
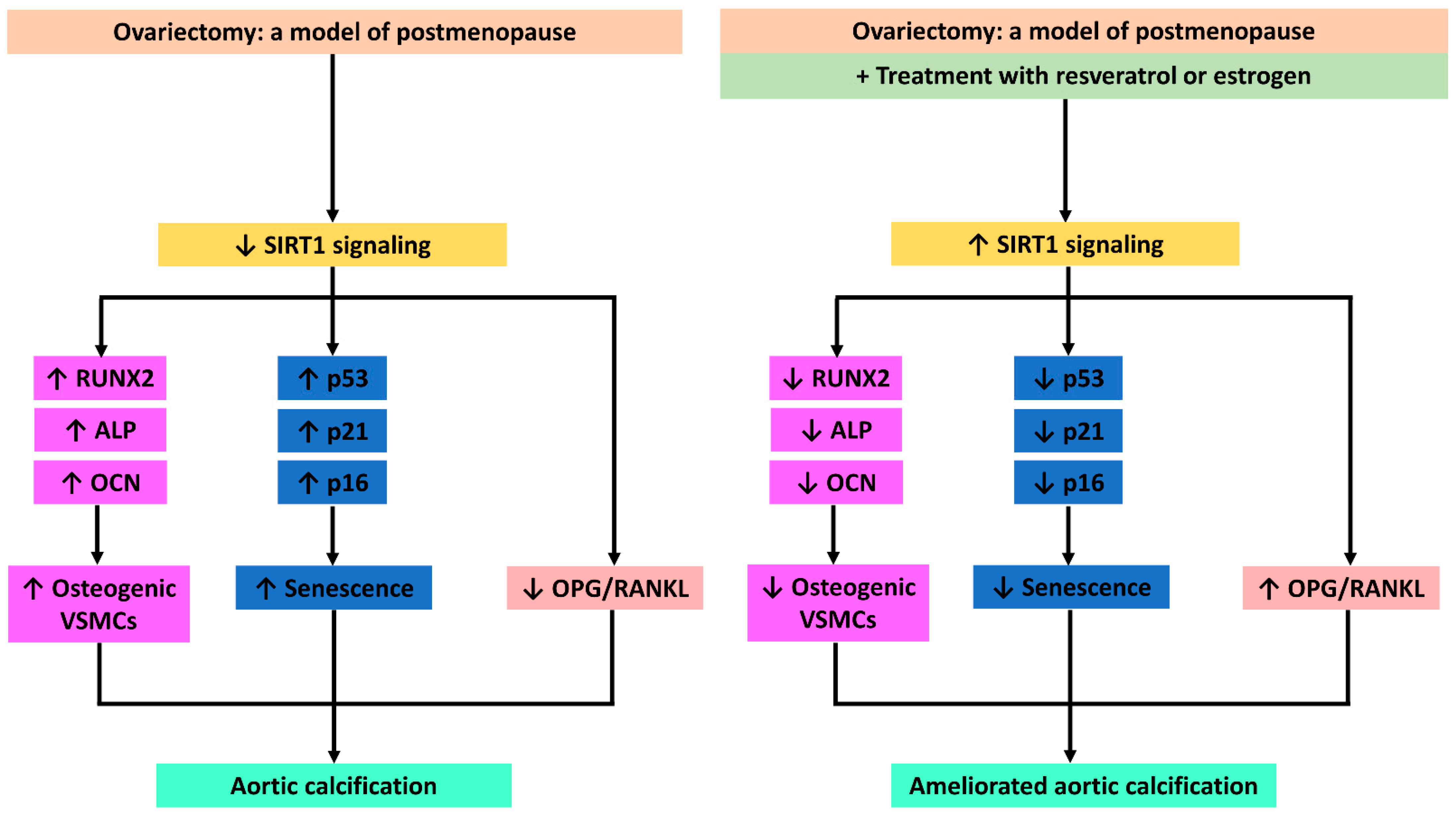
Resveratrol Ameliorates Aortic Calcification in Ovariectomized Rats via SIRT1 Signaling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs
2.2. Animals and Ovariectomy
2.3. Study Design
2.4. Sampling
2.5. Western Blot Analysis
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.8. Histological Study
2.9. Statistical Analysis
3. Results
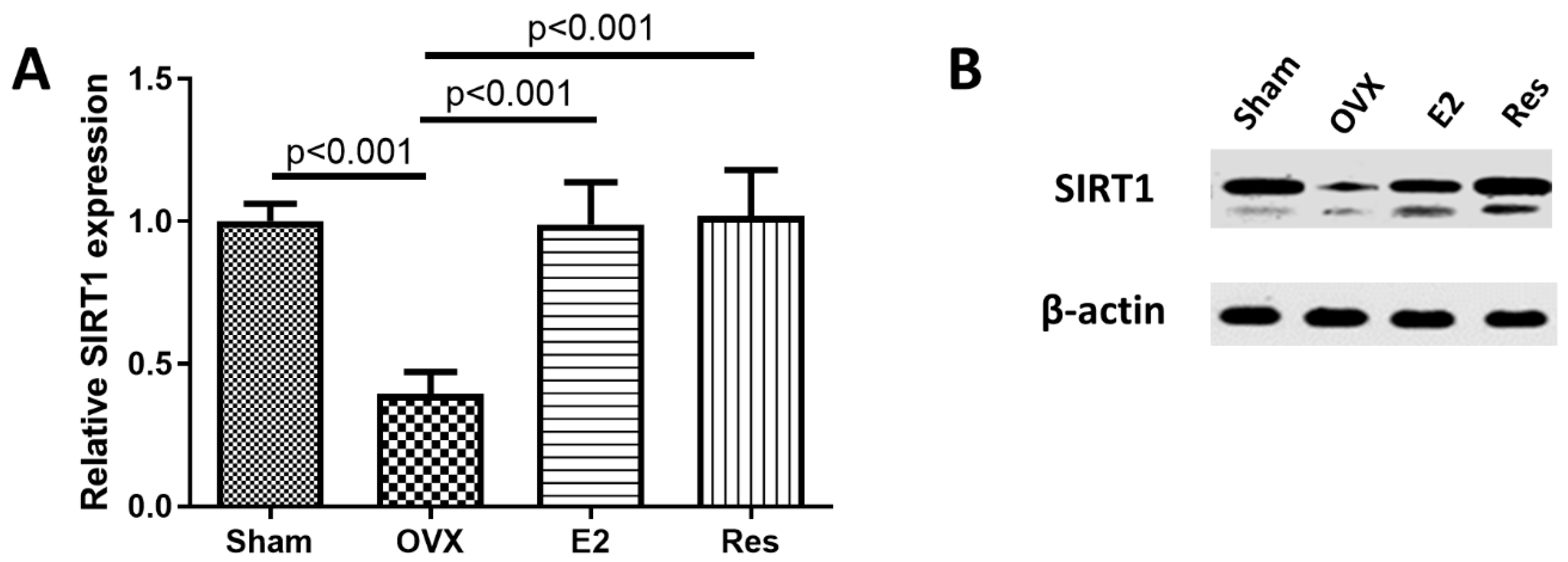
3.1. Resveratrol and E2 Upregulated SIRT1 Expression in the Aortas of Ovariectomized Rats
3.2. Resveratrol and E2 Reduced the Aortic Levels of Osteogenic Markers
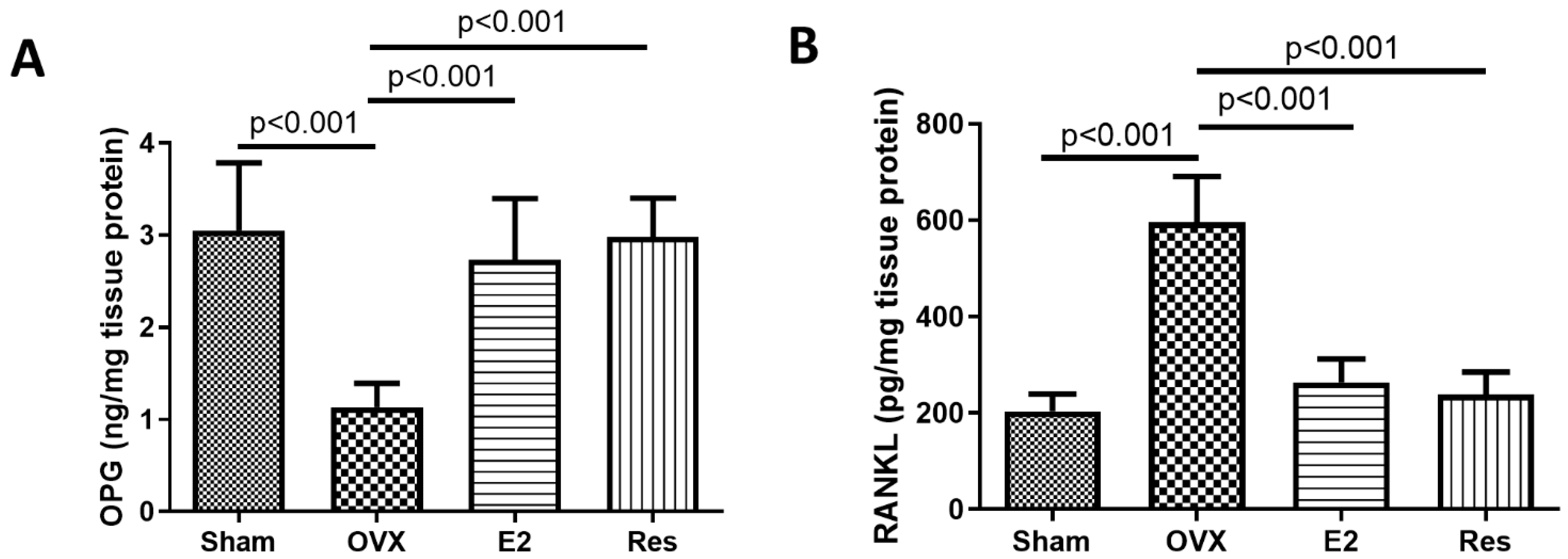
