
Soluble Expression of a Neo2/15-Conjugated Single Chain Fv against PD-L1 in Escherichia coli

Abstract
:1. Introduction
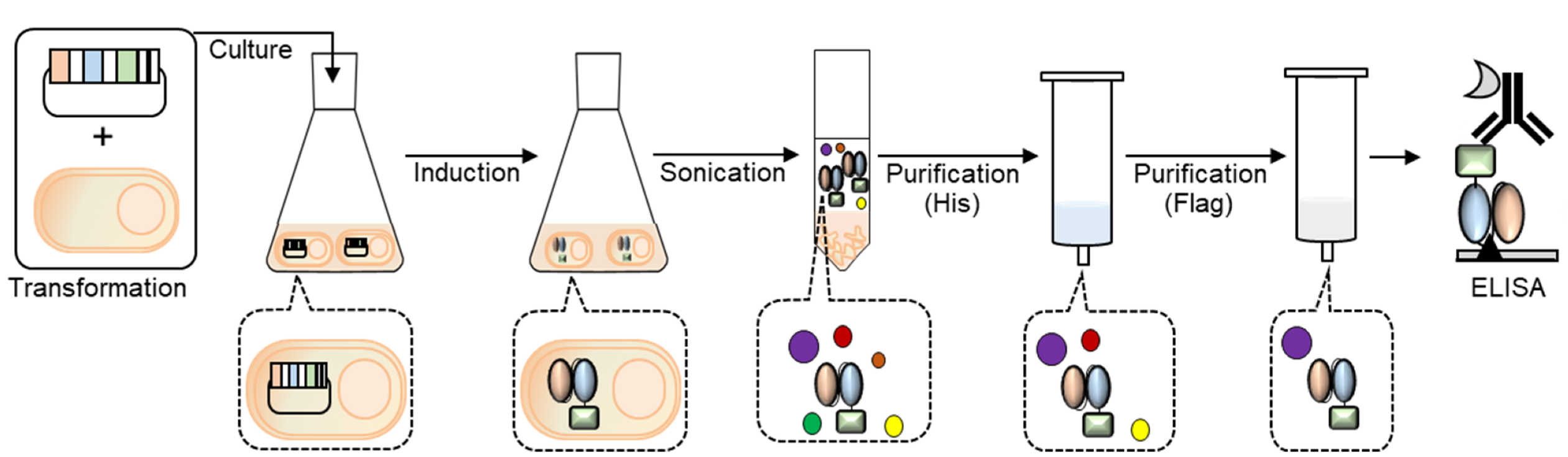
2. Materials and Methods
2.1. Materials
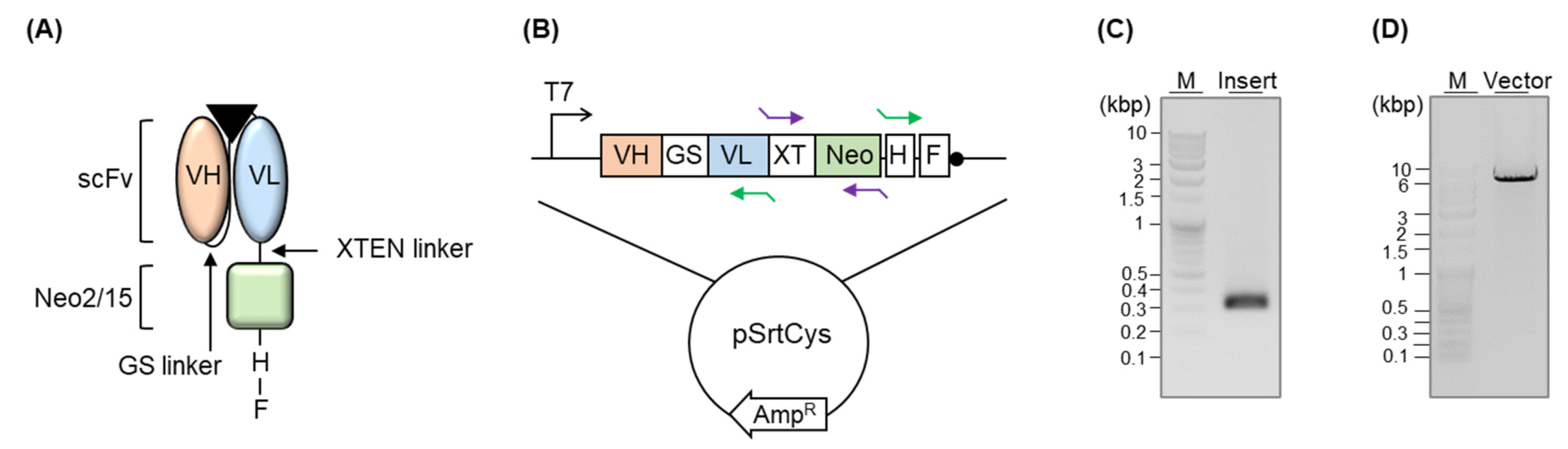
2.2. Gene Construction
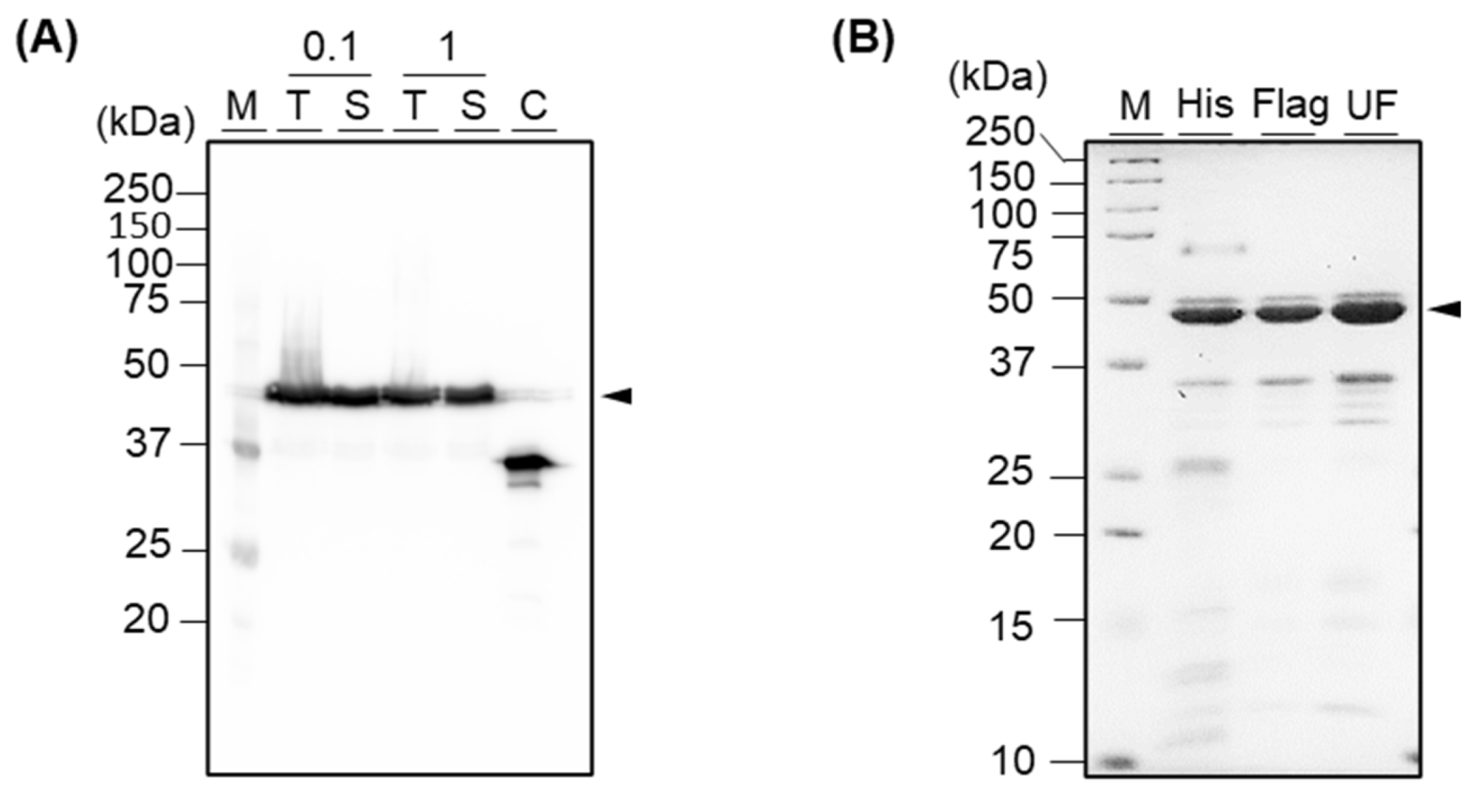
2.3. Expression
2.4. Western Blot Analysis
2.5. Talon Purification
2.6. Flag Purification
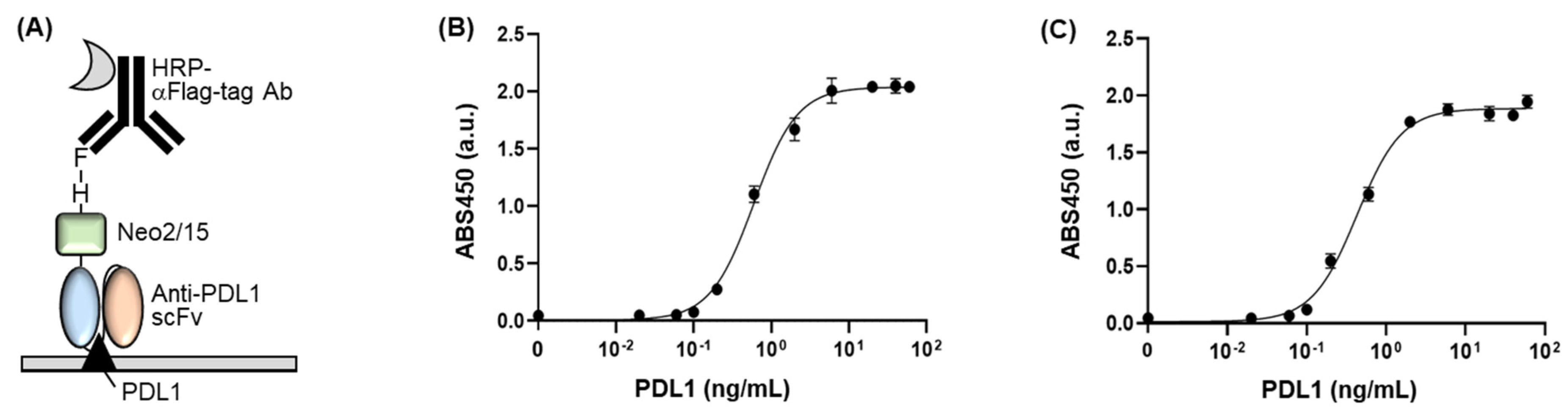
2.7. Enzyme-Linked Immunosorbent Assay
3. Results
3.1. Construction of Immunocytokine-Expressing Plasmid
3.2. Expression and Purification
3.3. Confirmation of Antigen-Binding Efficiency
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sharma, P.; Allison, J.P. Immune checkpoint targeting in cancer therapy: Toward combination strategies with curative potential. Cell 2015, 161, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-H.; Jeong, H.-J. Soluble expression of small antibody fragments against PD-L1 using Escherichia coli with high yield and purity. Appl. Sci. 2021, 11, 9149. [Google Scholar] [CrossRef]
- Beckman, R.A.; Weiner, L.M.; Davis, H.M. Antibody constructs in cancer therapy: Protein engineering strategies to improve exposure in solid tumors. Cancer 2007, 109, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Milenic, D.E.; Whitlow, M.; Schlom, J. Rapid tumor penetration of a single-chain Fv and comparison with other immunoglobulin forms. Cancer Res. 1992, 52, 3402–3408. [Google Scholar] [PubMed]
