
Therapeutic Anti-Depressant Potential of Microbial GABA Produced by Lactobacillus rhamnosus Strains for GABAergic Signaling Restoration and Inhibition of Addiction-Induced HPA Axis Hyperactivity


Abstract
:1. Introduction
2. GABA and GABA Receptors
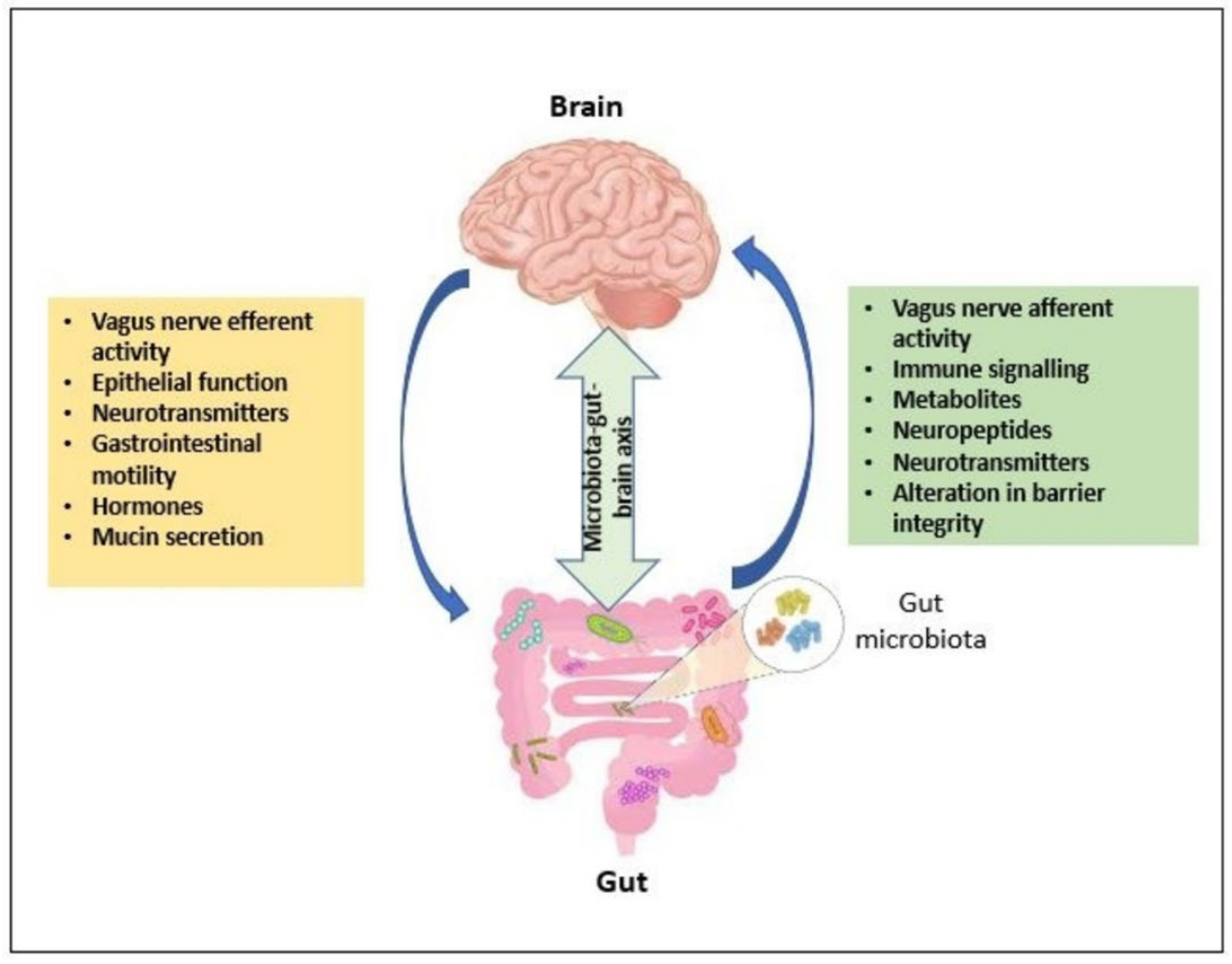
GABA-Ergic Signaling across the Microbiota–Gut–Brain (MGB) Axis
3. Probiotics
4. Lactobacillus rhamnosus
Therapeutic Advantages of L. rhamnosus Strains
5. Lactobacillus rhamnosus Strains and GABAergic Signaling
5.1. L. rhamnosus (JB-1)
5.2. L. rhamnosus GG
5.3. L. rhamnosus LR-2
5.4. L. rhamnosus YS9
5.5. L. rhamnosus SP1
5.6. L. rhamnosus HN001
6. Therapeutic Potential of Lactobacillus rhamnosus
7. GABAergic Signaling in Other Probiotic Bacterial Species
8. Challenges and Future Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- United Nations Office on Drugs and Crime. World Drug Report 2021. Available online: //www.unodc.org/unodc/en/data-and-analysis/wdr2021.html (accessed on 6 January 2022).