3.3. Resveratrol and E2 Significantly Increased OPG and Decreased RANKL Aortic Levels
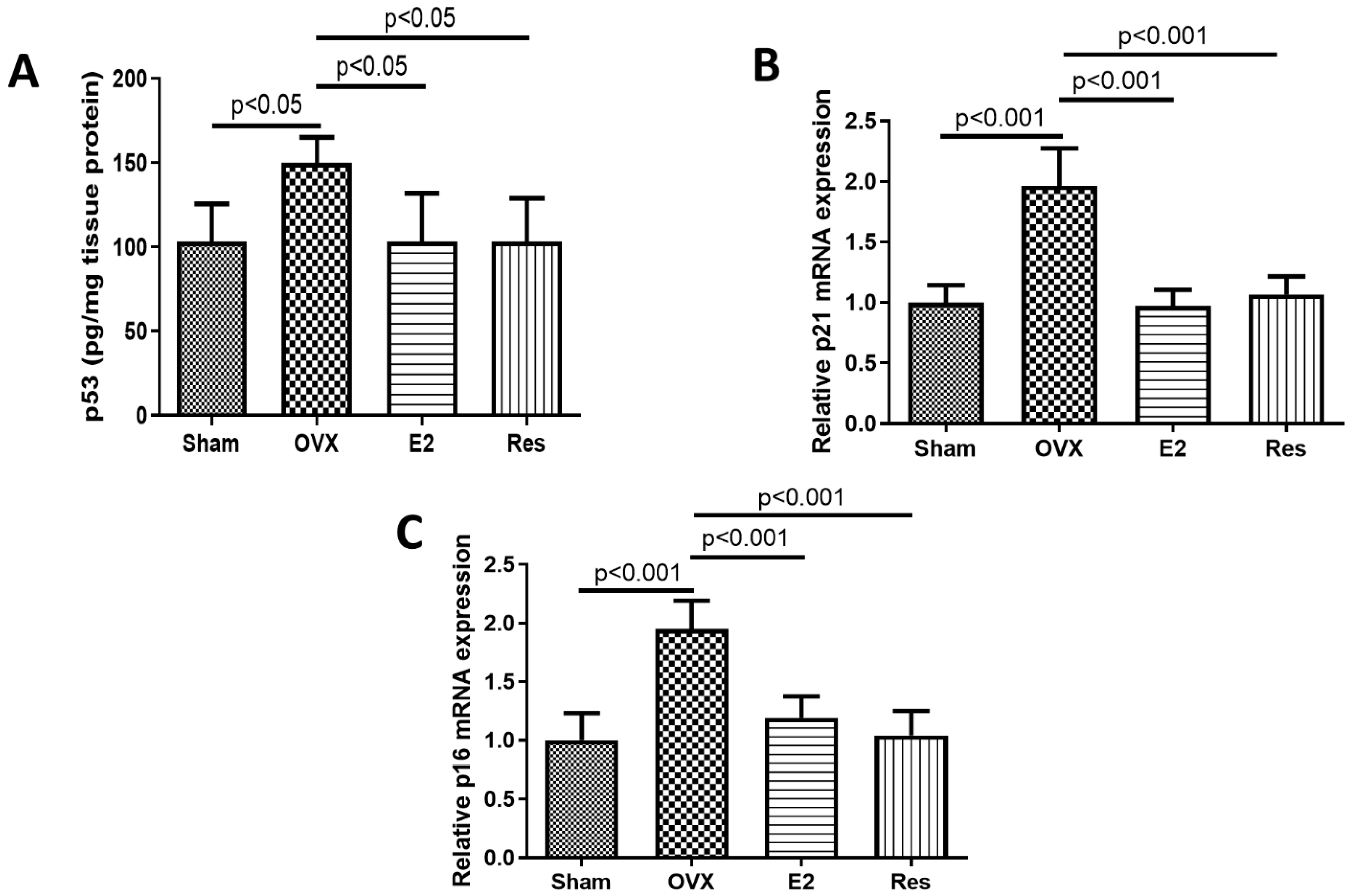
3.4. Resveratrol and E2 Downregulated the Expression of the Senescence Markers p53, p21 and p16 in the Aortas of OVX Rats
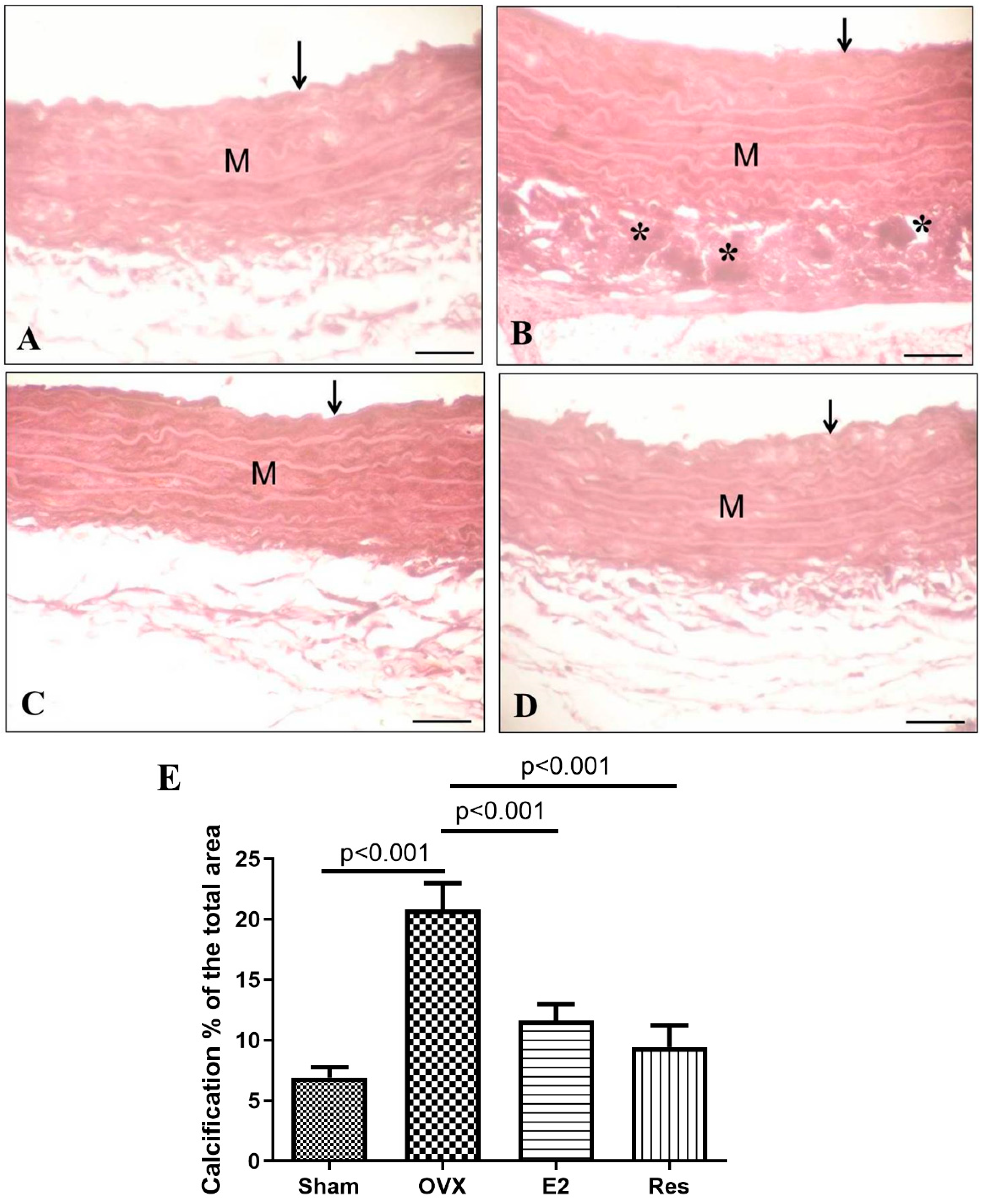
3.5. Resveratrol and E2 Improved Aortic Calcification in OVX Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| E2 | Estradiol |
| H&E | Hematoxylin and eosin |
| OPG | Osteoprotegerin |
| OVX | Ovariectomized |
| RANKL | Receptor activator of nuclear factor-κB ligand |
| Res | Resveratrol |
| RUNX2 | Runt-related transcription factor 2 |
| SIRT1 | Sirtuin 1 |
