1. Introduction
Pecans are an important crop in the southeastern United States and are increasing in importance in other countries [
1]. The U.S. produced 302 million pounds of in-shell pecans in 2020, and the state of Georgia is the second leading producer of pecans in the U.S., producing 64 million kilograms in 2020 [
2]. Pecan scab is caused by the plant pathogenic fungus
Venturia effusa, which thrives in conditions of high temperatures, high humidity, and frequent rainfall, which are common to the southeastern United States [
3,
4]. This disease is polycyclic, having multiple generations per year, and overwinters as lesions on twigs and shucks within the canopy of the tree. The lesions sporulate in the spring, and conidia are dispersed down through the canopy by rainfall, infecting the growing fruit and foliage [
5]. The fungus has historically been classified as being strictly asexual; however, evidence of the sexual stage has been documented in the lab but has still not been found in the field [
6,
7]. Scab infections can result in significant yield losses to pecans if left unmanaged [
8,
9]. Although the most effective method for controlling
V. effusa is to plant resistant cultivars, selection has resulted in the adaptation of the pathogen to be pathogenic on historically resistant cultivars [
10,
11,
12,
13,
14,
15]. Thus, frequent use of fungicides is the most widely adopted management approach for controlling scab in commercial pecan orchards in the Southeast. However, fungicide control of scab is one of the largest costs for pecan growers in the region.
Demethylation-inhibiting (DMI) fungicides were first introduced for use against plant diseases in the 1970s and are now the most widely adopted and important group of fungistatic agents used worldwide both in medicine and in agriculture [
16]. Since their discovery, over 30 DMI fungicides have been synthesized for use in agriculture [
17]. They function by impeding sterol C-14 a-demethylation of 24-methylenedihydrolanosterol, which is a critical component in the formation of ergosterol in fungi. Ergosterol is vital to fungal cell membranes and regulates membrane permeability and fluidity, which is why the enzymes that synthesize ergosterol are important targets for anti-fungal agents [
18]. Since the DMIs have been so widely used and repeatedly applied to large areas of crops year after year, there have been many reports of resistance in fungi, beginning in the 1980s, only a decade after their release [
19,
20,
21]. DMI fungicides are single-site, broad-spectrum fungicides that are used for the pre- and post-infection control of various pathogens [
22]. The development of resistance to the DMI fungicides has led to the decreased sensitivity of various pathogens to a range of DMI fungicides, including the apple scab pathogen
Venturia inaequalis, a close relative to the pecan scab pathogen,
V. effusa [
23]. DMI fungicides were first labeled for use against
V. effusa in 1988 and are heavily used to combat scab each year [
24]. The DMIs labeled for use in pecan include difenoconazole, propiconazole, fenbuconazole, tetraconazole, metconazole, flutriafol, mefentrifluconazole, and tebuconazole. Isolates of
V. effusa collected in 2003 were found to be less sensitive to propiconazole when compared to the baseline isolates reported in 1997 [
25,
26]. Standish et al. [
27] reported the field resistance of
V. effusa to tebuconazole and confirmed the reduced sensitivity using an in vitro assay. In a separate study, Standish et al. [
28] demonstrated insensitivity to tebuconazole to be phenotypically stable. Luckily, there are fungicides belonging to several other fungicide classes (Fungicide Resistance Action Committee [FRAC] Codes: 3, 11 U12, P7, 30, 1, M03, and 7) that are labelled for use on pecan to help mitigate
V. effusa [
29].
To date, resistance to DMI fungicides has been reported in 37 fungal species. Cross resistance has been documented among DMI fungicides used against the same pathogen [
30,
31,
32,
33]. Cross resistance has been observed in
V. effusa among tebuconazole, fenbuconazole, and propiconazole, but has not yet been found for difenoconazole, which is widely applied to control scab [
25,
26]. Resistance to DMIs is typically caused by amino acid changes in the
CYP51 gene, overexpression of the
CYP51 gene, or by efflux pumps reducing intracellular fungicide accumulation within the pathogen [
18,
34,
35]. Tucker et al. [
36] investigated the mechanism of resistance to DMI fungicides in the pathogen
Blumeria graminis and found five separate amino acid substitutions in the
CYP51B target gene, four of which were novel, showing that multiple mutations may confer resistance to DMI fungicides.
