
Reduced Tyrosine and Serine-632 Phosphorylation of Insulin Receptor Substrate-1 in the Gastrocnemius Muscle of Obese Zucker Rat


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Glucose Tolerance Test (GTT)
2.3. Protein Extraction and Immunoprecipitation
2.4. Immunoblotting Analysis of IRS1, IRβ, and Tyrosine and Serine Phosphorylation Levels in the Gastrocnemius Muscle of Zucker Lean and Obese Rats
2.5. Statistics
3. Results
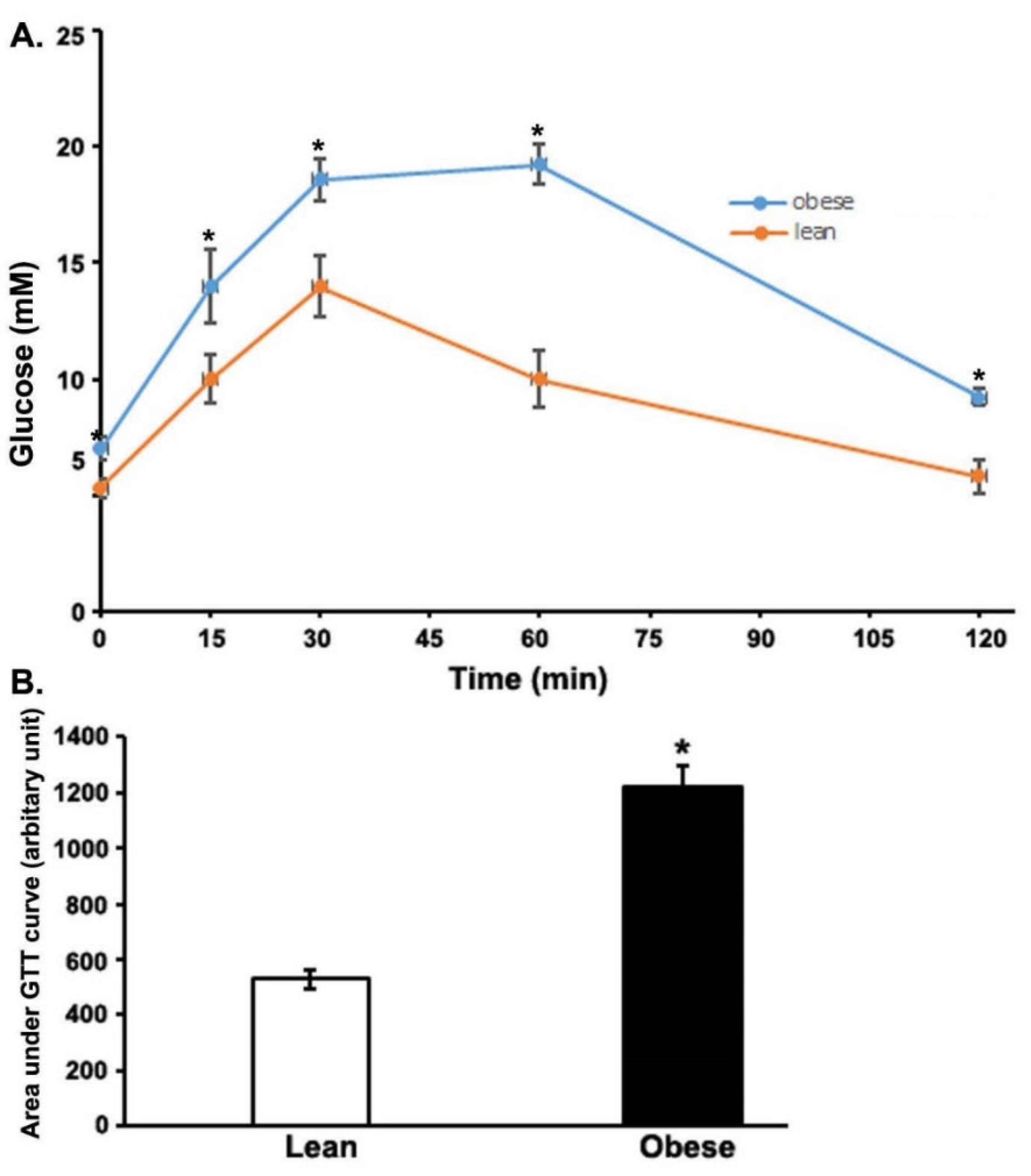
3.1. Impaired Glucose Tolerance Test (GTT) in the Obese Zucker Rats (OZRs)
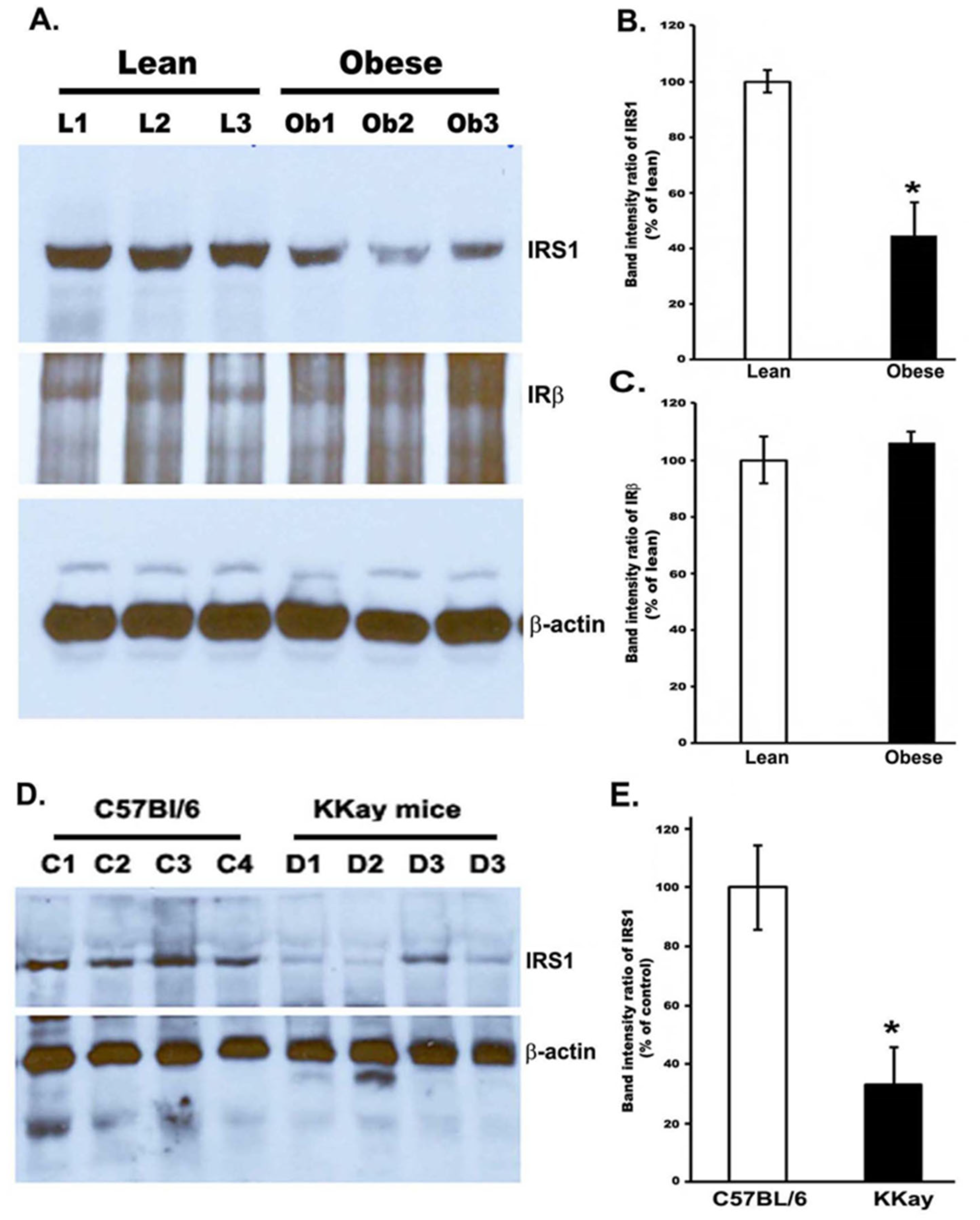
3.2. Reduced IRS-1 Level in the Gastrocnemius Muscle of Obese Zucker Rats and Non-Obese Insulin Resistance in KKay Mice
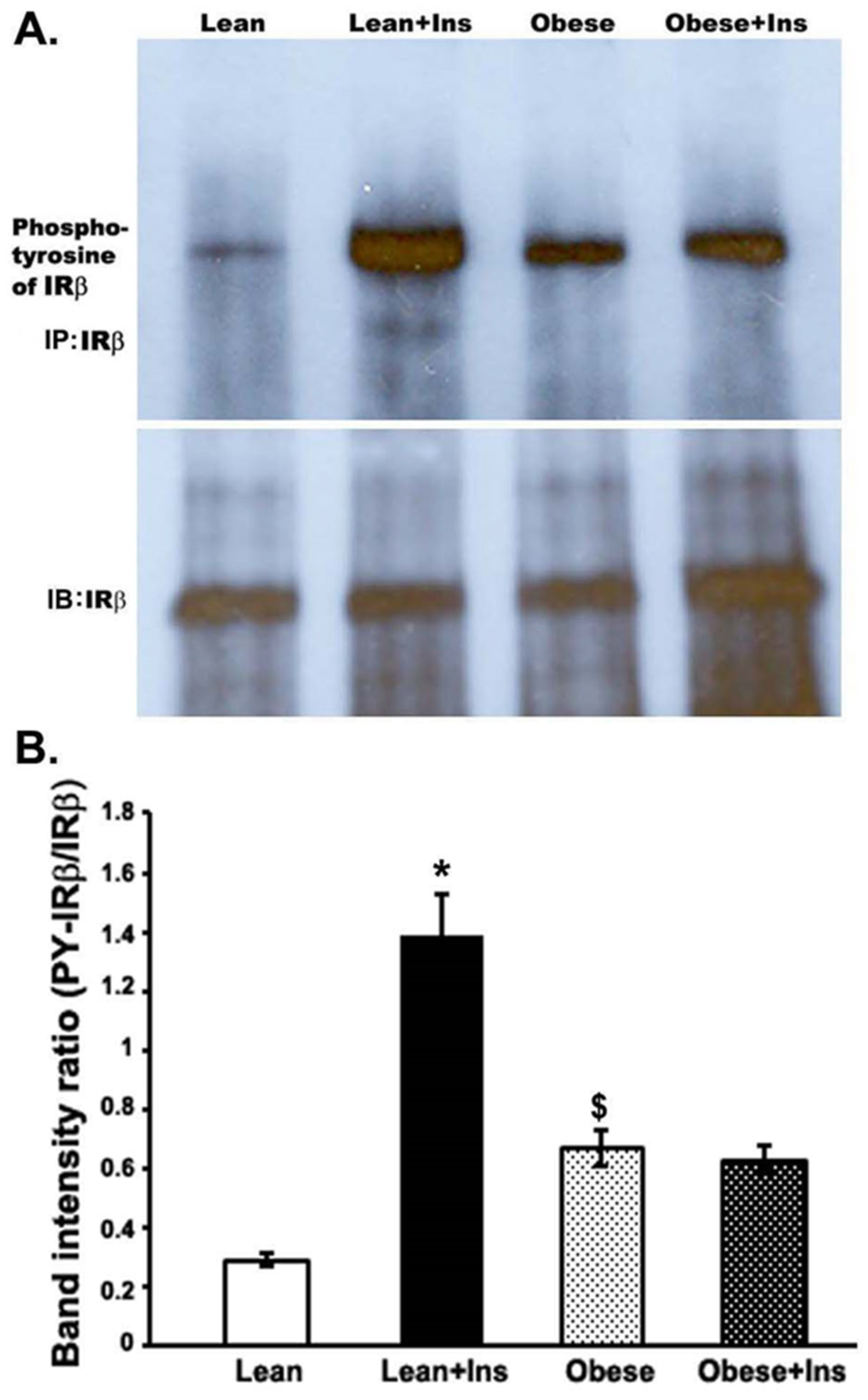
3.3. Altered Insulin-Stimulated Phosphotyrosine of IRβ (pY-IRβ) in the Gastrocnemius Muscle of Obese Zucker Rats
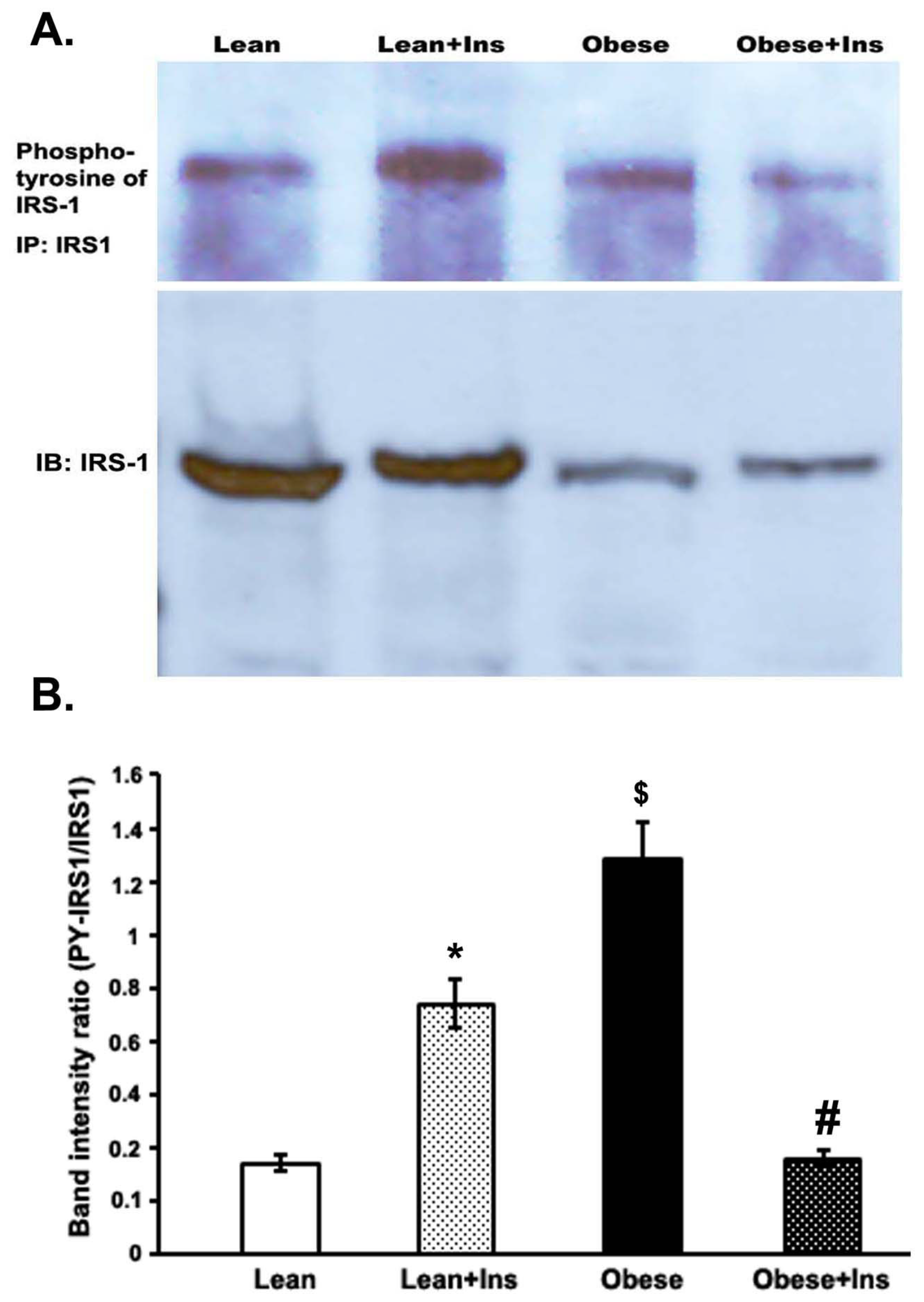
3.4. Altered Insulin-Stimulated Tyrosine Phosphorylation of IRS-1 (pY-IRS-1) in the Gastrocnemius Muscle of Obese Zucker Rats
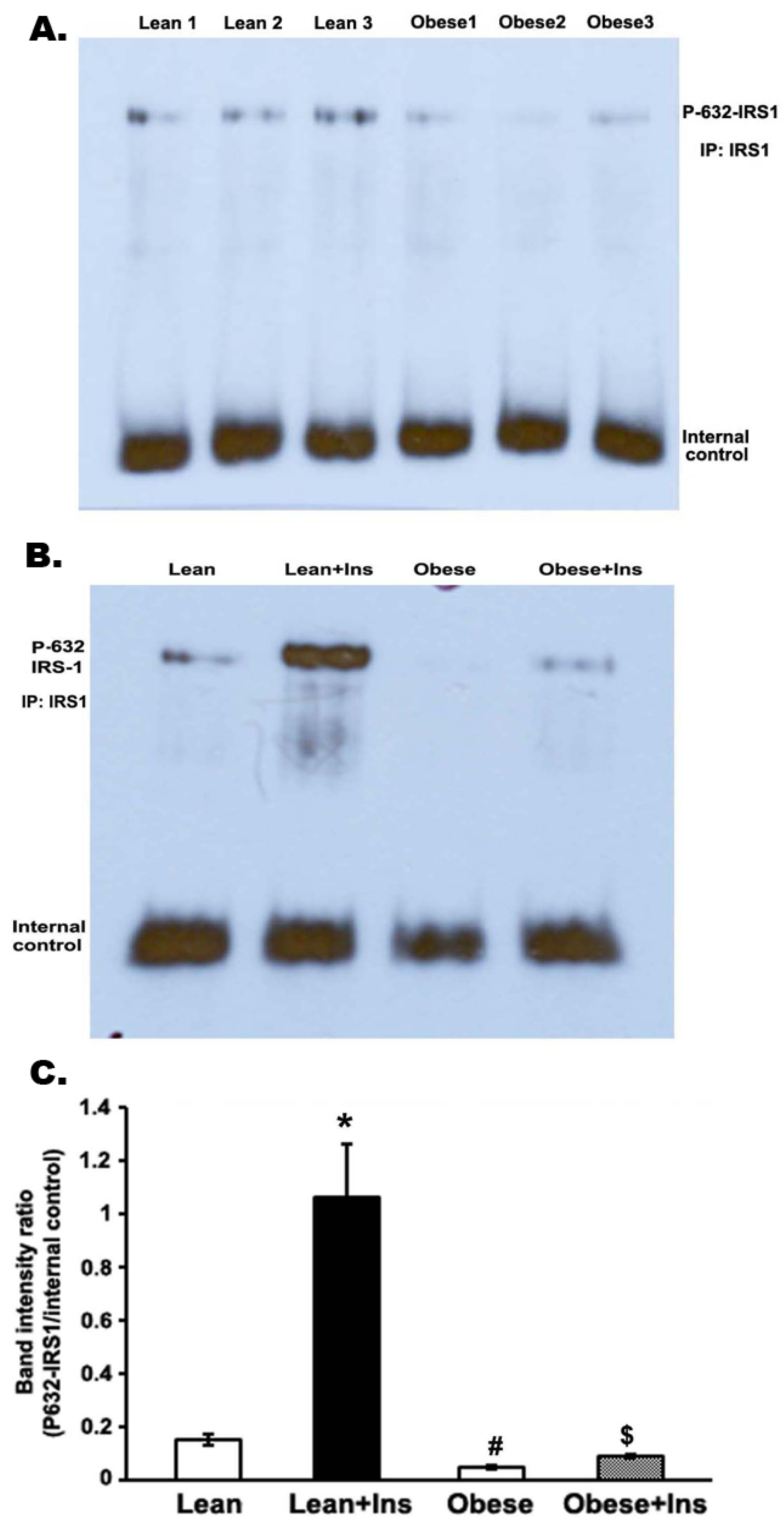
3.5. Insulin-Stimulated Decrease in Serine Phosphorylation of IRS-1 (Serine 632-IRS-1) in the Gastrocnemius Muscle of Obese Zucker Rats
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qi, Y.; Zhang, X.; Seyoum, B.; Msallaty, Z.; Mallisho, A.; Caruso, M.; Damacharla, D.; Ma, D.; Al-Janabi, W.; Tagett, R.; et al. Kinome Profiling Reveals Abnormal Activity of Kinases in Skeletal Muscle from Adults with Obesity and Insulin Resistance. J. Clin. Endocrinol. Metab. 2020, 105, 644–659. [Google Scholar] [CrossRef] [PubMed]
- Snell-Bergeon, J.K.; Chartier-Logan, C.; Maahs, D.M.; Ogden, L.G.; Hokanson, J.E.; Kinney, G.L.; Eckel, R.H.; Ehrlich, J.; Rewers, M. Adults with type 1 diabetes eat a high-fat atherogenic diet that is associated with coronary artery calcium. Diabetologia 2009, 52, 801–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylow, L.; Tokarz, V.L.; Richter, E.A.; Klip, A. The many actions of insulin in skeletal muscle, the paramount tissue determining glycemia. Cell Metab. 