| VSMCs | Vascular smooth muscle cells |
References
- El Khoudary, S.R.; Aggarwal, B.; Beckie, T.M.; Hodis, H.N.; Johnson, A.E.; Langer, R.D.; Limacher, M.C.; Manson, J.E.; Stefanick, M.L.; Allison, M.A.; et al. Menopause Transition and Cardiovascular Disease Risk: Implications for Timing of Early Prevention: A Scientific Statement from the American Heart Association. Circulation 2020, 142, e506–e532. [Google Scholar] [CrossRef]
- Zhu, D.; Chung, H.-F.; Pandeya, N.; Dobson, A.J.; Cade, J.E.; Greenwood, D.C.; Crawford, S.L.; Avis, N.E.; Gold, E.B.; Mitchell, E.S.; et al. Relationships between intensity, duration, cumulative dose, and timing of smoking with age at menopause: A pooled analysis of individual data from 17 observational studies. PLoS Med. 2018, 15, e1002704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochanek, K.D.; Murphy, S.L.; Xu, J.; Tejada-Vera, B. Deaths: Final Data for 2014. In National Vital Statistics Reports; National Vital Statistics System, National Center for Health Statistics, Centers for Disease Control and Prevention: Atlanta, GA, USA, 2016; Volume 65, pp. 1–122. [Google Scholar]
- WHO. Female Life Expectancy. 2021. Available online: https://www.who.int/gho/women_and_health/mortality/life_expectancy_text/en/ (accessed on 29 July 2021).