- Terpe, K. Overview of bacterial expression systems for heterologous protein production: From molecular and biochemical fundamentals to commercial systems. Appl. Microbiol. Biotechnol. 2006, 72, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Makrides, S.C. Strategies for achieving high-level expression of genes in Escherichia coli. Microbiol. Rev. 1996, 60, 512–538. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Li, X.; Dai, J.; Cole, L.; Camacho, J.A.; Zhang, Y.; Ji, Y.; Wang, J.; Yang, X.-F.; Wang, H. Immune cell subset differentiation and tissue inflammation. J. Hematol. Oncol. 2018, 11, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi Kalateh Shah Mohammad, G.; Ghahremanloo, A.; Soltani, A.; Fathi, E.; Hashemy, S.I. Cytokines as potential combination agents with PD-1/PD-L1 blockade for cancer treatment. J. Cell. Physiol. 2020, 235, 5449–5460. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.-A.; Yu, S.; Ulge, U.Y.; Spangler, J.B.; Jude, K.M.; Labão-Almeida, C.; Ali, L.R.; Quijano-Rubio, A.; Ruterbusch, M.; Leung, I.; et al. De novo design of potent and selective mimics of IL-2 and IL-15. Nature 2019, 565, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Barroso-Sousa, R.; Ott, P.A. Transformation of old concepts for a new era of cancer immunotherapy: Cytokine therapy and cancer vaccines as combination partners of PD1/PD-L1 inhibitors. Curr. Oncol. Rep. 2018, 21, 1. [Google Scholar] [CrossRef] [PubMed]