- Koob, G.F.; Volkow, N.D. Neurobiology of Addiction: A neurocircuitry analysis. Lancet Psychiatry 2016, 3, 760–773. [Google Scholar] [CrossRef]
- Koob, G.F. Neurobiology of opioid addiction: Opponent process, hyperkatifeia, and negative reinforcement. Biol. Psychiatry 2020, 87, 44–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochalska, K.; Oakden, W.; Słowik, T.; Chudzik, A.; Pankowska, A.; Łazorczyk, A.; Kozioł, P.; Andres-Mach, M.; Pietura, R.; Rola, R.; et al. Dietary Supplementation with Lactobacillus Rhamnosus JB-1 restores brain neurochemical balance and mitigates the progression of mood disorder in a rat model of chronic unpredictable mild stress. Nutr. Res. 2020, 82, 44–57. [Google Scholar] [CrossRef]
- Vasconcelos, M.; Stein, D.J.; Gallas-Lopes, M.; Landau, L.; de Almeida, R.M.M. Corticotropin-releasing factor receptor signaling and modulation: Implications for stress response and resilience. Trends Psychiatry Psychother. 2020, 42, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Slater, P.G.; Yarur, H.E.; Gysling, K. Corticotropin-releasing factor receptors and their interacting proteins: Functional consequences. Mol. Pharmacol. 2016, 90, 627–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mody, I.; Maguire, J. The reciprocal regulation of stress hormones and GABAA receptors. Front. Cell. Neurosci. 2012, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Peng, T.; Gaur, U.; Silva, M.; Little, P.; Chen, Z.; Qiu, W.; Zhang, Y.; Zheng, W. Role of corticotropin releasing factor in the neuroimmune mechanisms of depression: Examination of current pharmaceutical and herbal therapies. Front. Cell. Neurosci. 2019, 13, 290. [Google Scholar] [CrossRef]
- Yong, S.J.; Tong, T.; Chew, J.; Lim, W.L. Antidepressive mechanisms of probiotics and their therapeutic potential. Front. Neurosci. 2020, 13, 1361. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the manipulation of bacteria-gut-brain signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Bonaz, B.; Bazin, T.; Pellissier, S. The vagus nerve at the interface of the microbiota-gut-brain axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- Rashmi, D.; Zanan, R.; John, S.; Khandagale, K.; Nadaf, A. Chapter 13-γ-Aminobutyric Acid (GABA): Biosynthesis, Role, Commercial Production, and Applications. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 57, pp. 413–452. [Google Scholar]
- Jewett, B.E.; Sharma, S. Physiology, GABA. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Kondziella, D. The Top 5 Neurotransmitters from a clinical neurologist’s perspective. Neurochem. Res. 2017, 42, 1767–1771. [Google Scholar] [CrossRef]
- Pedrón, V.T.; Varani, A.P.; Bettler, B.; Balerio, G.N. GABAB receptors modulate morphine antinociception: Pharmacological and genetic approaches. Pharmacol. Biochem. Behav. 2019, 180, 11–21. [Google Scholar] [CrossRef]
- Strużyńska, L.; Sulkowski, G. Relationships between glutamine, glutamate, and GABA in nerve endings under Pb-toxicity conditions. J. Inorg. Biochem. 2004, 98, 951–958. [Google Scholar] [CrossRef]
