
Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application

Abstract
:1. Introduction
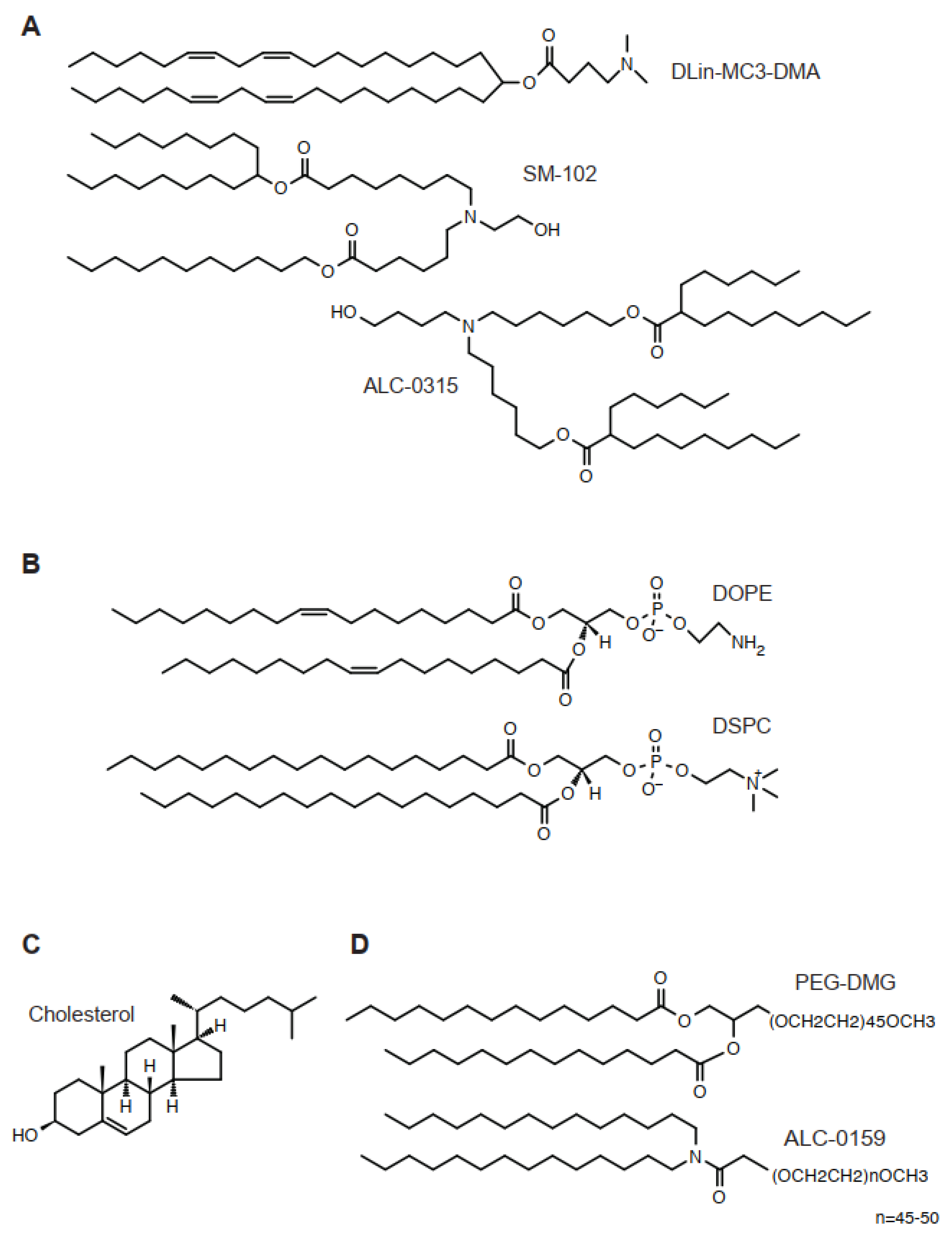
2. Lipids
2.1. Ionizable Cationic Lipids
2.2. Helper Lipids
2.3. Cholesterol
2.4. PEG-Lipids
3. RNA
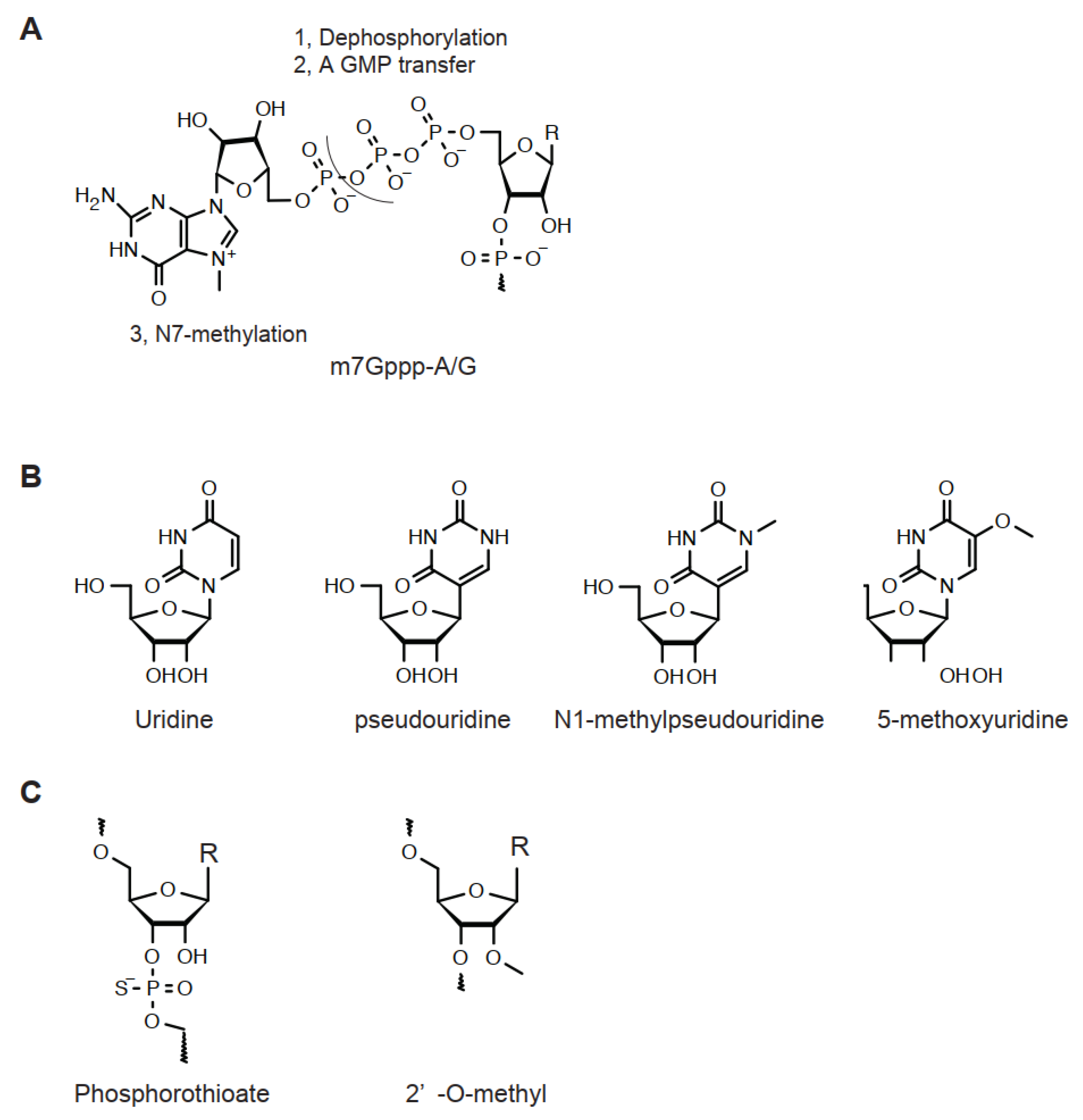
3.1. Modification of mRNA
3.1.1. 5′-Capping
3.1.2. Uridine Analog, Codon Optimization, and PolyA
3.2. Modification of Single Guide RNA (sgRNA)
4. Genetic Material
4.1. siRNA
4.2. mRNA
4.2.1. Therapeutic cDNA
4.2.2. Cas9 Nuclease
4.2.3. Base Editor/Prime Editor
5. Preclinical Study
5.1. Liver Disorders
5.2. Anti-Cancer Therapy
5.3. Congenital Disorders
5.4. Bleeding Disorder
5.5. Hemophilia A and B
5.6. Duchenne Muscular Dystrophy (DMD)
5.7. Human Immunodeficiency Virus (HIV)
6. Clinical Study
6.1. Transthyretin
6.1.1. siRNA
6.1.2. Cas9 Nuclease
6.2. Solid Tumor
6.3. Hepatic Fibrosis
6.4. Ornithine Transcarbamylase Deficiency (OTCD)
7. Tissue Targeting of LNP
7.1. Dendrimers
7.2. Polymers
7.3. Antibody-Modified LNPs
8. Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kulkarni, J.A.; Cullis, P.R.; van der Meel, R. Lipid Nanoparticles Enabling Gene Therapies: From Concepts to Clinical Utility. Nucleic Acid Ther. 2018, 28, 146–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.H.Y.; Brimacombe, C.A.; Verbeke, R.; Cullis, P.R. Exciting Times for Lipid Nanoparticles: How Canadian Discoveries Are Enabling Gene Therapies. Mol. Pharm. 2022, 19, 1663–1668. [Google Scholar] [CrossRef]
- Wilbie, D.; Walther, J.; Mastrobattista, E. Delivery Aspects of CRISPR/Cas for in Vivo Genome Editing. Acc. Chem. Res. 2019, 52, 1555–1564. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, S.J.; Warzecha, C.C.; Yadavali, S.; El-Mayta, R.; Alameh, M.G.; Wang, L.; Weissman, D.; Wilson, J.M.; Issadore, D.; Mitchell, M.J. Scalable mRNA and siRNA Lipid Nanoparticle Production Using a Parallelized Microfluidic Device. Nano Lett. 2021, 21, 5671–5680. [Google Scholar] [CrossRef]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.-C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi Therapeutic, for Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; el Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Witzigmann, D.; Kulkarni, J.A.; Leung, J.; Chen, S.; Cullis, P.R.; van der Meel, R. Lipid nanoparticle technology for therapeutic gene regulation in the liver. Adv. Drug Deliv. Rev. 2020, 159, 344–363. [Google Scholar] [CrossRef]
- Francia, V.; Schiffelers, R.M.; Cullis, P.R.; Witzigmann, D. The Biomolecular Corona of Lipid Nanoparticles for Gene Therapy. Bioconjug. Chem. 2020, 31, 2046–2059. [Google Scholar] [CrossRef]
- Kulkarni, J.A.; Witzigmann, D.; Thomson, S.B.; Chen, S.; Leavitt, B.R.; Cullis, P.R.; van der Meel, R. The current landscape of nucleic acid therapeutics. Nat. Nanotechnol. 2021, 16, 630–643. [Google Scholar] [CrossRef]
- Lee, S.M.; Cheng, Q.; Yu, X.; Liu, S.; Johnson, L.T.; Siegwart, D.J. A Systematic Study of Unsaturation in Lipid Nanoparticles Leads to Improved mRNA Transfection In Vivo. Angew. Chem. Int. Ed. Engl. 2021, 60, 5848–5853. [Google Scholar] [CrossRef]