V. inaequalis, a close relative of
V. effusa, also displays resistance to the DMI fungicides. Villani et al. [
37] investigated the mechanisms of DMI resistance in
V. inaequalis and found overexpression of the
CYP51A1 gene in resistant isolates compared to the sensitive isolates. Hayashi et al. [
38] demonstrated that ABC transporters can lead to the decreased sensitivity of
Botrytis cinerea to DMI fungicides. The results from these studies show that several potential factors may be contributing to the resistance of
V. effusa to the DMI fungicides.
The fungicide resistance studies describing resistance in
V. effusa to the DMIs to date have reported only phenotypic data, and the specific mechanism of resistance remains unknown [
26,
27,
39]. The goal of the current study was to determine the mechanism of resistance of
V. effusa to the DMI tebuconazole to provide further insights regarding DMI insensitivity prevalence in commercial pecan orchards in the southeastern U.S.
3. Discussion
Our results indicate that difenoconazole and mefentrifluconazole are both highly active on
V. effusa, while tebuconazole is not, due to the presence of resistant isolates. Confirmation of reduced sensitivity led to the collection of resistant isolates and investigation of the exact mechanism of resistance. Although some possible mechanisms of resistance in
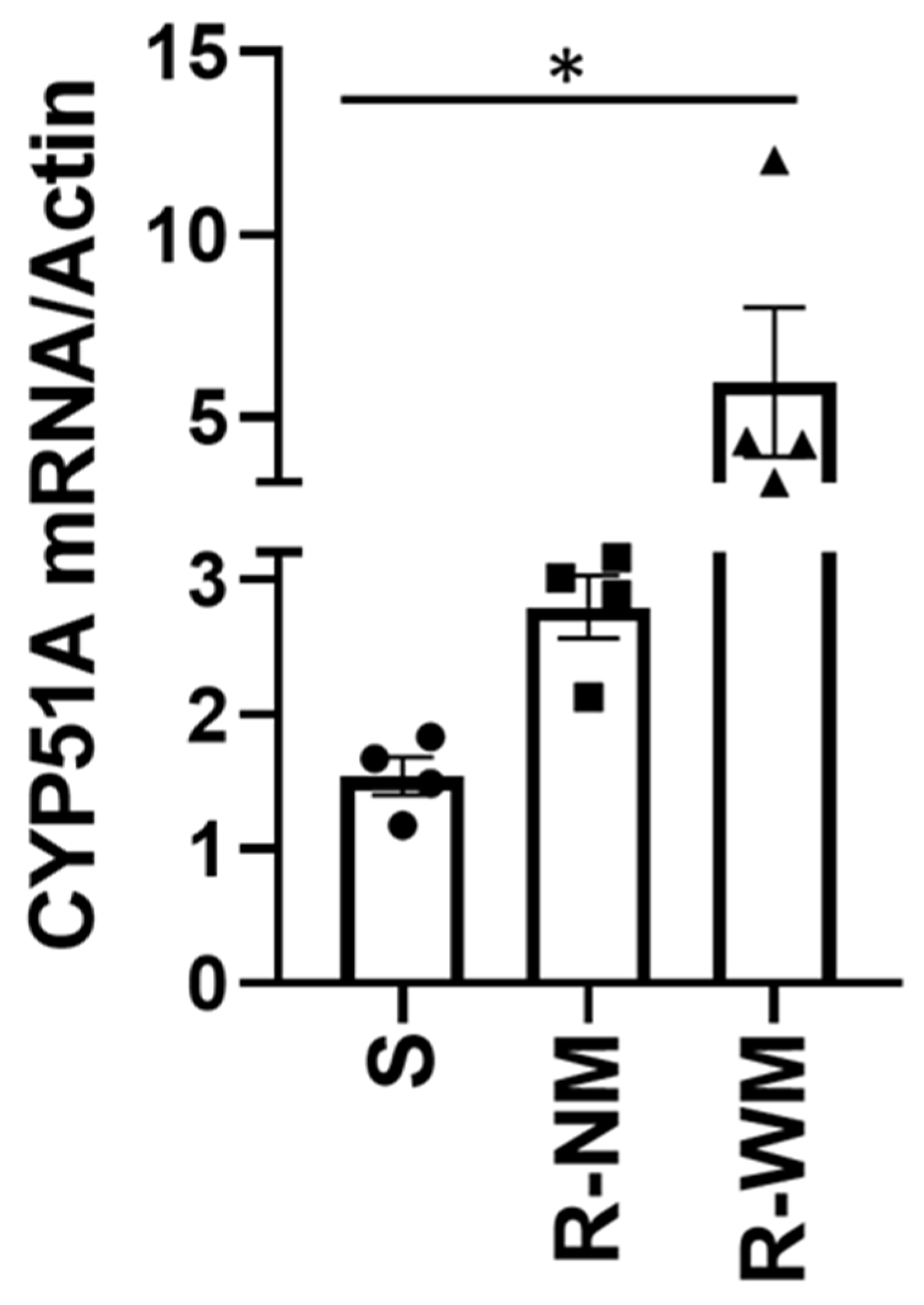
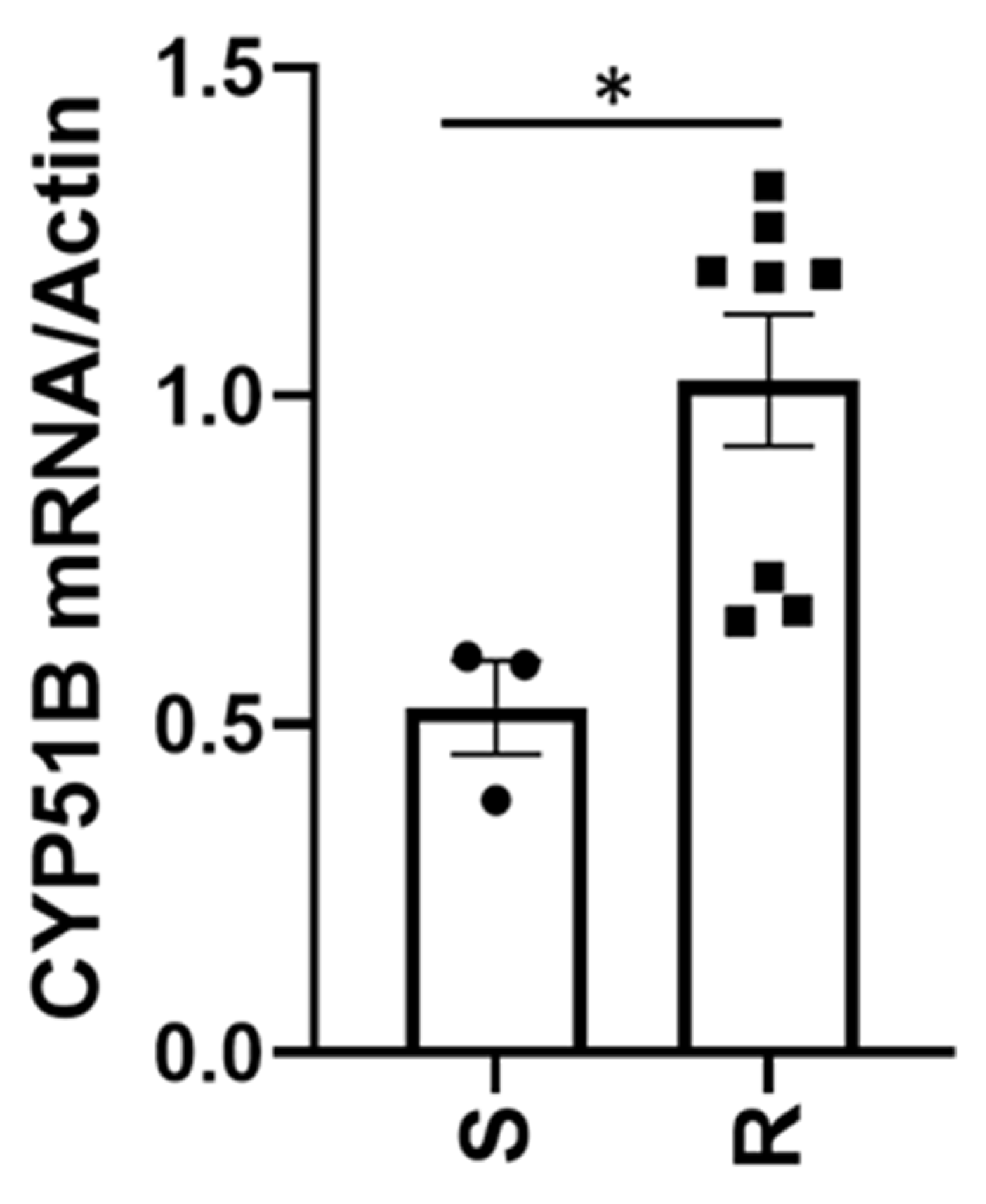
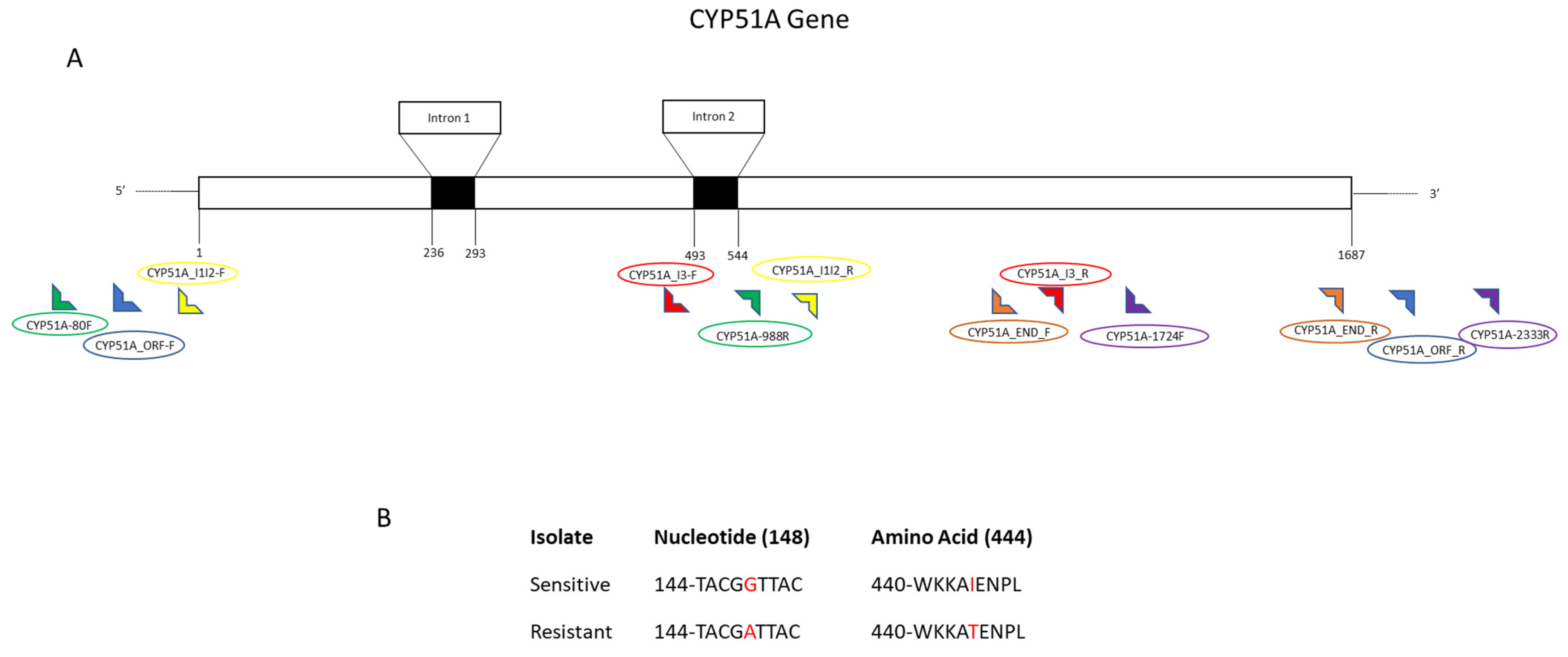
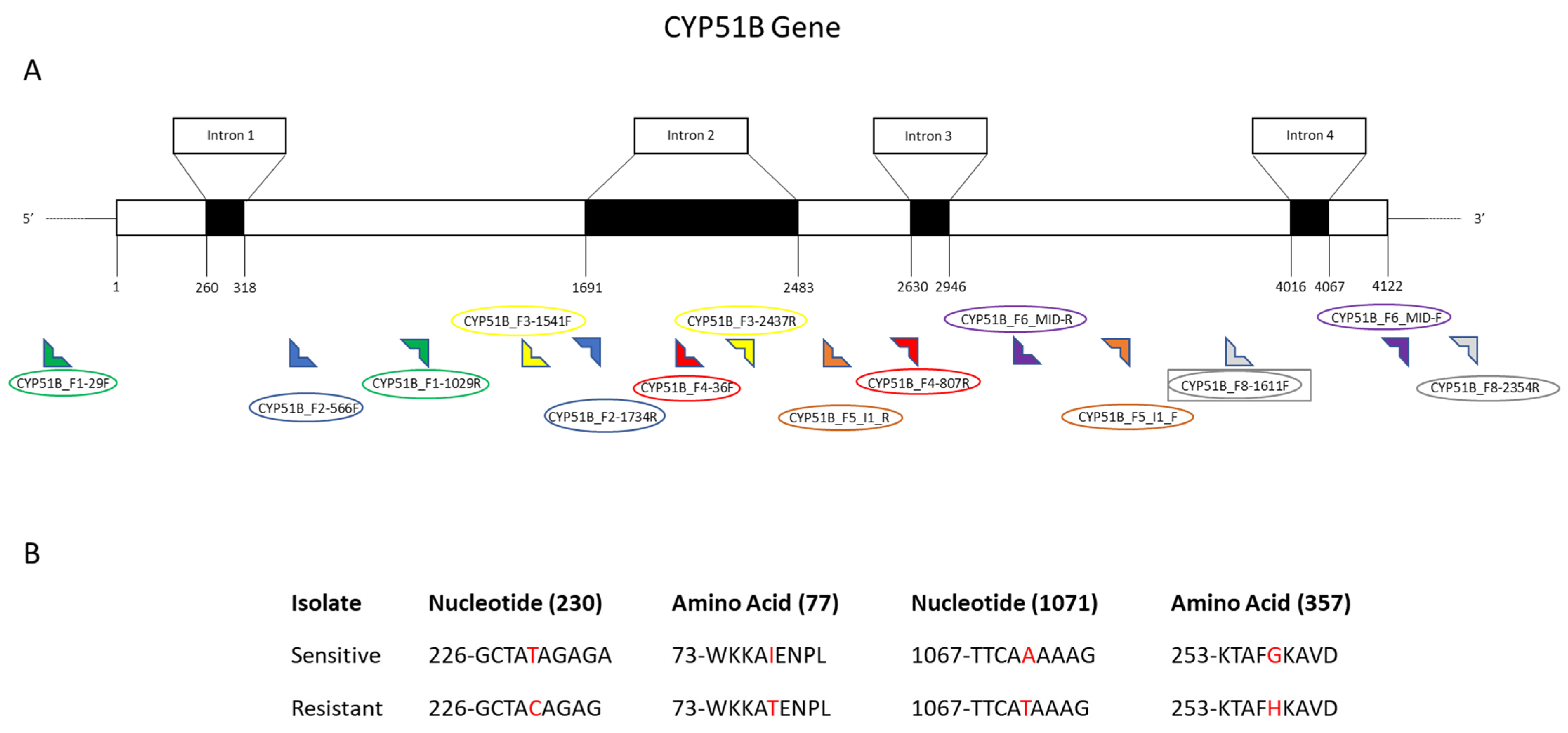