2021, 33, 758–780. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Chang, H.; La, X.; Li, J.-A. Wushenziye Formula Improves Skeletal Muscle Insulin Resistance in Type 2 Diabetes Mellitus via PTP1B-IRS1-Akt-GLUT4 Signaling Pathway. Evid.-Based Complement. Altern. Med. 2017, 2017, 4393529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, R.; Bryant, N.J.; Boyle, J.G.; Petrie, J.R.; Gould, G.W. Diabetes is accompanied by changes in the levels of proteins involved in endosomal GLUT4 trafficking in obese human skeletal muscle. Endocrinol. Diabetes Metab. 2022, 5, e361. [Google Scholar] [CrossRef]
- Shepherd, P.R.; Kahn, B.B. Glucose transporters and insulin action—Implications for insulin resistance and diabetes mellitus. N. Engl. J. Med. 1999, 341, 248–257. [Google Scholar] [CrossRef]
- Virkamäki, A.; Ueki, K.; Kahn, C.R. Protein-protein interaction in insulin signaling and the molecular mechanisms of insulin resistance. J. Clin. Investig. 1999, 103, 931–943. [Google Scholar] [CrossRef] [Green Version]
- Gual, P.; le Marchand-Brustel, Y.; Tanti, J.F. Positive and negative regulation of insulin signaling through IRS-1 phosphorylation. Biochimie 2005, 87, 99–109. [Google Scholar] [CrossRef]
- Gual, P.; Grémeaux, T.; Gonzalez, T.; Le Marchand-Brustel, Y.; Tanti, J.-F. MAP kinases and mTOR mediate insulin-induced phosphorylation of insulin receptor substrate-1 on serine residues 307, 612 and 632. Diabetologia 2003, 46, 1532–1542. [Google Scholar] [CrossRef]
- Rui, L.; Aguirre, V.; Kim, J.K.; Shulman, G.I.; Lee, A.; Corbould, A.; Dunaif, A.; White, M.F. Insulin/IGF-1 and TNF-alpha stimulate phosphorylation of IRS-1 at inhibitory Ser307 via distinct pathways. J. Clin. Investig. 2001, 107, 181–189. [Google Scholar] [CrossRef]