- Álvarez-Benedicto, E.; Farbiak, L.; Ramírez, M.M.; Wang, X.; Johnson, L.T.; Mian, O.; Guerrero, E.D.; Siegwart, D.J. Optimization of phospholipid chemistry for improved lipid nanoparticle (LNP) delivery of messenger RNA (mRNA). Biomater. Sci. 2022, 10, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Munye, M.M.; Tagalakis, A.D.; Manunta, M.D.I.; Hart, S.L. The role of the helper lipid on the DNA transfection efficiency of lipopolyplex formulations. Sci. Rep. 2014, 4, 7107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollu, A.; Peters, A.; Rentmeister, A. Chemo-Enzymatic Modification of the 5′ Cap to Study mRNAs. Acc. Chem. Res. 2022, 55, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- van Dülmen, M.; Muthmann, N.; Rentmeister, A. Chemo-Enzymatic Modification of the 5′ Cap Maintains Translation and Increases Immunogenic Properties of mRNA. Angew. Chem. Int. Ed. Engl. 2021, 60, 13280–13286. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Bhattacharya, M.; Lee, S.S. From COVID-19 to Cancer mRNA Vaccines: Moving from Bench to Clinic in the Vaccine Landscape. Front. Immunol. 2021, 12, 679344. [Google Scholar] [CrossRef]
- Nakahama, T.; Kawahara, Y. Adenosine-to-inosine RNA editing in the immune system: Friend or foe? Cell Mol Life Sci. 2020, 77, 2931–2948. [Google Scholar] [CrossRef]
- Qin, S.; Tang, X.; Chen, Y.; Chen, K.; Fan, N.; Xiao, W.; Zheng, Q.; Li, G.; Teng, Y.; Wu, M.; et al. mRNA-based therapeutics: Powerful and versatile tools to combat diseases. Signal Transduct. Target Ther. 2022, 7, 166. [Google Scholar] [CrossRef]
- Melamed, J.R.; Hajj, K.A.; Chaudhary, N.; Strelkova, D.; Arral, M.L.; Pardi, N.; Alameh, M.G.; Miller, J.B.; Farbiak, L.; Siegwart, D.J.; et al. Lipid nanoparticle chemistry determines how nucleoside base modifications alter mRNA delivery. J. Control. Release 2022, 341, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Loughrey, D.; Dahlman, J.E. Non-liver mRNA Delivery. Acc. Chem. Res. 2022, 55, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, D.; Paskaleva, E.; Vargason, T.; Ke, X.; Mccallum, S.A.; Linhardt, R.J.; Dordick, J.S. Elucidating the unusual reaction kinetics of D-glucuronyl C5-epimerase. Glycobiology 2020, 30, 847–858. [Google Scholar] [CrossRef]
- Vargason, A.M.; Anselmo, A.C.; Mitragotri, S. The evolution of commercial drug delivery technologies. Nat. Biomed. Eng. 2021, 5, 951–967. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Allen, T.M.; Cullis, P.R. Lipid nanoparticle delivery systems for siRNA-based therapeutics. Drug Deliv. Transl. Res. 2014, 4, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chang, J.; Jiang, Y.; Meng, X.; Sun, T.; Mao, L.; Xu, Q.; Wang, M. Fast and Efficient CRISPR/Cas9 Genome Editing In Vivo Enabled by Bioreducible Lipid and Messenger RNA Nanoparticles. Adv Mater. 2019, 31, e1902575. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, J.A.; Witzigmann, D.; Chen, S.; Cullis, P.R.; van der Meel, R. Lipid Nanoparticle Technology for Clinical Translation of siRNA Therapeutics. Acc. Chem. Res. 2019, 52, 2435–2444. [Google Scholar] [CrossRef] [PubMed]