- Pico de Coaña, Y.; Choudhury, A.; Kiessling, R. Checkpoint blockade for cancer therapy: Revitalizing a suppressed immune system. Trends Mol. Med. 2015, 21, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Wrangle, J.M.; Velcheti, V.; Patel, M.R.; Garrett-Mayer, E.; Hill, E.G.; Ravenel, J.G.; Miller, J.S.; Farhad, M.; Anderton, K.; Lindsey, K.; et al. ALT-803, an IL-15 superagonist, in combination with nivolumab in patients with metastatic non-small cell lung cancer: A non-randomised, open-label, phase 1b trial. Lancet Oncol. 2018, 19, 694–704. [Google Scholar] [CrossRef]
- Juliá, E.P.; Amante, A.; Pampena, M.B.; Mordoh, J.; Levy, E.M. Avelumab, an IgG1 anti-PD-L1 immune checkpoint inhibitor, triggers NK cell-mediated cytotoxicity and cytokine production against triple negative breast cancer cells. Front. Immunol. 2018, 9, 2140. [Google Scholar] [CrossRef] [PubMed]
- Lode, H.N.; Xiang, R.; Becker, J.C.; Gillies, S.D.; Reisfeld, R.A. Immunocytokines: A promising approach to cancer immunotherapy. Pharmacol. Ther. 1999, 80, 277–292. [Google Scholar] [CrossRef]
- Penichet, M.L.; Morrison, S.L. Antibody-cytokine fusion proteins for the therapy of cancer. J. Immunol. Methods 2001, 248, 91–101. [Google Scholar] [CrossRef]
- Young, P.A.; Morrison, S.L.; Timmerman, J.M. Antibody-cytokine fusion proteins for treatment of cancer: Engineering cytokines for improved efficacy and safety. Semin. Oncol. 2014, 41, 623–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neri, D.; Sondel, P.M. Immunocytokines for cancer treatment: Past, present and future. Curr. Opin. Immunol. 