- Paz, K.; Liu, Y.-F.; Shorer, H.; Hemi, R.; LeRoith, D.; Quan, M.; Kanety, H.; Seger, R.; Zick, Y. Phosphorylation of insulin receptor substrate-1 (IRS-1) by protein kinase B positively regulates IRS-1 function. J. Biol. Chem. 1999, 274, 28816–28822. [Google Scholar] [CrossRef] [Green Version]
- Phillips, M.S.; Liu, Q.; Hammond, H.A.; Dugan, V.; Hey, P.J.; Caskey, C.T.; Hess, J.F. Leptin receptor missense mutation in the fatty Zucker rat. Nat. Genet. 1996, 13, 18–19. [Google Scholar] [CrossRef]
- Iida, M.; Murakami, T.; Ishida, K.; Mizuno, A.; Kuwajima, M.; Shima, K. Phenotype-linked amino acid alteration in leptin receptor cDNA from Zucker fatty (fa/fa) rat. Biochem. Biophys. Res. Commun. 1996, 222, 19–26. [Google Scholar] [CrossRef]
- Suto, J.; Matsuura, S.; Imamura, K.; Yamanaka, H.; Sekikawa, K. Genetic analysis of non-insulin-dependent diabetes mellitus in KK and KK-Ay mice. Eur. J. Endcrinol. 1998, 139, 654–661. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.E.; Dorsey, D.A.; Beaudet, L.N.; Peterson, R.G. Analysis of the Zucker Diabetic Fatty (ZDF) type 2 diabetic rat model suggests a neurotrophic role for insulin/IGF-I in diabetic autonomic neuropathy. Am. J. Pathol. 2003, 163, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Terrettaz, J.; Jeanrenaud, B. In Vivo Hepatic and Peripheral Insulin Resistance in Genetically Obese (fa/fa) Rats. Endocrinology 1983, 112, 1346–1351. [Google Scholar] [CrossRef]
- Hurrell, D.G.; Pedersen, O.; Kahn, C.R. Alterations in the hepatic insulin receptor kinase in genetic and acquired obesity in rats. Endocrinology 1989, 125, 2454–2462. [Google Scholar] [CrossRef]
- Kasiske, B.L.; O’Donnell, M.P.; Keane, W.F. The Zucker rat model of obesity, insulin resistance, hyperlipidemia, and renal injury. Hypertension 1992, 19 (Suppl. S1), I110–I115. [Google Scholar] [CrossRef]