- Thess, A.; Grund, S.; Mui, B.L.; Hope, M.J.; Baumhof, P.; Fotin-Mleczek, M.; Schlake, T. Sequence-engineered mRNA Without Chemical Nucleoside Modifications Enables an Effective Protein Therapy in Large Animals. Mol. Ther. 2015, 23, 1456–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeRosa, F.; Guild, B.; Karve, S.; Smith, L.; Love, K.; Dorkin, J.R.; Kauffman, K.J.; Zhang, J.; Yahalom, B.; Anderson, D.G.; et al. Therapeutic efficacy in a hemophilia B model using a biosynthetic mRNA liver depot system. Gene Ther. 2016, 23, 699–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, L.; Lin, J.; Huang, Y.; Li, L.; Delcassian, D.; Ge, Y.; Shi, Y.; Anderson, D.G. Synergistic lipid compositions for albumin receptor mediated delivery of mRNA to the liver. Nat. Commun. 2020, 11, 2424. [Google Scholar] [CrossRef]
- Raguram, A.; Banskota, S.; Liu, D.R. Therapeutic in vivo delivery of gene editing agents. Cell 2022, 185, 2806–2827. [Google Scholar] [CrossRef]
- Inui, M.; Miyado, M.; Igarashi, M.; Tamano, M.; Kubo, A.; Yamashita, S.; Asahara, H.; Fukami, M.; Takada, S. Rapid generation of mouse models with defined point mutations by the CRISPR/Cas9 system. Sci. Rep. 2014, 4, 5396. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Cheng, Q.; Min, Y.L.; Olson, E.N.; Siegwart, D.J. Systemic nanoparticle delivery of CRISPR-Cas9 ribonucleoproteins for effective tissue specific genome editing. Nat. Commun. 2020, 11, 3232. [Google Scholar] [CrossRef]
- Yin, H.; Song, C.Q.; Suresh, S.; Wu, Q.; Walsh, S.; Rhym, L.H.; Mintzer, E.; Bolukbasi, M.F.; Zhu, L.J.; Kauffman, K.; et al. Structure-guided chemical modification of guide RNA enables potent non-viral in vivo genome editing. Nat. Biotechnol. 2017, 35, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Finn, J.D.; Smith, A.R.; Patel, M.C.; Shaw, L.; Youniss, M.R.; van Heteren, J.; Dirstine, T.; Ciullo, C.; Lescarbeau, R.; Seitzer, J.; et al. A Single Administration of CRISPR/Cas9 Lipid Nanoparticles Achieves Robust and Persistent In Vivo Genome Editing. Cell Rep. 2018, 22, 2227–2235. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yu, W.; Huang, S.; Wu, S.; Li, L.; Zhou, J.; Cao, Y.; Huang, X.; Qiao, Y. Structure-guided engineering of adenine base editor with minimized RNA off-targeting activity. Nat. Commun. 2021, 12, 2287. [Google Scholar] [CrossRef]
- Rothgangl, T.; Dennis, M.K.; Lin, P.J.C.; Oka, R.; Witzigmann, D.; Villiger, L.; Qi, W.; Hruzova, M.; Kissling, L.; Lenggenhager, D.; et al. In vivo adenine base editing of PCSK9 in macaques reduces LDL cholesterol levels. Nat. Biotechnol. 2021, 39, 949–957. [Google Scholar] [CrossRef]
- Musunuru, K.; Chadwick, A.C.; Mizoguchi, T.; Garcia, S.P.; DeNizio, J.E.; Reiss, C.W.; Wang, K.; Iyer, S.; Dutta, C.; Clendaniel, V.; et al. In vivo CRISPR base editing of PCSK9 durably lowers cholesterol in primates. Nature 2021, 593, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Villiger, L.; Rothgangl, T.; Witzigmann, D.; Oka, R.; Lin, P.J.C.; Qi, W.; Janjuha, S.; Berk, C.; Ringnalda, F.; Beattie, M.B.; et al. In vivo cytidine base editing of hepatocytes without detectable off-target mutations in RNA and DNA. Nat. Biomed. Eng. 2021, 5, 179–189. [Google Scholar] [CrossRef]