- El Khoudary, S.R.; Thurston, R.C. Cardiovascular Implications of the Menopause Transition: Endogenous Sex Hormones and Vasomotor Symptoms. Obstetr. Gynecol. Clin. N. Am. 2018, 45, 641–661. [Google Scholar] [CrossRef]
- Tepper, P.G.; Randolph, J.F., Jr.; McConnell, D.S.; Crawford, S.L.; El Khoudary, S.R.; Joffe, H.; Gold, E.B.; Zheng, H.; Bromberger, J.T.; Sutton-Tyrrell, K. Trajectory Clustering of Estradiol and Follicle-Stimulating Hormone during the Menopausal Transition among Women in the Study of Women’s Health across the Nation (SWAN). J. Clin. Endocrinol. Metab. 2012, 97, 2872–2880. [Google Scholar] [CrossRef]
- El Khoudary, S.R.; Greendale, G.; Crawford, S.L.; Avis, N.E.; Brooks, M.M.; Thurston, R.C.; Karvonen-Gutierrez, C.; Waetjen, L.E.; Matthews, K. The menopause transition and women’s health at midlife: A progress report from the Study of Women’s Health Across the Nation (SWAN). Menopause 2019, 26, 1213–1227. [Google Scholar]
- Buckler, H. The menopause transition: Endocrine changes and clinical symptoms. J. Br. Menopause Soc. 2005, 11, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Thurston, R.C.; Sutton-Tyrrell, K.; Everson-Rose, S.A.; Hess, R.; Matthews, K.A. Hot Flashes and Subclinical Cardiovascular Disease. Circulation 2008, 118, 1234–1240. [Google Scholar] [CrossRef]
- Thurston, R.C.; Sutton-Tyrrell, K.; Everson-Rose, S.A.; Hess, R.; Powell, L.H.; Matthews, K.A. Hot flashes and carotid intima media thickness among midlife women. Menopause 2011, 18, 352–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, K.A.; Everson-Rose, S.A.; Kravitz, H.M.; Lee, L.; Janssen, I.; Sutton-Tyrrell, K. Do reports of sleep disturbance relate to coronary and aortic calcification in healthy middle-aged women? Study of Women’s Health across the Nation. Sleep Med. 2013, 14, 282–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Lee, I.-K.; Jeon, J.-H. Vascular Calcification—New Insights into Its Mechanism. Int. J. Mol. Sci. 2020, 21, 2685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Rementer, C.; Giachelli, C.M. Vascular Calcification: An Update on Mechanisms and Challenges in Treatment. Calcified Tissue Int. 2013, 93, 365–373. [Google Scholar] [CrossRef]
- Kannel, W.B.; Hjortland, M.C.; McNamara, P.M.; Gordon, T. Menopause and Risk of Cardiovascular Disease: The Framingham Study. Ann. Internal Med. 1976, 85, 447–452. [Google Scholar] [CrossRef]
- Xing, D.; Nozell, S.; Chen, Y.-F.; Hage, F.; Oparil, S. Estrogen and mechanisms of vascular protection. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Vinogradova, Y.; Coupland, C.; Hippisley-Cox, J. Use of hormone replacement therapy and risk of breast cancer: Nested case-control studies using the QResearch and CPRD databases. BMJ 2020, 371, m3873. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.-f.; Wu, Y.; Li, C.-y. Hormone therapy and risk of ovarian cancer in postmenopausal women: A systematic review and meta-analysis. Menopause 2016, 23, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, L.L.; Mørch, L.S.; Løkkegaard, E. Hormone replacement therapy and the risk of endometrial cancer: A systematic review. Maturitas 2016, 91, 25–35. [Google Scholar] [CrossRef]
- Gołąbek, A.; Kowalska, K.; Olejnik, A. Polyphenols as a Diet Therapy Concept for Endometriosis—Current Opinion and Future Perspectives. Nutrients 2021, 13, 1347. [Google Scholar] [CrossRef]
- Rietjens, I.M.C.M.; Louisse, J.; Beekmann, K. The potential health effects of dietary phytoestrogens. Br. J. Pharmacol. 2017, 174, 1263–1280. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, Y.; Du, Y.; Li, G.; Wang, L.; Zhou, F. Resveratrol Ameliorated Vascular Calcification by Regulating Sirt-1 and Nrf2. Transpl. Proc. 2016, 48, 3378–3386. [Google Scholar] [CrossRef] [PubMed]
- Navarro, G.; Martínez -Pinilla, E.; Ortiz, R.; Noé, V.; Ciudad, C.J.; Franco, R. Resveratrol and Related Stilbenoids, Nutraceutical/Dietary Complements with Health-Promoting Actions: Industrial Production, Safety, and the Search for Mode of Action. Compr. Rev. Food Sci. Food Saf. 2018, 17, 808–826. [Google Scholar] [CrossRef] [Green Version]
