
Hepatic Gene Expression Changes in Rats Internally Exposed to Radioactive 56MnO2 Particles at Low Doses

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Irradiation
2.3. Pathology
2.4. Measurement of mRNA Levels by Quantitative RT-PCR
2.5. Statistical Analysis
3. Results
3.1. Radiation Doses
3.2. Body and Liver Weights
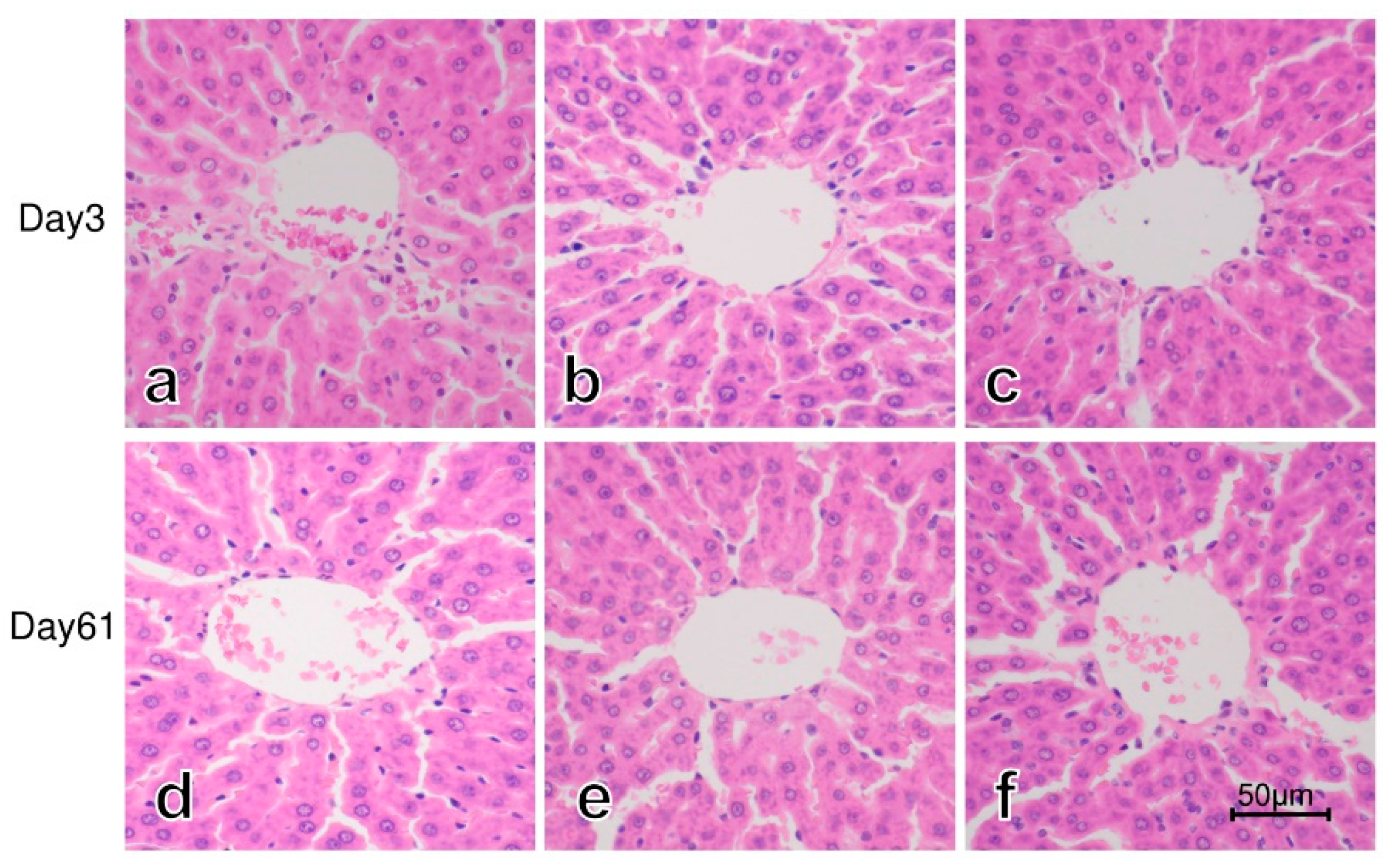
3.3. Histology of the Liver
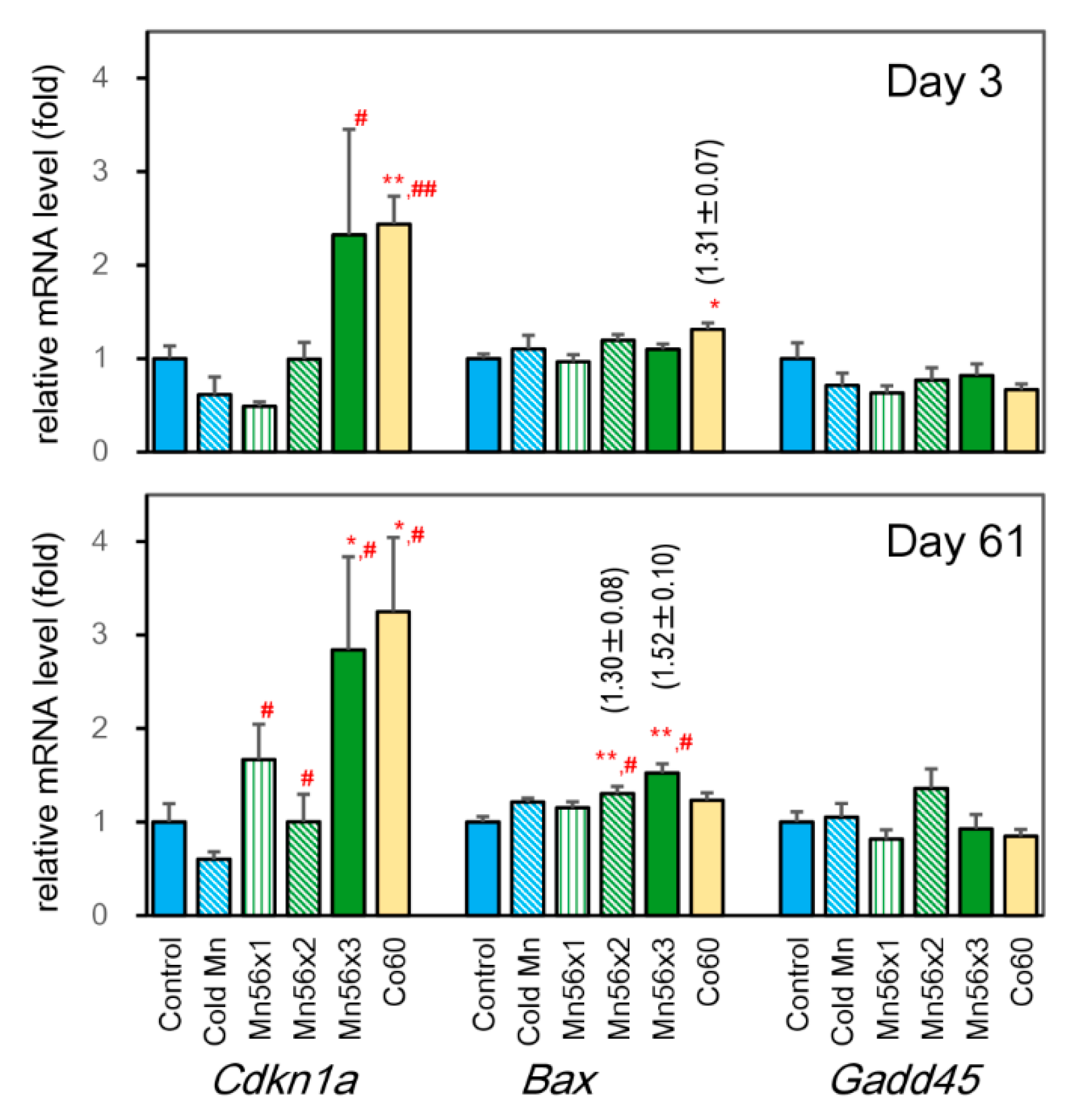
3.4. Effects on mRNA Expression Levels of Cdkn1a, BCL2 Associated X (Bax), and Growth-arrest-and-DNA-damage-inducible 45 (Gadd45)
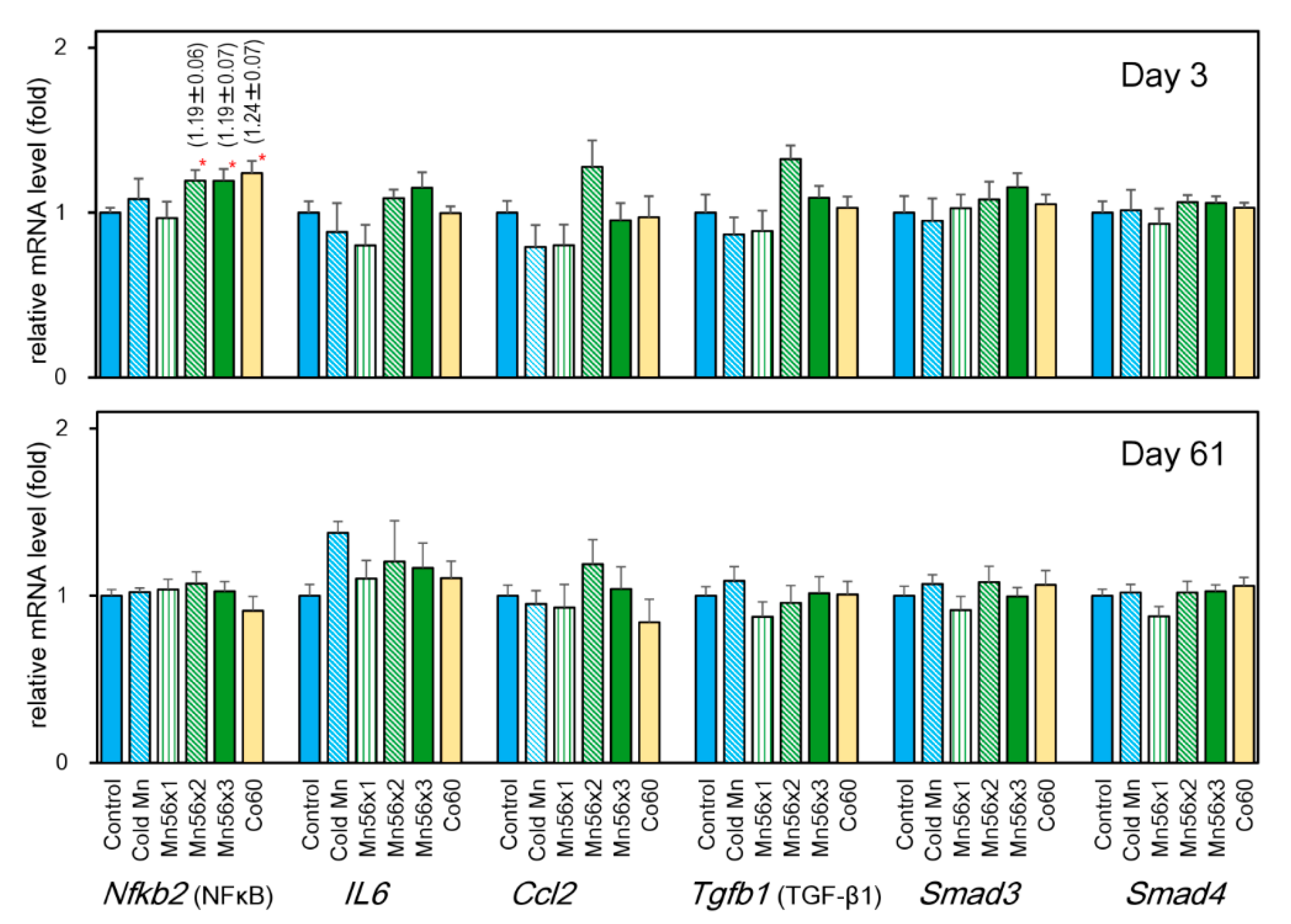
3.5. Effects on mRNA Levels of NFκB (Nfkb1), Interleukin (Il6), Chemokine Ligand 2 (Ccl2), TGF-β1 (Tgfb1), Smad3, and Smad4
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Imanaka, T.; Endo, S.; Tanaka, K.; Shizuma, K. Gamma-ray exposure from neutron-induced radionuclides in soil in Hiroshima and Nagasaki based on DS02 calculations. Radiat. Environ. Biophys. 2008, 47, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Endo, S.; Imanaka, T.; Shizuma, K.; Hasai, H.; Hoshi, M. Skin dose from neutron-activated soil for early entrants following the A-bomb detonation in Hiroshima: Contribution from β and γ rays. Radiat. Environ. Biophys. 2008, 47, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Stepanenko, V.; Kaprin, A.; Ivanov, S.; Shegay, P.; Zhumadilov, K.; Petukhov, A.; Kolyzhenkov, T.; Bogacheva, V.; Zharova, E.; Iaskova, E.; et al. Internal doses in experimental mice and rats following exposure to neutron-activated 56MnO2 powder: Results of an international, multicenter study. Radiat. Environ. Biophys. 2020, 59, 683–692. [Google Scholar] [CrossRef]