- Jiang, T.; Henderson, J.M.; Coote, K.; Cheng, Y.; Valley, H.C.; Zhang, X.O.; Wang, Q.; Rhym, L.H.; Cao, Y.; Newby, G.A.; et al. Chemical modifications of adenine base editor mRNA and guide RNA expand its application scope. Nat. Commun. 2020, 11, 1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenkowitz, T.; Kassner, U.; Wühle-Demuth, M.; Salewsky, B.; Rosada, A.; Zemojtel, T.; Hopfenmüller, W.; Isermann, B.; Borucki, K.; Heigl, F.; et al. Clinical characterization and mutation spectrum of German patients with familial hypercholesterolemia. Atherosclerosis 2016, 253, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, T.S.; Lee, A.C.H.; Akinc, A.; Bramlage, B.; Bumcrot, D.; Fedoruk, M.N.; Harborth, J.; Heyes, J.A.; Jeffs, L.B.; John, M.; et al. RNAi-mediated gene silencing in non-human primates. Nature 2006, 441, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, M.; Ansell, S.M.; Mui, B.L.; Tam, Y.K.; Chen, J.; Du, X.; Butler, D.; Eltepu, L.; Matsuda, S.; Narayanannair, J.K.; et al. Maximizing the potency of siRNA lipid nanoparticles for hepatic gene silencing in vivo. Angew. Chem. Int. Ed. Engl. 2012, 51, 8529–8533. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yin, L.; Theisen, M.; Zhuo, J.; Siddiqui, S.; Levy, B.; Presnyak, V.; Frassetto, A.; Milton, J.; Salerno, T.; et al. Systemic mRNA Therapy for the Treatment of Fabry Disease: Preclinical Studies in Wild-Type Mice, Fabry Mouse Model, and Wild-Type Non-human Primates. Am. J. Hum. Genet. 2019, 104, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeRosa, F.; Smith, L.; Shen, Y.; Huang, Y.; Pan, J.; Xie, H.; Yahalom, B.; Heartlein, M.W. Improved Efficacy in a Fabry Disease Model Using a Systemic mRNA Liver Depot System as Compared to Enzyme Replacement Therapy. Mol. Ther. 2019, 27, 878–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardi, N.; Secreto, A.J.; Shan, X.; Debonera, F.; Glover, J.; Yi, Y.; Muramatsu, H.; Ni, H.; Mui, B.L.; Tam, Y.K.; et al. Administration of nucleoside-modified mRNA encoding broadly neutralizing antibody protects humanized mice from HIV-1 challenge. Nat. Commun. 2017, 8, 14630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenjo, E.; Hozumi, H.; Makita, Y.; Iwabuchi, K.A.; Fujimoto, N.; Matsumoto, S.; Kimura, M.; Amano, Y.; Ifuku, M.; Naoe, Y.; et al. Low immunogenicity of LNP allows repeated administrations of CRISPR-Cas9 mRNA into skeletal muscle in mice. Nat. Commun. 2021, 12, 7101. [Google Scholar] [CrossRef]
- Zou, Y.; Sun, X.; Yang, Q.; Zheng, M.; Shimoni, O.; Ruan, W.; Wang, Y.; Zhang, D.; Yin, J.; Huang, X.; et al. Blood-brain barrier-penetrating single CRISPR-Cas9 nanocapsules for effective and safe glioblastoma gene therapy. Sci. Adv. 2022, 8, eabm8011. [Google Scholar] [CrossRef] [PubMed]
- Han, J.P.; Kim, M.J.; Choi, B.S.; Lee, J.H.; Lee, G.S.; Jeong, M.; Lee, Y.; Kim, E.A.; Oh, H.K.; Go, N.; et al. In vivo delivery of CRISPR-Cas9 using lipid nanoparticles enables antithrombin gene editing for sustainable hemophilia A and B therapy. Sci. Adv. 2022, 8, eabj6901. [Google Scholar] [CrossRef]
- Herskovitz, J.; Hasan, M.; Patel, M.; Blomberg, W.R.; Cohen, J.D.; Machhi, J.; Shahjin, F.; Mosley, R.L.; McMillan, J.E.; Kevadiya, B.D.; et al. CRISPR-Cas9 Mediated Exonic Disruption for HIV-1 Elimination. eBioMedicine 2021, 73, 103678. [Google Scholar] [CrossRef]