- Govindpani, K.; Calvo-Flores Guzmán, B.; Vinnakota, C.; Waldvogel, H.J.; Faull, R.L.; Kwakowsky, A. Towards a better understanding of GABAergic remodeling in Alzheimer’s disease. Int. J. Mol. Sci. 2017, 18, 1813. [Google Scholar] [CrossRef] [Green Version]
- Sherwin, E.; Sandhu, K.V.; Dinan, T.G.; Cryan, J.F. May the force be with you: The light and dark sides of the microbiota-gut-brain axis in neuropsychiatry. CNS Drugs 2016, 30, 1019–1041. [Google Scholar] [CrossRef] [Green Version]
- Janik, R.; Thomason, L.A.M.; Stanisz, A.M.; Forsythe, P.; Bienenstock, J.; Stanisz, G.J. Magnetic resonance spectroscopy reveals oral Lactobacillus Promotion of Increases in Brain GABA, N-acetyl aspartate and glutamate. NeuroImage 2016, 125, 988–995. [Google Scholar] [CrossRef]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [Green Version]
- Patterson, E.; Ryan, P.M.; Wiley, N.; Carafa, I.; Sherwin, E.; Moloney, G.; Franciosi, E.; Mandal, R.; Wishart, D.S.; Tuohy, K.; et al. Gamma-aminobutyric acid-producing lactobacilli positively affect metabolism and depressive-like behaviour in a mouse model of metabolic syndrome. Sci. Rep. 2019, 9, 16323. [Google Scholar] [CrossRef]
- Pirbaglou, M.; Katz, J.; de Souza, R.J.; Stearns, J.C.; Motamed, M.; Ritvo, P. Probiotic supplementation can positively affect anxiety and depressive symptoms: A systematic review of randomized controlled trials. Nutr. Res. N. Y. N 2016, 36, 889–898. [Google Scholar] [CrossRef]
- Wallace, C.J.K.; Milev, R. The Effects of probiotics on depressive symptoms in humans: A systematic review. Ann. Gen. Psychiatry 2017, 16, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browning, K.N.; Verheijden, S.; Boeckxstaens, G.E. The vagus nerve in appetite regulation, mood, and intestinal inflammation. Gastroenterology 2017, 152, 730–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaz, B.; Sinniger, V.; Pellissier, S. The vagus nerve in the neuro-immune axis: Implications in the pathology of the gastrointestinal tract. Front. Immunol. 2017, 8, 1452. [Google Scholar] [CrossRef] [PubMed]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The gut microbiota mediates the anti-seizure effects of the ketogenic diet. Cell 2018, 173, 1728–1741.e13. [Google Scholar] [CrossRef] [Green Version]
- Klarer, M.; Arnold, M.; Gunther, L.; Winter, C.; Langhans, W.; Meyer, U. Gut Vagal afferents differentially modulate innate anxiety and learned fear. J. Neurosci. 2014, 34, 7067–7076. [Google Scholar] [CrossRef] [Green Version]
- Dan, Z.; Mao, X.; Liu, Q.; Guo, M.; Zhuang, Y.; Liu, Z.; Chen, K.; Chen, J.; Xu, R.; Tang, J.; et al. Altered gut microbial profile is associated with abnormal metabolism activity of autism spectrum disorder. Gut Microbes 2020, 11, 1246–1267. [Google Scholar] [CrossRef]
- El-Ansary, A.; Bacha, A.B.; Bjørklund, G.; Al-Orf, N.; Bhat, R.S.; Moubayed, N.; Abed, K. Probiotic treatment reduces the autistic-like excitation/inhibition imbalance in juvenile hamsters induced by orally administered propionic acid and clindamycin. Metab. Brain Dis. 2018, 33, 1155–1164. [Google Scholar] [CrossRef]