- Fujimoto, N.; Baurzhan, A.; Chaizhunusova, N.; Amantayeva, G.; Kairkhanova, Y.; Shabdarbaeva, D.; Zhunussov, Y.; Zhumadilov, K.; Stepanenko, V.; Gnyrya, V.; et al. Effects of internal exposure to 56MnO2 powder on blood parameters in rats. Eurasian J. Med. 2020, 52, 52–56. [Google Scholar] [CrossRef]
- Chopra, S.; Moroni, M.; Martello, S.; Bylicky, M.; May, J.; Hritzo, B.; MacMillan, L.; Coleman, C.N.; Aryankalayil, M.J. Gene expression profiles from heart, lung and liver samples of total-body-irradiated minipigs: Implications for predicting radiation-induced tissue toxicity. Radiat. Res. 2020, 194, 411–430. [Google Scholar] [CrossRef]
- Christiansen, H.; Batusic, D.; Saile, B.; Hermann, R.M.; Dudas, J.; Rave-Frank, M.; Hess, C.F.; Schmidberger, H.; Ramadori, G. Identification of genes responsive to gamma radiation in rat hepatocytes and rat liver by cDNA array gene expression analysis. Radiat. Res. 2006, 165, 318–325. [Google Scholar] [CrossRef]
- Roudkenar, M.H.; Li, L.; Baba, T.; Kuwahara, Y.; Nakagawa, H.; Wang, L.; Kasaoka, S.; Ohkubo, Y.; Ono, K.; Fukumoto, M. Gene expression profiles in mouse liver cells after exposure to different types of radiation. J. Radiat. Res. 2008, 49, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Jung, Y. Radiation-induced liver disease: Current understanding and future perspectives. Exp. Mol. Med. 2017, 49, e359. [Google Scholar] [CrossRef] [Green Version]
- Han, N.-K.; Jung, M.G.; Jeong, Y.J.; Son, Y.; Han, S.C.; Park, S.; Lim, Y.-B.; Lee, Y.-J.; Kim, S.