- Crist, G.H. Effects of adenosine receptor antagonism on protein tyrosine phosphatase in rat skeletal muscle. Int. J. Biochem. Cell Biol. 2001, 33, 817–830. [Google Scholar] [CrossRef]
- Ola, M.S.; Hosoya, K.-I.; LaNoue, K.F. Influence of insulin on glutamine synthetase in the Müller glial cells of retina. Metab. Brain Dis. 2011, 26, 195–202. [Google Scholar] [CrossRef]
- Ola, M.S. Effect of hyperglycemia on insulin receptor signaling in the cultured retinal Müller glial cells. Biochem. Biophys. Res. Commun. 2014, 444, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Shulman, G.I. Cellular mechanisms of insulin resistance. J. Clin. Investig. 2000, 106, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb. Perspect. Biol. 2014, 6, a009191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeFronzo, R.A.; Gunnarsson, R.; Björkman, O.; Olsson, M.; Wahren, J. Effects of insulin on peripheral and splanchnic glucose metabolism in noninsulin-dependent (type II) diabetes mellitus. J. Clin. Investig. 1985, 76, 149–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moroki, T.; Yoshikawa, Y.; Yoshizawa, K.; Tsubura, A.; Yasui, H. Morphological characterization of systemic changes in KK-Ay mice as an animal model of type 2 diabetes. In Vivo 2013, 27, 465–472. [Google Scholar]
- Balazova, L.; Krskova, K.; Suski, M.; Sisovsky, V.; Hlavacova, N.; Olszanecki, R.; Jezova, D.; Zorad, S. Metabolic effects of subchronic peripheral oxytocin administration in lean and obese zucker rats. J. Physiol. Pharmacol. 2016, 67, 531–541. [Google Scholar]
- Anai, M.; Funaki, M.; Ogihara, T.; Terasaki, J.; Inukai, K.; Katagiri, H.; Fukushima, Y.; Yazaki, Y.; Kikuchi, M.; Oka, Y.; et al. Altered expression levels and impaired steps in the pathway to phosphatidylinositol 3-kinase activation via insulin receptor substrates 1 and 2 in Zucker fatty rats. Diabetes 1998, 47, 13–23. [Google Scholar] [CrossRef]