- Qiu, M.; Glass, Z.; Chen, J.; Haas, M.; Jin, X.; Zhao, X.; Rui, X.; Ye, Z.; Li, Y.; Zhang, F.; et al. Lipid nanoparticle-mediated codelivery of Cas9 mRNA and single-guide RNA achieves liver-specific in vivo genome editing of Angptl3. Proc. Natl. Acad. Sci. USA 2021, 118, e2020401118. [Google Scholar] [CrossRef]
- Rosa, S.S.; Prazeres, D.M.F.; Azevedo, A.M.; Marques, M.P.C. mRNA vaccines manufacturing: Challenges and bottlenecks. Vaccine 2021, 39, 2190–2200. [Google Scholar] [CrossRef]
- Oberli, M.A.; Reichmuth, A.M.; Dorkin, J.R.; Mitchell, M.J.; Fenton, O.S.; Jaklenec, A.; Anderson, D.G.; Langer, R.; Blankschtein, D. Lipid Nanoparticle Assisted mRNA Delivery for Potent Cancer Immunotherapy. Nano Lett. 2017, 17, 1326–1335. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ye, Z.; Huang, C.; Qiu, M.; Song, D.; Li, Y.; Xu, Q. Lipid nanoparticle-mediated lymph node-targeting delivery of mRNA cancer vaccine elicits robust CD8 + T cell response. Proc. Natl. Acad. Sci. USA 2022, 119, e2207841119. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Zhou, K.; Yan, Y.; Miller, J.B.; Siegwart, D.J. Tumor-Activated Water-Soluble Photosensitizers for Near-Infrared Photodynamic Cancer Therapy. ACS Appl. Mater. Interfaces 2018, 10, 16335–16343. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Kos, P.; Yan, Y.; Zhou, K.; Miller, J.B.; Elkassih, S.; Siegwart, D.J. Activatable Water-Soluble Probes Enhance Tumor Imaging by Responding to Dysregulated pH and Exhibiting High Tumor-to-Liver Fluorescence Emission Contrast. Bioconjug. Chem. 2016, 27, 1737–1744. [Google Scholar] [CrossRef]
- Xiong, H.; Liu, S.; Wei, T.; Cheng, Q.; Siegwart, D.J. Theranostic dendrimer-based lipid nanoparticles containing PEGylated BODIPY dyes for tumor imaging and systemic mRNA delivery in vivo. J. Control. Release 2020, 325, 198–205. [Google Scholar] [CrossRef]
- Solomon, S.D.; Adams, D.; Kristen, A.; Grogan, M.; González-Duarte, A.; Maurer, M.S.; Merlini, G.; Damy, T.; Slama, M.S.; Brannagan, T.H.; et al. Effects of Patisiran, an RNA Interference Therapeutic, on Cardiac Parameters in Patients with Hereditary Transthyretin-Mediated Amyloidosis. Circulation 2019, 139, 431–443. [Google Scholar] [CrossRef]
- Mashima, R.; Okuyama, T.; Ohira, M. Biomarkers for Lysosomal Storage Disorders with an Emphasis on Mass Spectrometry. Int. J. Mol. Sci. 2020, 21, 2704. [Google Scholar] [CrossRef] [Green Version]
- Mashima, R.; Okuyama, T.; Ohira, M. Physiology and Pathophysiology of Heparan Sulfate in Animal Models: Its Biosynthesis and Degradation. Int. J. Mol. Sci. 2022, 23, 1963. [Google Scholar] [CrossRef]
- Mashima, R.; Nakanishi, M. Mammalian Sulfatases: Biochemistry, Disease Manifestation, and Therapy. Int. J. Mol. Sci. 2022, 23, 8153. [Google Scholar] [CrossRef]
- Rodríguez-Castejón, J.; Alarcia-Lacalle, A.; Gómez-Aguado, I.; Vicente-Pascual, M.; Aspiazu, M.Á.S.; del Pozo-Rodríguez, A.; Rodríguez-Gascón, A. α-Galactosidase A Augmentation by Non-Viral Gene Therapy: Evaluation in Fabry Disease Mice. Pharmaceutics 2021, 13, 771. [Google Scholar] [CrossRef]
- Gillmore, J.D.; Gane, E.; Taubel, J.; Kao, J.; Fontana, M.; Maitland, M.L.; Seitzer, J.; O’Connell, D.; Walsh, K.R.; Wood, K.; et al. CRISPR-Cas9 In Vivo Gene Editing for Transthyretin Amyloidosis. N. Engl. J. Med. 2021, 385, 493–502. [Google Scholar] [CrossRef]