-H.; Park, S.C.; et al. Plasma fibrinogen-like 1 as a potential biomarker for radiation-induced liver injury. Cells 2019, 8, 1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imaeda, M.; Ishikawa, H.; Yoshida, Y.; Takahashi, T.; Ohkubo, Y.; Musha, A.; Komachi, M.; Nakazato, Y.; Nakano, T. Long-term pathological and immunohistochemical features in the liver after intraoperative whole-liver irradiation in rats. J. Radiat. Res. 2014, 55, 665–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.; Xiao, L.; Ainiwaer, A.; Wang, Y.; Wu, G.; Mao, R.; Yang, Y.; Bao, Y. Molecular responses of radiation-induced liver damage in rats. Mol. Med. Rep. 2015, 11, 2592–2600. [Google Scholar] [CrossRef]
- Flanders, K.C. Smad3 as a mediator of the fibrotic response. Int. J. Exp. Pathol. 2004, 85, 47–64. [Google Scholar] [CrossRef]
- Seong, J.; Kim, S.H.; Chung, E.J.; Lee, W.J.; Suh, C.O. Early alteration in TGF-beta mRNA expression in irradiated rat liver. Int. J. Radiat. Oncol. Biol. Phys. 2000, 46, 639–643. [Google Scholar] [CrossRef]
- Ali, O.S.M.; Amin, N.E.D.; Abdel Fattah, S.M.; Abd El-Rahman, O. Ameliorative effect of kefir against γ-irradiation induced liver injury in male rats: Impact on oxidative stress and inflammation. Environ. Sci. Pollut. Res. 2020, 27, 35161–35173. [Google Scholar] [CrossRef]
- Eassawy, M.M.T.; Salem, A.A.; Ismail, A.F.M. Biochemical study on the protective effect of curcumin on acetaminophen and gamma-irradiation induced hepatic toxicity in rats. Environ. Toxicol. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Kruse, J.J.C.M.; Stewart, F. Gene expression arrays as a tool to unravel mechanisms of normal tissue radiation injury and prediction of response. World J. Gastroenterol. 2007, 13, 2669–2674. [Google Scholar] [CrossRef] [Green Version]