- Hevener, A.L.; Reichart, D.; Olefsky, J. Exercise and thiazolidinedione therapy normalize insulin action in the obese Zucker fatty rat. Diabetes 2000, 49, 2154–2159. [Google Scholar] [CrossRef] [Green Version]
- Kerouz, N.J.; Hörsch, D.; Pons, S.; Kahn, C.R. Differential regulation of insulin receptor substrates-1 and -2 (IRS-1 and IRS-2) and phosphatidylinositol 3-kinase isoforms in liver and muscle of the obese diabetic (ob/ob) mouse. J. Clin. Investig. 1997, 100, 3164–3172. [Google Scholar] [CrossRef]
- Sesti, G.; Federici, M.; Hribal, M.L.; Lauro, D.; Sbraccia, P.; Lauro, R. Defects of the insulin receptor substrate (IRS) system in human metabolic disorders. FASEB J. 2001, 15, 2099–2111. [Google Scholar] [CrossRef] [Green Version]
- Goodyear, L.J.; Giorgino, F.; Sherman, L.A.; Carey, J.; Smith, R.J.; Dohm, G.L. Insulin receptor phosphorylation, insulin receptor substrate-1 phosphorylation, and phosphatidylinositol 3-kinase activity are decreased in intact skeletal muscle strips from obese subjects. J. Clin. Investig. 1995, 95, 2195–2204. [Google Scholar] [CrossRef]
- Carvalho, E.; Jansson, P.; Axelsen, M.; Eriksson, J.W.; Huang, X.; Groop, L.; Rondinone, C.; Sjöström, L.; Smith, U. Low cellular IRS 1 gene and protein expression predict insulin resistance and NIDDM. FASEB J. 1999, 13, 2173–2178. [Google Scholar] [CrossRef]
- Shah, O.J.; Wang, Z.; Hunter, T. Inappropriate activation of the TSC/Rheb/mTOR/S6K cassette induces IRS1/2 depletion, insulin resistance, and cell survival deficiencies. Curr. Biol. 2004, 14, 1650–1656. [Google Scholar] [CrossRef] [Green Version]
- Christ, C.Y.; Hunt, D.; Hancock, J.; Garcia-Macedo, R.; Mandarino, L.J.; Ivy, J.L. Exercise training improves muscle insulin resistance but not insulin receptor signaling in obese Zucker rats. J. Appl. Physiol. 2002, 92, 736–744. [Google Scholar] [CrossRef] [Green Version]