V. effusa to the DMI fungicides are presented, definitive conclusions regarding the mechanism of resistance should not be made. We suggest two mechanisms based on the results of this study, including novel mutations in both the
CYP51A and
B genes (G444D, G357H, I77T, and I77L) (
Table 4), as well as 3.0- and 1.9-fold increases in expression of the
CYP51A and
B genes, respectively (
Figure 2 and
Figure 3). These are novel mutations and have not been previously described for resistance to fungicidal compounds. These same mutations, as well as overexpression may also be responsible for the resistance of
V. effusa observed to fenbuconazole and propiconazole [
25,
26,
40]. Mutations in the
CYP51 gene are a common cause of resistance to DMIs and have been found in several other pathogens [
36,
41,
42]. Overexpression of the
CYP51 genes is also commonly found in resistant isolates of various pathogens [
43,
44,
45]. We anticipated either mutations or overexpression of the
CYP51 gene to be the cause of the resistance, and we found mutations and overexpression to be present in DMI-resistant isolates of
V. effusa. The resulting amino acid changes and overexpression in isolates of a single pathogen species exhibiting resistance to the DMI fungicides is also not uncommon [
46,
47]. While this study did not address ABC transporters and other efflux transporters, it is possible that they may also play a role in resistance to the DMI fungicides in
V. effusa, and future studies should be aimed at investigating ABC transporters and other efflux transporters as potential sources of DMI resistance.
