
Effects of Serine/Arginine Enriched Protein BmUP on the Development of Male Silkworm Reproductive Organs


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Cells
2.2. SgRNA Design and Synthesis
2.3. Vector Construction
2.4. Construction of BmUP Overexpressing Transgenic Line
2.5. Quantitative Real-Time PCR
2.6. Western Blot
2.7. Subcellular Localization
2.8. Dual Luciferase Reporter Experiment
2.9. Electrophoretic Mobility Shift Assays
2.10. Data Analysis
3. Results
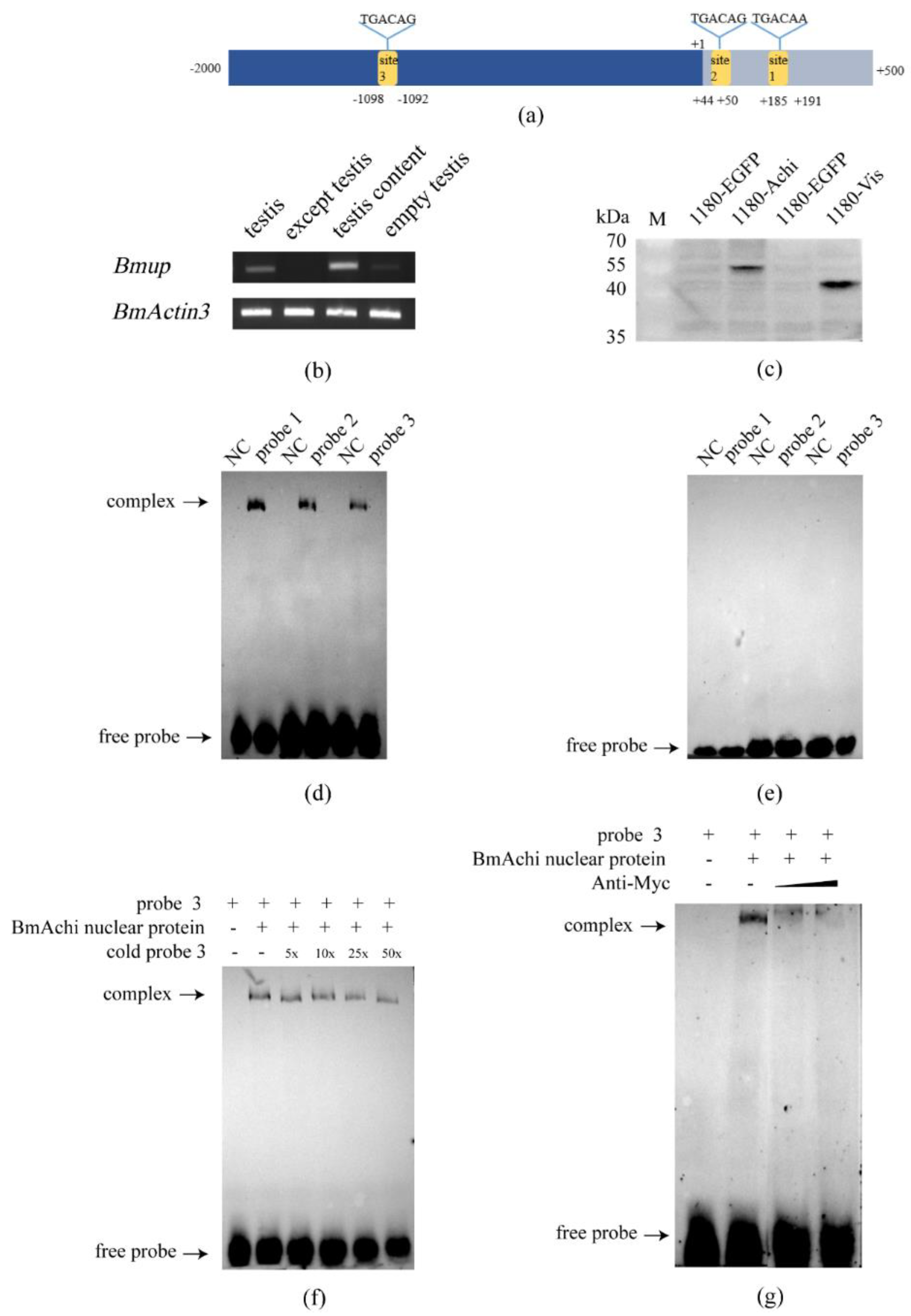
3.1. BmUP Is Highly Expressed in the Testis
3.2. CRISPR/Cas9-Mediated Mutagenesis of the BmUP Gene
3.3. Construction of a BmUP Overexpressing Transgenic Line
3.4. Cellular Location of the BmUP Gene and Observation of the Transgenic Phenotype
3.5. The Silkworm Meiosis Blocker Gene Achi/Vis Regulates the Expression of BmUP
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| achi | Achiya |
| BmE | Embryonic cells of silkworm |
| BmN | Ovary cells of silkworm |
| EMSA EGFP | Electrophoretic mobility shift assay Enhanced green fluorescent protein |
| QPCR | Quantitative Real-Time PCR |
| SR | Serine/Arginine-riched protein |
| vis | Vismay |
| BCA | Bicinchoninic acid |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| PVDF | Polyvinylidene fluoride |
| TBST | TBS with Tween-20 |
References
- Black, D.L. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [Green Version]
- Black, D.L. Protein diversity from alternative splicing: A challenge for bioinformatics and post-genome biology. Cell 2000, 103, 367–370. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Will, C.L.; Luhrmann, R. Spliceosomal UsnRNP biogenesis, structure and function. Curr. Opin. Cell Biol. 2001, 13, 290–301. [Google Scholar] [CrossRef]
- Lin, S.R.; Fu, X.D. SR proteins and related factors in alternative splicing. Altern. Splicing Postgenomic Eraa 2007, 623, 107–122. [Google Scholar]
- Beier, D.H.; Carrocci, T.J.; van der Feltz, C.; Tretbar, U.S.; Paulson, J.C.; Grabowski, N.; Hoskins, A.A. Dynamics of the DEAD-box ATPase Prp5 RecA-like domains provide a conformational switch during spliceosome assembly. Nucleic Acids Res. 2019, 47, 10842–10851. [Google Scholar] [CrossRef]
- Chou, T.B.; Zachar, Z.; Bingham, P.M. Developmental expression of a regulatory gene is programmed at the level of splicing. Embo J. 1987, 6, 4095–4101. [Google Scholar] [CrossRef]
- Boggs, R.T.; Gragor, P.; Idrlss, S.; Belote, J.M.; Mckeown, M. Regulation of Sexual-Differentiation in D. melanogaster via Alternative Splicing Of RNA from the transformer Gene. Cell 1987, 50, 739–747. [Google Scholar] [CrossRef]
- Amrein, H.; Gorman, M.; Nothiger, R. The Sex-Determining Gene tra-2 of Drosophila Encodes a Putative RNA-Binging Protein. Cell 1988, 55, 1025–1035. [Google Scholar] [CrossRef]
- Fu, X.D.; Maniatis, T. Isolation of a Complementary-DNA That Encodes the Mammalian Splicing Factor SC35. Science 1992, 256, 535–538. [Google Scholar] [CrossRef]
