phase II—completed | Neuropathic pain | [175] |
| PF-05PR105679 (Pfizer) | TRPM8 (channel blocker) | Phase I—completed | Healthy volunteers | [176,177] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Nociceptive TRP Channels: Sensory Detectors and Transducers in Multiple Pain Pathologies. Pharmaceuticals 2016, 9, 72. https://doi.org/10.3390/ph9040072
Mickle AD, Shepherd AJ, Mohapatra DP. Nociceptive TRP Channels: Sensory Detectors and Transducers in Multiple Pain Pathologies. Pharmaceuticals. 2016; 9(4):72. https://doi.org/10.3390/ph9040072
Chicago/Turabian StyleMickle, Aaron D., Andrew J. Shepherd, and Durga P. Mohapatra. 2016. "Nociceptive TRP Channels: Sensory Detectors and Transducers in Multiple Pain Pathologies" Pharmaceuticals 9, no. 4: 72. https://doi.org/10.3390/ph9040072
APA StyleMickle, A. D., Shepherd, A. J., & Mohapatra, D. P. (2016). Nociceptive TRP Channels: Sensory Detectors and Transducers in Multiple Pain Pathologies. Pharmaceuticals, 9(4), 72. https://doi.org/10.3390/ph9040072