- Yunes, R.A.; Poluektova, E.U.; Dyachkova, M.S.; Klimina, K.M.; Kovtun, A.S.; Averina, O.V.; Orlova, V.S.; Danilenko, V.N. GABA Production and Structure of GadB/GadC Genes in Lactobacillus and Bifidobacterium Strains from Human Microbiota. Anaerobe 2016, 42, 197–204. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Nielsen, C.U.; Carstensen, M.; Brodin, B. Carrier-mediated γ-aminobutyric acid transport across the basolateral membrane of human intestinal Caco-2 Cell Monolayers. Eur. J. Pharm. Biopharm. 2012, 81, 458–462. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of action of probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [Green Version]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Immune-mediated mechanisms of action of probiotics and synbiotics in treating pediatric intestinal diseases. Nutrients 2018, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Geva-Zatorsky, N.; Sefik, E.; Kua, L.; Pasman, L.; Tan, T.G.; Ortiz-Lopez, A.; Yanortsang, T.B.; Yang, L.; Jupp, R.; Mathis, D.; et al. Mining the human gut microbiota for immunomodulatory organisms. Cell 2017, 168, 928–943.e11. [Google Scholar] [CrossRef] [Green Version]
- Bambury, A.; Sandhu, K.; Cryan, J.F.; Dinan, T.G. Finding the needle in the haystack: Systematic identification of psychobiotics: Finding the needle in the haystack. Br. J. Pharmacol. 2018, 175, 4430–4438. [Google Scholar] [CrossRef] [Green Version]
- Berings, M.; Karaaslan, C.; Altunbulakli, C.; Gevaert, P.; Akdis, M.; Bachert, C.; Akdis, C.A. Advances and highlights in allergen immunotherapy: On the way to sustained clinical and immunologic tolerance. J. Allergy Clin. Immunol. 2017, 140, 1250–1267. [Google Scholar] [CrossRef] [Green Version]
- Pogačar, M.Š.; Mičetić-Turk, D.; Fijan, S. Chapter 31-Probiotics: Current regulatory aspects of probiotics for use in different disease conditions. In Probiotics in the Prevention and Management of Human Diseases; Dwivedi, M.K., Amaresan, N., Sankaranarayanan, A., Kemp, E.H., Eds.; Academic Press: Boston, MA, USA, 2022; pp. 465–499. ISBN 978-0-12-823733-5. [Google Scholar]
- Katkowska, M.; Garbacz, K.; Kusiak, A. Probiotics: Should all patients take them? Microorganisms 2021, 9, 2620. [Google Scholar] [CrossRef]
- Chiang, M.-C.; Chen, C.-L.; Feng, Y.; Chen, C.-C.; Lien, R.; Chiu, C.-H. Lactobacillus Rhamnosus Sepsis associated with probiotic therapy in an extremely preterm infant: Pathogenesis and a review for clinicians. J. Microbiol. Immunol. Infect. Wei Mian Yu Gan Ran Za Zhi 2021, 54, 575–580. [Google Scholar] [CrossRef]
- Naqvi, S.S.B.; Nagendra, V.; Hofmeyr, A. Probiotic Related Lactobacillus Rhamnosus Endocarditis in a patient with liver cirrhosis. IDCases 2018, 13, e00439. [Google Scholar] [CrossRef]
- Appel-da-Silva, M.C.; Narvaez, G.A.; Perez, L.R.R.; Drehmer, L.; Lewgoy, J. Saccharomyces Cerevisiae Var. Boulardii Fungemia following probiotic treatment. Med. Mycol. Case Rep. 2017, 18, 15–17. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Girones, R.; Koutsoumanis, K.; Herman, L.; Lindqvist, R.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 5: Suitability of taxonomic units notified to EFSA until September 2016. EFSA J. Eur. Food Saf. Auth. 2017, 15, e04663. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Cheng, M.; Yang, X.; Lu, Y.; Yin, H.; Zeng, Y.; Wang, R.; Jiang, Y.; Yang, W.; Wang, J.; et al. Probiotic Lactobacillus Rhamnosus GG promotes mouse gut microbiota diversity and T cell differentiation. Front. Microbiol. 2020, 11, 3216. [Google Scholar] [CrossRef] [PubMed]