- Ge, H.; Zuo, P.; Manley, J.L. Primary Structure of the Human Splicing Factor ASF Reveals Similarities with Drosophila Regulators. Cell 1991, 66, 373–382. [Google Scholar] [CrossRef]
- Manley, J.L.; Krainer, A.R. A rational nomenclature for serine/arginine-rich protein splicing factors (SR proteins). Genes Dev. 2010, 24, 1073–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, N.; Bachorik, J.L.; Dreyfuss, G. Transportin-SR, a nuclear import receptor for SR proteins. J. Cell Biol. 1999, 145, 1145–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcarcel, J.; Green, M.R. The SR protein family: Pleiotropic functions in pre-mRNA splicing. Trends Biochem. Sci. 1996, 21, 296–301. [Google Scholar] [CrossRef]
- Long, J.C.; Caceres, J.F. The SR protein family of splicing factors: Master regulators of gene expression. Biochem. J. 2009, 417, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S. SR Proteins: Binders, Regulators, and Connectors of RNA. Mol. Cells 2017, 40, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Xiao, R.; Sun, Y.; Ding, J.H.; Lin, S.R.; Rose, D.W.; Rosenfeld, M.G.; Fu, X.D.; Li, X. Splicing regulator SC35 is essential for genomic stability and cell proliferation during mammalian organogenesis. Mol. Cell. Biol. 2007, 27, 5393–5402. [Google Scholar] [CrossRef] [Green Version]
- Li, X.L.; Wang, J.; Manley, J.L. Loss of splicing factor ASF/SF2 induces G2 cell cycle arrest and apoptosis, but inhibits internucleosomal DNA fragmentation. Genes Dev. 2005, 19, 2705–2714. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.Y.; Wang, P.; Han, J.; Rosenfeld, M.G.; Fu, X.D. SR proteins in vertical integration of gene expression from transcription to RNA processing to translation. Mol. Cell 2009, 35, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wee, C.D.; Havens, M.A.; Jodelka, F.M.; Hastings, M.L. Targeting SR proteins improves SMN expression in spinal muscular atrophy cells. PLoS ONE 2014, 9, e115205. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, Y. SRSF7 knockdown promotes apoptosis of colon and lung cancer cells. Oncol. Lett. 2018, 15, 5545–5552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, L.; Yu, W.; Shen, E.; Sun, W.; Liu, Y.; Kong, J.; Wu, Y.; Han, F.; Zhang, L.; Yu, T.; et al. SRSF6-regulated alternative splicing that promotes tumour progression offers a therapy target for colorectal cancer. Gut 2019, 68, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Sciabica, K.S.; Hertel, K.J. The splicing regulators Tra and Tra2 are unusually potent activators of pre-mRNA splicing. Nucleic Acids Res. 2006, 34, 6612–6620. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.G. Sex determination: Insights from the silkworm. J. Genet. 2010, 89, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Mann, R.S. Requirement for two nearly identical TGIF-related homeobox genes in Drosophila spermatogenesis. Development 2003, 130, 2853–2865. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Yang, H.; Sturgill, D.; Oliver, B.; Rabinow, L.; Samson, M.L. Sxl-Dependent, tra/tra2-Independent Alternative Splicing of the Drosophila melanogaster X-Linked Gene found in neurons. G3 2015, 5, 2865–2874. [Google Scholar] [CrossRef] [Green Version]