- Pawlik, A.; Delmar, P.; Bosse, S.; Sainz, L.; Petat, C.; Piétu, G.; Thierry, D.; Roux, D.T.-L. Changes in transcriptome after in vivo exposure to ionising radiation reveal a highly specialised liver response. Int. J. Radiat. Biol. 2009, 85, 656–671. [Google Scholar] [CrossRef] [PubMed]
- Stepanenko, V.; Rakhypbekov, T.; Otani, K.; Endo, S.; Satoh, K.; Kawano, N.; Shichijo, K.; Nakashima, M.; Takatsuji, T.; Sakaguchi, A.; et al. Internal exposure to neutron-activated 56Mn dioxide powder in Wistar rats: Part 1: Dosimetry. Radiat. Environ. Biophys. 2017, 56, 47–54. [Google Scholar] [CrossRef]
- Fujimoto, N.; Suzuki, T.; Ohta, S.; Kitamura, S. Identification of rat prostatic secreted proteins using mass spectrometric analysis and androgen-dependent mRNA expression. J. Androl. 2009, 30, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, N.; Amantayeva, G.; Chaizhunussova, N.; Shabdarbayeva, D.; Abishev, Z.; Ruslanova, B.; Zhunussov, Y.; Azhimkhanov, A.; Zhumadilov, K.; Petukhov, A.; et al. Low-dose radiation exposure with 56MnO2 powder changes gene expressions in the testes and the prostate in rats. Int. J. Mol. Sci. 2020, 21, 4989. [Google Scholar] [CrossRef] [PubMed]
- Straub, J.M.; New, J.; Hamilton, C.D.; Lominska, C.; Shnayder, Y.; Thomas, S.M. Radiation-induced fibrosis: Mechanisms and implications for therapy. J. Cancer Res. Clin. Oncol. 2015, 141, 1985–1994. [Google Scholar] [CrossRef] [Green Version]
- Alkhalf, M.I.; Khalifa, F.K. Blueberry extract attenuates gamma-radiation-induced hepatocyte damage by modulating oxidative stress and suppressing NF-kappaB in male rats. Saudi J. Biol. Sci. 2018, 25, 1272–1277. [Google Scholar] [CrossRef]