- Shimoyama, Y.; Mitsuda, Y.; Tsuruta, Y.; Suzuki, K.; Hamajima, N.; Niwa, T. SIRTUIN 1 Gene Polymorphisms are Associated with Cholesterol Metabolism and Coronary Artery Calcification in Japanese Hemodialysis Patients. J. Renal Nutr. 2012, 22, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Ramkaran, P.; Phulukdaree, A.; Khan, S.; Moodley, D.; Chuturgoon, A.A. Sirtuin 1 rs1467568 and rs7895833 in South African Indians with early-onset coronary artery disease. Cardiovasc. J. Afr. 2016, 27, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandatori, D.; Pipino, C.; Di Tomo, P.; Schiavone, V.; Ranieri, A.; Pantalone, S.; Di Silvestre, S.; Di Pietrantonio, N.; Ucci, M.; Palmerini, C.; et al. Osteogenic transdifferentiation of vascular smooth muscle cells isolated from spontaneously hypertensive rats and potential menaquinone-4 inhibiting effect. J. Cell. Physiol. 2019, 234, 19761–19773. [Google Scholar] [CrossRef] [PubMed]
- Leopold, J.A. Vascular calcification: Mechanisms of vascular smooth muscle cell calcification. Trends Cardiovasc. Med. 2015, 25, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, X.; Liu, Y.-Z.; Cheng, Z.-B.; Sun, J.-X.; Shao, Y.-D.; Qu, S.-L.; Huang, L.; Zhang, C. Adipokines in vascular calcification. Clin. Chim. Acta 2021, 516, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Drozdov, I.; Shroff, R.; Beltran Leilani, E.; Shanahan Catherine, M. Prelamin A Accelerates Vascular Calcification Via Activation of the DNA Damage Response and Senescence-Associated Secretory Phenotype in Vascular Smooth Muscle Cells. Circul. Res. 2013, 112, e99–e109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartoli-Leonard, F.; Wilkinson, F.L.; Schiro, A.; Inglott, F.S.; Alexander, M.Y.; Weston, R. Suppression of SIRT1 in Diabetic Conditions Induces Osteogenic Differentiation of Human Vascular Smooth Muscle Cells via RUNX2 Signalling. Sci. Rep. 2019, 9, 878. [Google Scholar] [CrossRef]
- García-Gómez, M.C.; Vilahur, G. Osteoporosis and vascular calcification: A shared scenario. Clín. Investig. Arterioscler. Engl. Ed. 2020, 32, 32–41. [Google Scholar] [CrossRef]
- Osako Mariana, K.; Nakagami, H.; Koibuchi, N.; Shimizu, H.; Nakagami, F.; Koriyama, H.; Shimamura, M.; Miyake, T.; Rakugi, H.; Morishita, R. Estrogen Inhibits Vascular Calcification via Vascular RANKL System. Circul. Res. 2010, 107, 466–475. [Google Scholar] [CrossRef] [Green Version]
- Ndip, A.; Williams, A.; Jude, E.B.; Serracino-Inglott, F.; Richardson, S.; Smyth, J.V.; Boulton, A.J.M.; Alexander, M.Y. The RANKL/RANK/OPG Signaling Pathway Mediates Medial Arterial Calcification in Diabetic Charcot Neuroarthropathy. Diabetes 2011, 60, 2187. [Google Scholar] [CrossRef] [Green Version]
- Ndip, A.; Wilkinson, F.L.; Jude, E.B.; Boulton, A.J.M.; Alexander, M.Y. RANKL–OPG and RAGE modulation in vascular calcification and diabetes: Novel targets for therapy. Diabetologia 2014, 57, 2251–2260. [Google Scholar] [CrossRef]
- Idris, A.I. Ovariectomy/Orchidectomy in Rodents. In Bone Research Protocols; Helfrich, M.H., Ralston, S.H., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 545–551. [Google Scholar]
- Zhao, H.; Niu, Q.; Li, X.; Liu, T.; Xu, Y.; Han, H.; Wang, W.; Fan, N.; Tian, Q.; Zhang, H.; et al. Long-term resveratrol consumption protects ovariectomized rats chronically treated with d-galactose from developing memory decline without effects on the uterus. Brain Res. 2012, 1467, 67–80. [Google Scholar] [CrossRef]