- Bouzakri, K.; Roques, M.; Gual, P.; Espinosa, S.; Guebre-Egziabher, F.; Riou, J.-P.; Laville, M.; Le Marchand-Brustel, Y.; Tanti, J.-F.; Vidal, H. Reduced activation of phosphatidylinositol-3 kinase and increased serine 636 phosphorylation of insulin receptor substrate-1 in primary culture of skeletal muscle cells from patients with type 2 diabetes. Diabetes 2003, 52, 1319–1325. [Google Scholar] [CrossRef] [Green Version]
- Hotamisligil, G.S.; Budavari, A.; Murray, D.; Spiegelman, B.M. Reduced tyrosine kinase activity of the insulin receptor in obesity-diabetes. Central role of tumor necrosis factor-alpha. J. Clin. Investig. 1994, 94, 1543–1549. [Google Scholar] [CrossRef]
- Friedman, J.E.; Ishizuka, T.; Liu, S.; Farrell, C.J.; Bedol, D.; Koletsky, R.J.; Kaung, H.-L.; Ernsberger, P. Reduced insulin receptor signaling in the obese spontaneously hypertensive Koletsky rat. Am. J. Physiol. 1997, 273, E1014–E1023. [Google Scholar] [CrossRef]
- Youngren, J.F.; Paik, J.; Barnard, R.J. Impaired insulin-receptor autophosphorylation is an early defect in fat-fed, insulin-resistant rats. J. Appl. Physiol. 2001, 91, 2240–2247. [Google Scholar] [CrossRef] [Green Version]
- Fujii, N.; Ho, R.C.; Manabe, Y.; Jessen, N.; Toyoda, T.; Holland, W.L.; Summers, S.A.; Hirshman, M.F.; Goodyear, L.J. Ablation of AMP-activated protein kinase alpha2 activity exacerbates insulin resistance induced by high-fat feeding of mice. Diabetes 2008, 57, 2958–2966. [Google Scholar] [CrossRef] [Green Version]
- Zolotnik, I.A.; Figueroa, T.Y.; Yaspelkis, B.B., 3rd. Insulin receptor and IRS-1 co-immunoprecipitation with SOCS-3, and IKKα/β phosphorylation are increased in obese Zucker rat skeletal muscle. Life Sci. 2012, 91, 816–822. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.Y.; Lin, Y.-W.; Clemont, A.; Feener, E.P.; Hein, K.D.; Igarashi, M.; Yamauchi, T.; White, M.F.; King, G.L. Characterization of selective resistance to insulin signaling in the vasculature of obese Zucker (fa/fa) rats. J. Clin. Investig. 