- El-Gazzar, M.G.; Zaher, N.H.; El-Hossary, E.M.; Ismail, A.F.M. Radio-protective effect of some new curcumin analogues. J. Photochem. Photobiol. B 2016, 162, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Moriconi, F.; Christiansen, H.; Raddatz, D.; Dudas, J.; Hermann, R.M.; Rave-Fränk, M.; Sheikh, N.; Saile, B.; Hess, C.F.; Ramadori, G. Effect of Radiation on Gene Expression of Rat Liver Chemokines: In Vivo and In Vitro Studies. Radiat. Res. 2008, 169, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Rodel, F.; Hantschel, M.; Hildebrandt, G.; Schultze-Mosgau, S.; Rodel, C.; Herrmann, M.; Sauer, R.; Voll, R.E. Dose-dependent biphasic induction and transcriptional activity of nuclear factor kappa B (NF-kappaB) in EA.hy.926 endothelial cells after low-dose X-irradiation. Int. J. Radiat. Biol. 2004, 80, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Song, L.-H.; Yan, H.-L.; Cai, D.-L. Gene expression profiles in the liver of mice irradiated with 60Co gamma rays and treated with soybean isoflavone. Eur. J. Nutr. 2006, 45, 406–417. [Google Scholar] [CrossRef] [PubMed]
- Roy, L.; Gruel, G.; Vaurijoux, A. Cell response to ionising radiation analysed by gene expression patterns. Ann. Dell’istituto Super. Sanità 2009, 45, 272–277. [Google Scholar]
- Taylor, W.R.; Stark, G.R. Regulation of the G2/M transition by p53. Oncogene 2001, 20, 1803–1815. [Google Scholar] [CrossRef] [Green Version]
- Daino, K.; Ichimura, S.; Nenoi, M. Early induction of CDKN1A (p21) and GADD45 mRNA by a low dose of ionizing radiation is due to their dose-dependent post-transcriptional regulation. Radiat. Res. 2002, 157, 478–482. [Google Scholar] [CrossRef]