- Shepard, P.J.; Hertel, K. The SR protein family. Genome Biol. 2009, 10, 242. [Google Scholar] [CrossRef] [Green Version]
- Ayyar, S.; Jiang, J.; Collu, A.; White-Cooper, H.; White, R.A. Drosophila TGIF is essential for developmentally regulated transcription in spermatogenesis. Development 2003, 130, 2841–2852. [Google Scholar] [CrossRef] [Green Version]
- Hyman, C.A.; Bartholin, L.; Newfeld, S.J.; Wotton, D. Drosophila TGIF proteins are transcriptional activators. Mol. Cell. Biol. 2003, 23, 9262–9274. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Cao, G.; Sheng, J.; Xue, R.; Gong, C. BmTGIF, a Bombyx mori homolog of Drosophila DmTGIF, regulates progression of spermatogenesis. PLoS ONE 2012, 7, e47861. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences | Purpose |
|---|---|---|
| sgRNA1-F | 5′GGGCAACTATCCCGACCATG3′ | gene editing |
| sgRNA1-R | 5′CATGGTCGGGATAGTTGCCC3′ | gene editing |
| sgRNA2-F | 5′AACGGTCAGCAGGAGTGCAT3′ | gene editing |
| sgRNA2-R | 5′ATGCACTCCTGCTGACCGTTC3′ | gene editing |
| BmUP-F | 5′ATGAAGTGTCATTTTGACGTTATTT3′ | RT-PCR |
| BmUP-R | 5′TTAAACTGCTTGAATGAATCCC3′ | RT-PCR |
| JC-BmUP-F | 5′TCAAATTAATGGATGGATATGCGTA3′ | PCR |
| JC-BmUP-R | 5′AAGGGGGTCAAAATCATGTTCAAAG3′ | PCR |
| Qpcr-BmUP-F | 5′GAGAACGGTCAGCAGGAGTG3′ | qPCR |
| Qpcr-BmUP-R | 5′ACCATCGCTTTGAGCCCAGT3′ | qPCR |
| Qpcr-BmAchi-F | 5′CTCTGGAGTCGTTGTCGCTT3′ | qPCR |
| Qpcr-BmAchi-R | 5′GTATCTGATGACCGACTGCCA3′ | qPCR |
| Qpcr-BmVis-F | 5′GGGCGAACTGGAAAGAG3′ | qPCR |
| Qpcr-BmVis-R | 5′TCTACTGAATATATCTGACATCCG3′ | qPCR |
| Transgenic Line | Sequence of Insertion Sites | Chromosome |
|---|---|---|
| Over-BmUP | AAAGTCGGTCAACCTTAA-piggybac-TTAAAAAATATTGATTCC | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-B.; Li, J.; Deng, X.; Liu, L.-L.; Deng, J.; Zha, X.-F. Effects of Serine/Arginine Enriched Protein BmUP on the Development of Male Silkworm Reproductive Organs. Curr. Issues Mol. Biol. 2022, 44, 928-941. https://doi.org/10.3390/cimb44020061
Chen C-B, Li J, Deng X, Liu L-L, Deng J, Zha X-F. Effects of Serine/Arginine Enriched Protein BmUP on the Development of Male Silkworm Reproductive Organs. Current Issues in Molecular Biology. 2022; 44(2):928-941. https://doi.org/10.3390/cimb44020061
Chicago/Turabian StyleChen, Chun-Bing, Juan Li, Xuan Deng, Lian-Lian Liu, Jing Deng, and Xing-Fu Zha. 2022. "Effects of Serine/Arginine Enriched Protein BmUP on the Development of Male Silkworm Reproductive Organs" Current Issues in Molecular Biology 44, no. 2: 928-941. https://doi.org/10.3390/cimb44020061
APA StyleChen, C. -B., Li, J., Deng, X., Liu, L. -L., Deng, J., & Zha, X. -F. (2022). Effects of Serine/Arginine Enriched Protein BmUP on the Development of Male Silkworm Reproductive Organs. Current Issues in Molecular Biology, 44(2), 928-941. https://doi.org/10.3390/cimb44020061