1999, 104, 447–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Dallas-Yang, Q.; Li, Z.; Szalkowski, D.; Liu, F.; Shen, X.; Wu, M.; Zhou, G.; Doebber, T.; Berger, J.; et al. Potentiation of insulin signaling in tissues of Zucker obese rats after acute and long-term treatment with PPARgamma agonists. Diabetes 2002, 51, 2412–2419. [Google Scholar] [CrossRef] [PubMed]
- Slieker, L.J.; Roberts, E.F.; Shaw, W.N.; Johnson, W.T. Effect of streptozocin-induced diabetes on insulin-receptor tyrosine kinase activity in obese Zucker rats. Diabetes 1990, 39, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Dolan, P.L.; Dohm, G.L. Dephosphorylation increases insulin-stimulated receptor kinase activity in skeletal muscle of obese Zucker rats. Mol. Cell. Biochem. 1999, 194, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.; Araki, E.; Miralpeix, M.; Rothenberg, P.L.; White, M.F.; Kahn, C.R. Regulation of insulin receptor substrate-1 in liver and muscle of animal models of insulin resistance. J. Clin. Investig. 1992, 90, 1839–1849. [Google Scholar] [CrossRef] [Green Version]
- Zick, Y. Insulin resistance: A phosphorylation-based uncoupling of insulin signaling. Trends Cell Biol. 2001, 11, 437–441. [Google Scholar] [CrossRef]
- Greene, M.W.; Garofalo, R.S. Positive and negative regulatory role of insulin receptor substrate 1 and 2 (IRS-1 and IRS-2) serine/threonine phosphorylation. Biochemistry 2002, 41, 7082–7091. [Google Scholar] [CrossRef]
- Langlais, P.; Yi, Z.; Finlayson, J.; Luo, M.; Mapes, R.; De Filippis, E.; Meyer, C.; Plummer, E.; Tongchinsub, P.; Mattern, M.; et al. Global IRS-1 phosphorylation analysis in insulin resistance. Diabetologia 2011, 54, 2878–2889. [Google Scholar] [CrossRef] [Green Version]
- Morino, K.; Petersen, K.F.; Dufour, S.; Befroy, D.; Frattini, J.; Shatzkes, N.; Neschen, S.; White, M.F.; Bilz, S.; Sono, S.; et al. Reduced mitochondrial density and increased IRS-1 serine phosphorylation in muscle of insulin-resistant offspring of type 2 diabetic parents. J. Clin. Investig. 2005, 115, 3587–3593. [Google Scholar] [CrossRef] [Green Version]