- Gu, Z.; Flemington, C.; Chittenden, T.; Zambetti, G.P. ei24, a p53 Response Gene Involved in Growth Suppression and Apoptosis. Mol. Cell. Biol. 2000, 20, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Kabacik, S.; MacKay, A.; Tamber, N.; Manning, G.; Finnon, P.; Paillier, F.; Ashworth, A.; Bouffler, S.; Badie, C. Gene ex-pression following ionising radiation: Identification of biomarkers for dose estimation and prediction of individual response. Int. J. Radiat. Res. 2011, 87, 115–129. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession Number | Q-PCR Primer Sequences (5′ → 3′) | |
|---|---|---|---|
| Forward | Reverse | ||
| Cdkn1a | NM_080782 | TGTCCGACCTGTTCCACACA | CGTCTCAGTGGCGAAGTCAA |
| Bax | NM_017059 | TGTGGATACAGACTCCCCCC | TGATCAGCTCGGGCACTTTA |
| Gadd45 | L32591 | GAGTCAGCGCACCATAACTGTC | AATGAGGGTGAAATGGATCTGC |
| Nfkb1(NFκB) | LC369719 | CCTGTCTGAAGCCCTGCTACA | CACACCCTGGTTCAGAAGCTG |
| Il6 | NM_012589 | TCACAGAGGATACCACCCACAA | TCTGACAGTGCATCATCGCTG |
| Ccl2 | NM_031530 | AAGCCAGATCTCTCTTCCTCCA | CAGCAACTGTGAACAACAGGC |
| Tgfb1(TGFβ1) | AY550025 | GCTGAACCAAGGAGACGGAAT | GAAGGGTCGGTTCATGTCATG |
| Smad3 | NM_013095 | AGAACGTGAACACCAAGTGCAT | CCCGTAACTCATGGTGGCTG |
| Smad4 | AB010954 | ACGGAAGGACATTCGATTCAA | GACTTGTGGAAGCCACAGGAA |
| Groups 1 | Body Weight (g) | Liver (g) | Liver (Relative) (g/kg bw) | |
|---|---|---|---|---|
| Day3 | Control | 248 ± 16 | 6.80 ± 0.40 | 28 ± 1.2 |
| Cold Mn | 235 ± 14 | 7.08 ± 0.41 | 30 ± 0.4 | |
| Mn56x1 | 235 ± 11 | 6.68 ± 0.40 | 28 ± 1.0 | |
| Mn56x2 | 245 ± 16 | 6.86 ± 0.39 | 28 ± 0.8 | |
| Mn56x3 | 237 ± 12 | 6.94 ± 0.37 | 29 ± 0.5 | |
| Co60 | 234 ± 14 | 6.71 ± 0.34 | 29 ± 1.5 | |
| Day61 | Control | 330 ± 17 | 8.73 ± 0.54 | 26 ± 0.7 |
| Cold Mn | 337 ± 19 | 10.8 ± 0.51 | 32 ± 1.0 | |