The DMI fungicides are used heavily in commercial pecan orchards to control
V. effusa. Understanding fungicide sensitivity in
V. effusa can aide in the further development and strengthening of fungicide resistance management rotation programs. The results help outline the necessity for further fungicide research and development required to control
V. effusa efficiently and sustainably. Difenoconazole is the most widely used DMI fungicide in commercial pecan orchards and is labeled for use on pecan only as a mixture combined with other active ingredients. The popular combination products are Amistar Top (difenoconazole + azoxystrobin) and Miravis Top (difenoconazole + pydiflumetofen). Combination products with more than one active ingredient in different fungicide classes can increase disease control and contribute to delaying fungicide resistance in certain pathogens. However, if one of the active ingredients in the premixture begins to lose its efficacy due to resistance development in the pathogen, the other active ingredient in the combination is more at risk of resistance development. So far, no resistance of
V. effusa to difenoconazole has been reported, and combination products containing difenoconazole remain popular among commercial pecan growers. However, there is currently no baseline sensitivity data pertaining to
V. effusa sensitivity to difenoconazole; therefore, slight shifts in sensitivity may go unnoticed and cannot be confirmed in the lab. Although resistance to difenoconazole has not been reported for
V. effusa, it has been reported for several other pathogens, including
V. inaequalis,
Lasiodiplodia theobromae,
Penicillium spp.,
Botrytis cinereal,
Alternaria spp.,
Aspergillus fumigatus, etc. [
36,
45,
48,
49,
50,
51,
52].
Mefentrifluconazole was recently labeled for use on pecan (Cevya), and although resistance has not yet been reported in
V. effusa, resistance has already been detected in other pathosystems [
33]. Not only has resistance to mefentrifluconazole been detected in other pathosystems, but cross-resistance between mefentrifluconazole and tebuconazole, as well as mefentrifluconazole and difenoconazole has been shown in other pathosystems [
33,
53,