- Adams, D.; Suhr, O.B.; Dyck, P.J.; Litchy, W.J.; Leahy, R.G.; Chen, J.; Gollob, J.; Coelho, T. Trial design and rationale for APOLLO, a Phase 3, placebo-controlled study of patisiran in patients with hereditary ATTR amyloidosis with polyneuropathy. BMC Neurol. 2017, 17, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamisawa, M.; Claggett, B.; Adams, D.; Kristen, A.v.; Merlini, G.; Slama, M.S.; Dispenzieri, A.; Shah, A.M.; Falk, R.H.; Karsten, V.; et al. Association of Patisiran, an RNA Interference Therapeutic, with Regional Left Ventricular Myocardial Strain in Hereditary Transthyretin Amyloidosis: The APOLLO Study. JAMA Cardiol. 2019, 4, 466–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obici, L.; Berk, J.L.; González-Duarte, A.; Coelho, T.; Gillmore, J.; Schmidt, H.H.J.; Schilling, M.; Yamashita, T.; Labeyrie, C.; Brannagan, T.H.; et al. Quality of life outcomes in APOLLO, the phase 3 trial of the RNAi therapeutic patisiran in patients with hereditary transthyretin-mediated amyloidosis. Amyloid 2020, 27, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Goel, V.; Attarwala, H.; Sweetser, M.T.; Clausen, V.A.; Robbie, G.J. Patisiran Pharmacokinetics, Pharmacodynamics, and Exposure-Response Analyses in the Phase 3 APOLLO Trial in Patients with Hereditary Transthyretin-Mediated (hATTR) Amyloidosis. J. Clin. Pharmacol. 2020, 60, 37–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srikrishna, D.; Sachsenmeier, K. We need to bring R0. Genome Med. 2021, 13, 120. [Google Scholar] [CrossRef]
- Dilliard, S.A.; Cheng, Q.; Siegwart, D.J. On the mechanism of tissue-specific mRNA delivery by selective organ targeting nanoparticles. Proc. Natl. Acad. Sci. USA 2021, 118, e2109256118. [Google Scholar] [CrossRef]
- Cheng, Q.; Wei, T.; Farbiak, L.; Johnson, L.T.; Dilliard, S.A.; Siegwart, D.J. Selective organ targeting (SORT) nanoparticles for tissue-specific mRNA delivery and CRISPR-Cas gene editing. Nat. Nanotechnol. 2020, 15, 313–320. [Google Scholar] [CrossRef]
- Liu, S.; Cheng, Q.; Wei, T.; Yu, X.; Johnson, L.T.; Farbiak, L.; Siegwart, D.J. Membrane-destabilizing ionizable phospholipids for organ-selective mRNA delivery and CRISPR-Cas gene editing. Nat. Mater. 2021, 20, 701–710. [Google Scholar] [CrossRef]
- Sago, C.D.; Lokugamage, M.P.; Paunovska, K.; Vanover, D.A.; Monaco, C.M.; Shah, N.N.; Castro, M.G.; Anderson, S.E.; Rudoltz, T.G.; Lando, G.N.; et al. High-throughput in vivo screen of functional mRNA delivery identifies nanoparticles for endothelial cell gene editing. Proc. Natl. Acad. Sci. USA 2018, 115, E9944–E9952. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, L.; Xiong, H.; Miller, J.B.; Zhou, K.; Kos, P.; Huffman, K.E.; Elkassih, S.; Norman, J.W.; Carstens, R.; et al. Functional polyesters enable selective siRNA delivery to lung cancer over matched normal cells. Proc. Natl. Acad. Sci. USA 2016, 113, E5702–E5710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Liu, S.; Cheng, Q.; Lee, S.M.; Wei, T.; Wang, X.; Siegwart, D.J.; Zhang, D.; Farbiak, L.; Johnson, L.T. Hydrophobic Optimization of Functional Poly(TPAE-co-suberoyl chloride) for Extrahepatic mRNA Delivery following Intravenous Administration. Pharmaceutics 2021, 13, 1914. [Google Scholar] [CrossRef] [PubMed]