- Stuart, C.A.; Howell, M.E.A.; Cartwright, B.M.; McCurry, M.P.; Lee, M.L.; Ramsey, M.W.; Stone, M.H. Insulin resistance and muscle insulin receptor substrate-1 serine hyperphosphorylation. Physiol. Rep. 2014, 2, e12236. [Google Scholar] [CrossRef]
- Furukawa, N.; Ongusaha, P.; Jahng, W.J.; Araki, K.; Choi, C.S.; Kim, H.-J.; Lee, Y.H.; Kaibuchi, K.; Kahn, B.B.; Masuzaki, H.; et al. Role of Rho-kinase in regulation of insulin action and glucose homeostasis. Cell Metab. 2005, 2, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Mothe, I.; Van Obberghen, E. Phosphorylation of insulin receptor substrate-1 on multiple serine residues, 612, 632, 662, and 731, modulates insulin action. J. Biol. Chem. 1996, 271, 11222–11227. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ola, M.S. Reduced Tyrosine and Serine-632 Phosphorylation of Insulin Receptor Substrate-1 in the Gastrocnemius Muscle of Obese Zucker Rat. Curr. Issues Mol. Biol. 2022, 44, 6015-6027. https://doi.org/10.3390/cimb44120410
Ola MS. Reduced Tyrosine and Serine-632 Phosphorylation of Insulin Receptor Substrate-1 in the Gastrocnemius Muscle of Obese Zucker Rat. Current Issues in Molecular Biology. 2022; 44(12):6015-6027. https://doi.org/10.3390/cimb44120410
Chicago/Turabian StyleOla, Mohammad Shamsul. 2022. "Reduced Tyrosine and Serine-632 Phosphorylation of Insulin Receptor Substrate-1 in the Gastrocnemius Muscle of Obese Zucker Rat" Current Issues in Molecular Biology 44, no. 12: 6015-6027. https://doi.org/10.3390/cimb44120410
APA StyleOla, M. S. (2022). Reduced Tyrosine and Serine-632 Phosphorylation of Insulin Receptor Substrate-1 in the Gastrocnemius Muscle of Obese Zucker Rat. Current Issues in Molecular Biology, 44(12), 6015-6027. https://doi.org/10.3390/cimb44120410