2016, 40, 96–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutmacher, C.; Neri, D. Antibody-cytokine fusion proteins: Biopharmaceuticals with immunomodulatory properties for cancer therapy. Adv. Drug Deliv. Rev. 2019, 141, 67–91. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | |
|---|---|
| Nucleotide (5′-3′) | atgaaaaagacagctatcgcgattgcagtggcactggctggtttcgctaccgtggcccaggcggccctg actcagccgtcctcggtgtcagcaaacctgggaggaaccgtcaagatcacctgctccgggggtagtggc agctacggctggtatcagcagaaggcacctggcagtgcccctgtcagtctgatctatgacaacaccaac agaccctcggacatcccttcacgattctccggtgccctatccggctccacagccacattaaccatcactgg aggtccaagccgaggacgaggctgtctattactgtgggagcagggacagcagtaatgctggttctgtatt tggggccgggacaaccctgaccgtcctaggtcagtcctctagatcttccggcggtggtggcagctccgg tggtggcggttccgccctgacgttggacgagtccgggggcggcctccagacgcccggaggagcgct cagcctcgtctgcaaggcctccgggttcaccttcagtgaccgtggcatgcactgggtgcgacaggcgcc cggcaaggggctggagtgggtcggtgctattagcaggagagggagtaccacaacttacgcacccgcgg tgaagggccgtgccaccatcacgagggacaacgggcagagcacagtgaggctgcagctgaacaacct cactgctgaggacaccgccacctacttctgcgccaaaaatgatgattctgtcggtatagtgactacttctac tatcgacgcatggggccacgggaccgaagtcatcgtctcctccactagtggccaggccggccagagcg gcagcgagactcccgggacctcagagtccgccacacccgaaagtcccaagaagaagatccaattacatgct gaacatgcactgtatgacgccttgatgatcttgaatattgtcaaaaccaactcgccgccggcagaagagaagcttg aagattatgcatttaattttgaacttatccttgaggaaattgcacgtttattcgaaagtggtgatcaaaaagatgaag ccgagaaggccaagcgcatgaaagagtggatgaaacgtatcaagaccaccgcttcagaggatgagcaggagga gatggcgaacgcgattattacaatcctgcagagttggatcttttcaggggggggttctcatcatcatcatcatcatg gcggatccgactacaaggacgacgatgacaaa |