- Little, R.; Zi, M.; Hammad, S.K.; Nguyen, L.; Njegic, A.; Kurusamy, S.; Prehar, S.; Armesilla, A.L.; Neyses, L.; Austin, C.; et al. Reduced expression of PMCA1 is associated with increased blood pressure with age which is preceded by remodelling of resistance arteries. Aging Cell 2017, 16, 1104–1113. [Google Scholar] [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Liou, S.-F.; Nguyen, T.T.N.; Hsu, J.-H.; Sulistyowati, E.; Huang, S.-E.; Wu, B.-N.; Lin, M.-C.; Yeh, J.-L. The Preventive Effects of Xanthohumol on Vascular Calcification Induced by Vitamin D3 Plus Nicotine. Antioxidants 2020, 9, 956. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, L.; Chen, S.; Liu, X.; Li, H.; Lu, X.; Yang, X.; Huang, J.; Gu, D. Association between the SIRT1 mRNA Expression and Acute Coronary Syndrome. J. Atheroscler. Thromb. 2015, 22, 165–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thangavel, P.; Puga-Olguín, A.; Rodríguez-Landa, J.F.; Zepeda, R.C. Genistein as Potential Therapeutic Candidate for Menopausal Symptoms and Other Related Diseases. Molecules 2019, 24, 3892. [Google Scholar] [CrossRef] [Green Version]
- Lampropoulos, C.E.; Kalamara, P.; Konsta, M.; Papaioannou, I.; Papadima, E.; Antoniou, Z.; Andrianopoulou, A.; Vlachoyiannopoulos, P.G. Osteoporosis and vascular calcification in postmenopausal women: A cross-sectional study. Climacteric 2016, 19, 303–307. [Google Scholar] [CrossRef]
- Yan, J.; Stringer Sally, E.; Hamilton, A.; Charlton-Menys, V.; Götting, C.; Müller, B.; Aeschlimann, D.; Alexander, M.Y. Decorin GAG Synthesis and TGF-β Signaling Mediate Ox-LDL–Induced Mineralization of Human Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 608–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Tanaka, T.; Iso, T.; Kawai-Kowase, K.; Kurabayashi, M. Azelnidipine Inhibits Msx2-Dependent Osteogenic Differentiation and Matrix Mineralization of Vascular Smooth Muscle Cells. Int. Heart J. 2012, 53, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, T.; Yan, J.; Jiagbogu, N.; Heideman, D.A.M.; Canfield, A.E.; Alexander, M.Y. HGF/c-Met signalling promotes Notch3 activation and human vascular smooth muscle cell osteogenic differentiation in vitro. Atherosclerosis 2011, 219, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.; Ikeda, Y.; Miyauchi, T.; Uchikado, Y.; Akasaki, Y.; Ohishi, M. Estrogen-SIRT1 Axis Plays a Pivotal Role in Protecting Arteries Against Menopause-Induced Senescence and Atherosclerosis. J. Atheroscler. Thromb. 2020, 27, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, T.; Takei, A.; Tsujikado, K.; Inukai, T. Effects of androgens and estrogens on sirtuin 1 gene expression in human aortic endothelial cells. Saudi Med. J. 2020, 41, 361–368. [Google Scholar] [CrossRef]
- Sasaki, Y.; Ikeda, Y.; Uchikado, Y.; Akasaki, Y.; Sadoshima, J.; Ohishi, M. Estrogen Plays a Crucial Role in Rab9-Dependent Mitochondrial Autophagy, Delaying Arterial Senescence. J. Am. Heart Assoc. 2021, 10, e019310. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhang, Y.; Zhang, M.; Guo, L.; Wang, J.; Zeng, F.; Xu, D.; Yin, Z.; Xu, Y.; Wang, D.; et al. Interleukin-1β-Induced Senescence Promotes Osteoblastic Transition of Vascular Smooth Muscle Cells. Kidney Blood Pressure Res. 2020, 45, 314–330. [Google Scholar] [CrossRef]
- Singh, A.; Tandon, S.; Tandon, C. An update on vascular calcification and potential therapeutics. Mol. Biol. Rep. 2021, 48, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Millar, S.A.; Patel, H.; Anderson, S.I.; England, T.J.; O’Sullivan, S.E. Osteocalcin, Vascular Calcification, and Atherosclerosis: A Systematic Review and Meta-analysis. Front. Endocrinol. 2017, 8, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- London, G.M. Mechanisms of arterial calcifications and consequences for cardiovascular function. Kidney Int. Suppl. 2013, 3, 442–445. [Google Scholar] [CrossRef] [Green Version]
- Patel, J.J.; Zhu, D.; Opdebeeck, B.; D’Haese, P.; Millán, J.L.; Bourne, L.E.; Wheeler-Jones, C.P.D.; Arnett, T.R.; MacRae, V.E.; Orriss, I.R. Inhibition of arterial medial calcification and bone mineralization by extracellular nucleotides: The same functional effect mediated by different cellular mechanisms. J. Cell. Physiol. 2018, 233, 3230–3243. [Google Scholar] [CrossRef] [PubMed]