- Ansari, J.M.; Colasacco, C.; Emmanouil, E.; Kohlhepp, S.; Harriott, O. Strain-level diversity of commercial probiotic isolates of Bacillus, Lactobacillus, and Saccharomyces Species Illustrated by molecular identification and phenotypic profiling. PLoS ONE 2019, 14, e0213841. [Google Scholar] [CrossRef] [PubMed]
- Calasso, M.; Gobbetti, M. Lactic acid bacteria|Lactobacillus spp.: Other species. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 125–131. ISBN 978-0-12-374407-4. [Google Scholar]
- Wang, T.; Yan, H.; Lu, Y.; Li, X.; Wang, X.; Shan, Y.; Yi, Y.; Liu, B.; Zhou, Y.; Lü, X. Anti-obesity effect of Lactobacillus rhamnosus LS-8 and Lactobacillus Crustorum MN047 on high-fat and high-fructose diet mice base on inflammatory response alleviation and gut microbiota regulation. Eur. J. Nutr. 2020, 59, 2709–2728. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and their role as members of the human gut microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitellio, P.; Celano, G.; Bonfrate, L.; Gobbetti, M.; Portincasa, P.; De Angelis, M. Effects of bifidobacterium longum and Lactobacillus Rhamnosus on gut microbiota in patients with lactose intolerance and persisting functional gastrointestinal symptoms: A randomised, double-blind, cross-over study. Nutrients 2019, 11, 886. [Google Scholar] [CrossRef] [Green Version]
- Bozzi Cionci, N.; Baffoni, L.; Gaggìa, F.; Di Gioia, D. Therapeutic microbiology: The role of bifidobacterium breve as food supplement for the prevention/treatment of paediatric diseases. Nutrients 2018, 10, 1723. [Google Scholar] [CrossRef] [Green Version]
- Burton, J.P.; Chanyi, R.M.; Schultz, M. Chapter 19-common organisms and probiotics: Streptococcus Thermophilus (Streptococcus Salivarius Subsp. Thermophilus). In The Microbiota in Gastrointestinal Pathophysiology; Floch, M.H., Ringel, Y., Walker, W.A., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 165–169. ISBN 978-0-12-804024-9. [Google Scholar]
- Vitetta, L.; Llewellyn, H.; Oldfield, D. Gut dysbiosis and the intestinal microbiome: Streptococcus thermophilus a key probiotic for reducing uremia. Microorganisms 2019, 7, 228. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Yu, Z.; Liu, W.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Probiotic characteristics of Bacillus coagulans and associated implications for human health and diseases. J. Funct. Foods 2020, 64, 103643. [Google Scholar] [CrossRef]
- Gaziano, R.; Sabbatini, S.; Roselletti, E.; Perito, S.; Monari, C. Saccharomyces cerevisiae-based probiotics as novel antimicrobial agents to prevent and treat vaginal infections. Front. Microbiol. 2020, 11, 954–966. [Google Scholar] [CrossRef] [Green Version]