| Mn56x1 | 371 ± 21 | 13.2 ± 0.71 ** | 36 ± 1.4 ** | |
| Mn56x2 | 337 ± 17 | 8.94 ± 0.54 | 27 ± 1.0 | |
| Mn56x3 | 353 ± 17 | 9.51 ± 0.40 | 27 ± 0.4 | |
| Co60 | 328 ± 23 | 8.67 ± 0.70 | 26 ± 0.8 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruslanova, B.; Abishev, Z.; Chaizhunussova, N.; Shabdarbayeva, D.; Tokesheva, S.; Amantayeva, G.; Kairkhanova, Y.; Stepanenko, V.; Hoshi, M.; Fujimoto, N. Hepatic Gene Expression Changes in Rats Internally Exposed to Radioactive 56MnO2 Particles at Low Doses. Curr. Issues Mol. Biol. 2021, 43, 758-766. https://doi.org/10.3390/cimb43020055
Ruslanova B, Abishev Z, Chaizhunussova N, Shabdarbayeva D, Tokesheva S, Amantayeva G, Kairkhanova Y, Stepanenko V, Hoshi M, Fujimoto N. Hepatic Gene Expression Changes in Rats Internally Exposed to Radioactive 56MnO2 Particles at Low Doses. Current Issues in Molecular Biology. 2021; 43(2):758-766. https://doi.org/10.3390/cimb43020055
Chicago/Turabian StyleRuslanova, Bakhyt, Zhaslan Abishev, Nailya Chaizhunussova, Dariya Shabdarbayeva, Sholpan Tokesheva, Gaukhar Amantayeva, Ynkar Kairkhanova, Valeriy Stepanenko, Masaharu Hoshi, and Nariaki Fujimoto. 2021. "Hepatic Gene Expression Changes in Rats Internally Exposed to Radioactive 56MnO2 Particles at Low Doses" Current Issues in Molecular Biology 43, no. 2: 758-766. https://doi.org/10.3390/cimb43020055
APA StyleRuslanova, B., Abishev, Z., Chaizhunussova, N., Shabdarbayeva, D., Tokesheva, S., Amantayeva, G., Kairkhanova, Y., Stepanenko, V., Hoshi, M., & Fujimoto, N. (2021). Hepatic Gene Expression Changes in Rats Internally Exposed to Radioactive 56MnO2 Particles at Low Doses. Current Issues in Molecular Biology, 43(2), 758-766. https://doi.org/10.3390/cimb43020055