| Amino acid (N′-C′) | MKKTAIAIAVALAGFATVAQAALTQPSSVSANLGGTVKITCSGGSGSY GWYQQKAPGSAPVSLIYDNTNRPSDIPSRFSGALSGSTATLTITGVQAE DEAVYYCGSRDSSNAGSVFGAGTTLTVLGQSSRSSGGGGSSGGGGSAL TLDESGGGLQTPGGALSLVCKASGFTFSDRGMHWVRQAPGKGLEW VGAISRRGSTTTYAPAVKGRATITRDNGQSTVRLQLNNLTAEDTATYFC AKNDDSVGIVTTSTIDAWGHGTEVIVSSTSGQAGQSGSETPGTSESAT PESPKKKIQLHAEHALYDALMILNIVKTNSPPAEEKLEDYAFNFELILE EIARLFESGDQKDEAEKAKRMKEWMKRIKTTASEDEQEEMANAIITIL QSWIFSGGGSHHHHHHGGSDYKDDDDK |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.-H.; Jeong, H.-J. Soluble Expression of a Neo2/15-Conjugated Single Chain Fv against PD-L1 in Escherichia coli. Curr. Issues Mol. Biol. 2022, 44, 301-308. https://doi.org/10.3390/cimb44010022
Kim S-H, Jeong H-J. Soluble Expression of a Neo2/15-Conjugated Single Chain Fv against PD-L1 in Escherichia coli. Current Issues in Molecular Biology. 2022; 44(1):301-308. https://doi.org/10.3390/cimb44010022
Chicago/Turabian StyleKim, Sun-Hee, and Hee-Jin Jeong. 2022. "Soluble Expression of a Neo2/15-Conjugated Single Chain Fv against PD-L1 in Escherichia coli" Current Issues in Molecular Biology 44, no. 1: 301-308. https://doi.org/10.3390/cimb44010022
APA StyleKim, S. -H., & Jeong, H. -J. (2022). Soluble Expression of a Neo2/15-Conjugated Single Chain Fv against PD-L1 in Escherichia coli. Current Issues in Molecular Biology, 44(1), 301-308. https://doi.org/10.3390/cimb44010022