- Rurik, J.G.; Tombácz, I.; Yadegari, A.; Fernández, P.O.M.; Shewale, S.V.; Li, L.; Kimura, T.; Soliman, O.Y.; Papp, T.E.; Tam, Y.K.; et al. CAR T cells produced in vivo to treat cardiac injury. Science 2022, 375, 91–96. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Product | Lipid Composition | Ref |
|---|---|---|
| Cationic ionizable lipid/helper phospholipid/cholesterol/PEG-lipid | ||
| Onpattro | DLin-MC3-DMA/DSPC/Chol/DMG-PEG2000 = 50/10/38.5/1.5 | [5] |
| mRNA-1273 | SM-102/DSPC/Chol/DMG-PEG2000 = 50/10/38.5/1.5 | [6] |
| BNT126b2 | ALC-0315/DSPC/Chol/ALC-0159 = 46.3/9.4/42.7/1.6 | [7] |
| mRNA | sgRNA | |
|---|---|---|
| Preparation | In vitro transcription | Solid phase synthesis |
| Modification | ||
| 5′-Capping | Enzymatic | Not applicable |
| 2′-O-methylation | Enzymatic | Synthetic |
| 3′/5′-O-phosphorothioation | Not applicable | Synthetic |
| Replacement by uridine analog | Enzymatic | Not applicable |
| Codon optimization | In silico | Not applicable |
| Nomenclature | 5′-5′ Bridge | Base | Enzyme | |||
|---|---|---|---|---|---|---|
| Non-Transcribed Nucleoside | 1st Transcribed Nucleoside | 2nd Transcribed Nucleoside | ||||
| Guanine | Ribose 2′- | Adenine | Ribose 2′- | |||
| Uncapped | GpppA/G | No modification | OH | H | OH | |
| Cap 0 | GpppA/G | N7-Me | OH | H | OH | TGS |
| Cap 1 | GpppA/G | N7-Me | Me | H | OH | CMTR1 |
| Cap 2 | GpppA/G | N7-Me | Me | H | Me | CMTR2 |
| GpppG | N7-Me; N2-Me; N2,2′-di-Me | OH | H | OH | TGS | |
| GpppA | N7-Me | OH | N2-Me | OH | SAMT | |
| GpppA | N7-Me | Me | N6-Me | Me | CAPAM | |
| Disorders | Target | Genetic Material | Ref |
|---|---|---|---|
| Bleeding disorder | FVII | siRNA | [41] |
| Hypercholesterolemia | Pcsk9 | siRNA | [23] |
| Hypercholesterolemia | ApoB | siRNA | [40] |
| Transthyretin-mediated amyloidosis | Ttr | siRNA | [24] |
| Transthyretin-mediated amyloidosis | Ttr | siRNA | [23] |
| Anemia | mEPO, pEPO | Therapeutic mRNA | [26] |
| Anemia | hEPO | Therapeutic mRNA | [27] |
| Anemia | hEPO | Therapeutic mRNA | [28] |
| Fabry disease | hGLA | Therapeutic mRNA | [42] |
| Fabry disease | hGLA | Therapeutic mRNA | [43] |
| HIV | anti-HIV-1 antibody | Therapeutic mRNA | [44] |
| DMD | Human DMD exon 45 | Cas9 | [45] |
| Glioblastoma | Plk1 | Cas9 | [46] |
| Hemophilia A and B | Serpinc1 (encoding antithrombin) | Cas9 | [47] |
| HIV | TatDE | Cas9 | [48] |
| Hypercholesterolemia | Angptl3 | Cas9 | [49] |
| Hypercholesterolemia | Pcsk9 | Cas9 | [24] |
| Hypercholesterolemia | Pcsk9 | Cas9 | [31] |
| Hypercholesterolemia | Pcsk9 | Cas9 | [32] |
| Transthyretin-mediated amyloidosis | Ttr | Cas9 | [33] |
| Hypercholesterolemia | Pcsk9 | ABE | [36] |
| Hypercholesterolemia | Pcsk9 | ABE | [35] |
| Phenylketonuria | Pah | CBE | [37] |
| Tyrosinemia I | Fah | ABE | [38] |
| NCT Number | Conditions-1 | Conditions-2 | Target | Payload | Phases | Status | Sponsor/Collaborators | Ref |
|---|---|---|---|---|---|---|---|---|
| NCT04601051 | Amyloidosis | hATTR | Transthyretin | Cas9 mRNA | Phase 1 | Recruiting | Intellia Therapeutics | [61] |
| NCT01960348 | Amyloidosis | hATTR | Transthyretin | siRNA | Phase 3 | Completed | Alnylam Pharmaceuticals | [5,56,62,63,64,65] |