- Takemura, A.; Iijima, K.; Ota, H.; Son, B.K.; Ito, Y.; Ogawa, S.; Eto, M.; Akishita, M.; Ouchi, Y. Sirtuin 1 retards hyperphosphatemia-induced calcification of vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2054–2062. [Google Scholar] [CrossRef] [Green Version]
- Weiss, R.M.; Lund, D.D.; Chu, Y.; Brooks, R.M.; Zimmerman, K.A.; Accaoui, R.E.; Davis, M.K.; Hajj, G.P.; Zimmerman, M.B.; Heistad, D.D. Osteoprotegerin Inhibits Aortic Valve Calcification and Preserves Valve Function in Hypercholesterolemic Mice. PLoS ONE 2013, 8, e65201. [Google Scholar] [CrossRef] [Green Version]
- Bucay, N.; Sarosi, I.; Dunstan, C.R.; Morony, S.; Tarpley, J.; Capparelli, C.; Scully, S.; Tan, H.L.; Xu, W.; Lacey, D.L.; et al. Osteoprotegerin-deficient mice develop early onset osteoporosis and arterial calcification. Genes Dev. 1998, 12, 1260–1268. [Google Scholar] [CrossRef]
- Callegari, A.; Coons, M.L.; Ricks, J.L.; Rosenfeld, M.E.; Scatena, M. Increased Calcification in Osteoprotegerin-Deficient Smooth Muscle Cells: Dependence on Receptor Activator of NF-κB Ligand and Interleukin 6. J. Vasc. Res. 2014, 51, 118–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elseweidy, M.M.; El-Swefy, S.E.; Shaheen, M.A.; Baraka, N.M.; Hammad, S.K. Effect of resveratrol and mesenchymal stem cell monotherapy and combined treatment in management of osteoporosis in ovariectomized rats: Role of SIRT1/FOXO3a and Wnt/β-catenin pathways. Arch. Biochem. Biophys. 2021, 703, 108856. [Google Scholar] [CrossRef]
- Hofbauer, L.C.; Khosla, S.; Dunstan, C.R.; Lacey, D.L.; Spelsberg, T.C.; Riggs, B.L. Estrogen Stimulates Gene Expression and Protein Production of Osteoprotegerin in Human Osteoblastic Cells. Endocrinology 1999, 140, 4367–4370. [Google Scholar] [CrossRef]
- Bond, J.; Haughton, M.; Blaydes, J.; Gire, V.; Wynford-Thomas, D.; Wyllie, F. Evidence that transcriptional activation by p53 plays a direct role in the induction of cellular senescence. Oncogene 1996, 13, 2097–2104. [Google Scholar] [PubMed]
- Luo, J.; Su, F.; Chen, D.; Shiloh, A.; Gu, W. Deacetylation of p53 modulates its effect on cell growth and apoptosis. Nature 2000, 408, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Miyauchi, H.; Yoshida, T.; Ishida, Y.; Yoshida, H.; Komuro, I. Endothelial Cell Senescence in Human Atherosclerosis. Circulation 2002, 105, 1541–1544. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Li, M.; Tang, Y.; Laszkowska, M.; Roeder, R.G.; Gu, W. Acetylation of p53 augments its site-specific DNA binding both in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2004, 101, 2259–2264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, S.M.; Quelle, D.E. p53 Acetylation: Regulation and Consequences. Cancers 2015, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Pan, K.; Chen, Y.; Huang, C.; Zhang, X. The acetylation of transcription factor HBP1 by p300/CBP enhances p16INK4A expression. Nucleic Acids Res. 2012, 40, 981–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hammad, S.K.; Eissa, R.G.; Shaheen, M.A.; Younis, N.N. Resveratrol Ameliorates Aortic Calcification in Ovariectomized Rats via SIRT1 Signaling. Curr. Issues Mol. Biol. 2021, 43, 1057-1071. https://doi.org/10.3390/cimb43020075
Hammad SK, Eissa RG, Shaheen MA, Younis NN. Resveratrol Ameliorates Aortic Calcification in Ovariectomized Rats via SIRT1 Signaling. Current Issues in Molecular Biology. 2021; 43(2):1057-1071. https://doi.org/10.3390/cimb43020075
Chicago/Turabian StyleHammad, Sally K., Rana G. Eissa, Mohamed A. Shaheen, and Nahla N. Younis. 2021. "Resveratrol Ameliorates Aortic Calcification in Ovariectomized Rats via SIRT1 Signaling" Current Issues in Molecular Biology 43, no. 2: 1057-1071. https://doi.org/10.3390/cimb43020075
APA StyleHammad, S. K., Eissa, R. G., Shaheen, M. A., & Younis, N. N. (2021). Resveratrol Ameliorates Aortic Calcification in Ovariectomized Rats via SIRT1 Signaling. Current Issues in Molecular Biology, 43(2), 1057-1071. https://doi.org/10.3390/cimb43020075