54]. Mefentrifluconazole is not yet widely used in commercial pecan orchards but may play a role in future fungicide rotation programs. However, since Cevya is a stand-alone DMI fungicide that runs a high risk of resistance development, its use should be limited, and it should only be applied in strict rotation and never applied alone in consecutive applications. Tebuconazole products are no longer widely used among commercial pecan growers due to very low activity as a result of fungicide resistance developing in
V. effusa, as confirmed by our field and in vitro studies. Our results indicate that resistance of
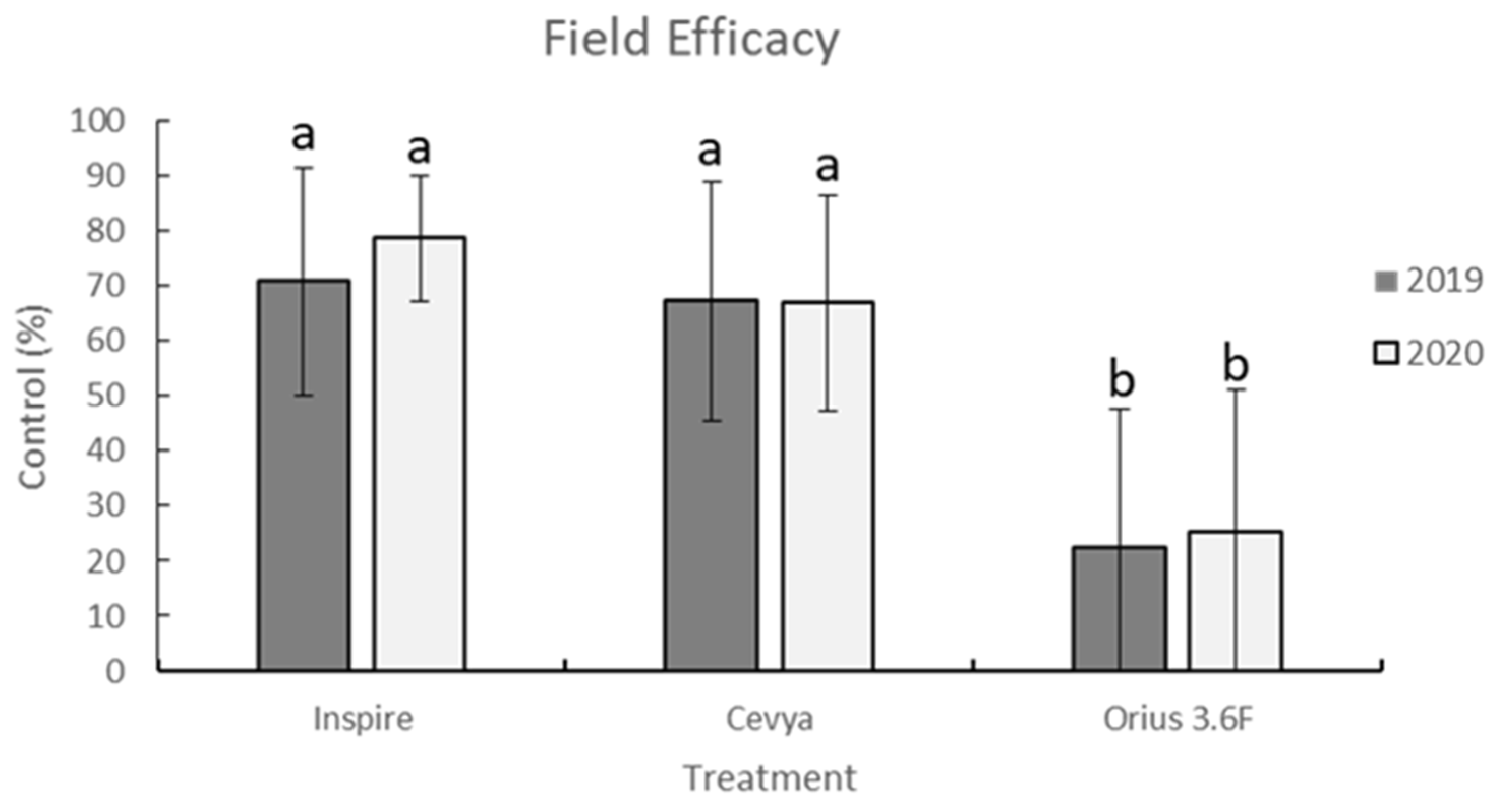
V. effusa to tebuconazole is widespread, and that the efficacy of tebuconazole across southern Georgia is low (
Figure 1;
Table 1 and
Table 2). Mefentrifluconazole and difenoconazole are both newer DMI fungicides and are active on scab even when other DMI fungicides are not. Newer DMI fungicides maintaining efficacy while older DMI fungicides fall victim to resistance development is not a novel observation [
33,
55]. It has been proposed that the high structural flexibility of the mefentrifluconazole molecule is to blame for the limited cross resistance being observed [
56]. Because of flexible isopropanol linkers, mefentrifluconazole molecules are able to settle into the binding pocket of the
CYP51 enzyme, resulting in strong inhibition of enzymatic activity, even when target site alterations due to amino acid substitutions may be present [
33,
56].
The goal of determining the mechanism of resistance is to both add to our knowledge regarding resistance development in a pathogen, as well as providing a basis for developing detection methods that can be used rapidly to identify and track the specific resistance trait in orchard populations of
V. effusa in the future. These rapid detection methods are not uncommon and have been proven effective in various other pathogens in regard to resistance to the DMIs, as well as other fungicide groups [
57,
58]. Since
V. effusa is a very slow-growing pathogen, taking approximately 30 days for a colony to grow to a diameter of 25 mm, a rapid method for detecting resistance to DMI (and other) fungicides would be a valuable tool to provide to growers and other stakeholders to better characterize the pathogen population present in their orchard, which ultimately will help optimize management of the populations of
V. effusa to minimize the risk of further development and spread of fungicide resistance, and improve the efficacy and sustainability of scab control.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}