| NCT01437007 | Cancer | Hepatic Metastases | PLK1 | siRNA | Phase 1 | Completed | NCI; NIH Clinical Center | |
| NCT03323398 | Cancer | Solid Tumor | Human OX40L | mRNA | Phase 1/2 | Terminated | ModernaTX, Inc. | |
| NCT03739931 | Cancer | Solid Tumor | Human OX40L, IL-23, and IL-36γ | mRNA | Phase 1 | Recruiting | ModernaTX, Inc.; AstraZeneca | [66] |
| NCT04675996 | Cancer | Solid Tumor | miR-193a-3p | miRNA | Phase 1 | Recruiting | InteRNA | |
| NCT02110563 | Cancer | Solid Tumor | MYC | siRNA | Phase 1 | Terminated | Dicerna Pharmaceuticals, Inc. | |
| NCT02314052 | Cancer | Hepatocellular Carcinoma | MYC | siRNA | Phase 1/2 | Terminated | Dicerna Pharmaceuticals, Inc. | |
| NCT04486833 | Cancer | Carcinoma, Non-Small Cell Lung | TUSC2 | DNA plasmid | Phase 1/2 | Recruiting | Genprex, Inc. | |
| NCT05062980 | Cancer | Non-Small Cell Lung Cancer | TUSC2 | DNA plasmid | Phase 1/2 | Recruiting | Genprex, Inc. | |
| NCT05497453 | Cancer | Solid Tumor | 2 independent epigenomic controllers | biscistronic mRNA | Phase 1/2 | Recruiting | Omega Therapeutics | |
| NCT02227459 | Hepatic Fibrosis | HSP47 | siRNA | Phase 1 | Completed | Bristol-Myers Squibb; Nitto Denko Corporation | ||
| NCT04416126 | OTCD | Ornithine transcarbamylase | Therapeutic mRNA | Phase 1 | Completed | Arcturus Therapeutics, Inc. | ||
| NCT04442347 | OTCD | Ornithine transcarbamylase | Therapeutic mRNA | Phase 1 | Recruiting | Arcturus Therapeutics, Inc. | ||
| NCT05526066 | OTCD | Ornithine transcarbamylase | Therapeutic mRNA | Phase 2 | Recruiting | Arcturus Therapeutics, Inc. |
| Target Organ | Basic Component | Additive | Ref |
|---|---|---|---|
| Cationic ionizable lipid/helper phospholipid/cholesterol/PEG-lipid | (SORT molecule) | ||
| Conventional | LP01/DSPC/Chol/PEG2000-DMG = 45/9/44/2 | Not added | [33] |
| DLin-MC3-DMA/DSPC/Chol/DMG-PEG2000 = 50/10/38.5/1.5 | Not added | [69] | |
| 5A2-SC8/DOPE/Chol/C14PEG2000 = 23.8/23.8/47.8/4.8 | Not added | [67] | |
| Liver | 5A2-SC8/DOPE/Chol/C14PEG2000 = 19/19/38.4/4 | DODAP = 20 | [67,68] |
| 9A1P9/5A2-SC8/Chol/DMG-PEG2000 = 25/30/30/1 | Not added | [69] | |
| Spleen | 5A2-SC8/DOPE/Chol/C14PEG2000 = 16.7/16.7/33.3/3.3 | 18:1 PA = 30 | [67,68] |
| Lung | 5A2-SC8/DOPE/Chol/C14PEG2000 = 11.9/11.9/23.8/2.4 | DOTAP = 50 | [67,68] |
| 9A1P9/DDAB/Chol/DMG-PEG2000 = 60/30/40/0.4 | Not added | [69] | |
| 5A2-SC8/DOPE/Chol/DMG-PEG = 15/15/30/3 | DOTAP = 63 | [31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mashima, R.; Takada, S. Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application. Curr. Issues Mol. Biol. 2022, 44, 5013-5027. https://doi.org/10.3390/cimb44100341
Mashima R, Takada S. Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application. Current Issues in Molecular Biology. 2022; 44(10):5013-5027. https://doi.org/10.3390/cimb44100341
Chicago/Turabian StyleMashima, Ryuichi, and Shuji Takada. 2022. "Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application" Current Issues in Molecular Biology 44, no. 10: 5013-5027. https://doi.org/10.3390/cimb44100341
APA StyleMashima, R., & Takada, S. (2022). Lipid Nanoparticles: A Novel Gene Delivery Technique for Clinical Application. Current Issues in Molecular Biology, 44(10), 5013-5027. https://doi.org/10.3390/cimb44100341