- Zommiti, M.; Cambronel, M.; Maillot, O.; Barreau, M.; Sebei, K.; Feuilloley, M.; Ferchichi, M.; Connil, N. Evaluation of probiotic properties and safety of enterococcus faecium isolated from artisanal tunisian meat “Dried Ossban”. Front. Microbiol. 2018, 9, 1685. [Google Scholar] [CrossRef] [Green Version]
- Wan, M.L.-Y.; Chen, Z.; Shah, N.P.; El-Nezami, H. Effects of Lactobacillus rhamnosus GG and Escherichia coli nissle 1917 cell-free supernatants on modulation of mucin and cytokine secretion on human intestinal epithelial HT29-MTX Cells. J. Food Sci. 2018, 83, 1999–2007. [Google Scholar] [CrossRef]
- Larsen, K.; Hardison, R.L.; Justice, S.S.; Schatz, F.; Arlier, S.; Semerci, N.; Guzeloglu-Kayisli, O.; Lockwood, C.J.; Kayisli, U.A. The Escherichia coli Nissle 1917-Based Probiotic (EcN) Promotes Anti-Inflammation by Inhibiting Decidual Cell Expressed Pro-Inflammatory Cytokines; Sage Publications Inc.: Thousand Oaks, CA, USA, 2017; Volume 24, p. 111A. [Google Scholar]
- Martín, R.; Chamignon, C.; Mhedbi-Hajri, N.; Chain, F.; Derrien, M.; Escribano-Vázquez, U.; Garault, P.; Cotillard, A.; Pham, H.P.; Chervaux, C.; et al. The potential probiotic Lactobacillus Rhamnosus CNCM I-3690 strain protects the intestinal barrier by stimulating both mucus production and cytoprotective response. Sci. Rep. 2019, 9, 5398. [Google Scholar] [CrossRef] [Green Version]
- Gorbach, S.; Doron, S.; Magro, F. Chapter 7-Lactobacillus rhamnosus GG. In The Microbiota in Gastrointestinal Pathophysiology; Floch, M.H., Ringel, Y., Allan Walker, W., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 79–88. ISBN 978-0-12-804024-9. [Google Scholar]
- Didari, T.; Solki, S.; Mozaffari, S.; Nikfar, S.; Abdollahi, M. A systematic review of the safety of probiotics. Expert Opin. Drug Saf. 2014, 13, 227–239. [Google Scholar] [CrossRef]
- Hoffmann, A.; Kleniewska, P.; Pawliczak, R. Antioxidative activity of probiotics. Arch. Med. Sci. AMS 2019, 17, 792–804. [Google Scholar] [CrossRef]
- Song, H.Y.; Yu, R.C. Optimization of culture conditions for gamma-aminobutyric acid production in fermented adzuki bean milk. J. Food Drug Anal. 2018, 26, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Jin, G.; Pang, X.; Mo, Q.; Bao, J.; Liu, T.; Wu, J.; Xie, R.; Liu, X.; Liu, J.; et al. Lactobacillus Rhamnosus GG Colonization in early life regulates gut-brain axis and relieves anxiety-like behavior in adulthood. Pharmacol. Res. 2022, 177, 106090. [Google Scholar] [CrossRef]
- Jhun, J.; Cho, K.-H.; Lee, D.-H.; Kwon, J.Y.; Woo, J.S.; Kim, J.; Na, H.S.; Park, S.-H.; Kim, S.J.; Cho, M.-L. Oral administration of Lactobacillus rhamnosus ameliorates the progression of osteoarthritis by inhibiting joint pain and inflammation. Cells 2021, 10, 1057. [Google Scholar] [CrossRef]
- Lin, Q. Submerged Fermentation of Lactobacillus Rhamnosus YS9 for γ-aminobutyric acid (GABA) production. Braz. J. Microbiol. 2013, 44, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Lorusso, A.; Coda, R.; Montemurro, M.; Rizzello, C.G. Use of selected lactic acid bacteria and quinoa flour for manufacturing novel yogurt-like beverages. Foods Basel Switz. 2018, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Slykerman, R.F.; Hood, F.; Wickens, K.; Thompson, J.M.D.; Barthow, C.; Murphy, R.; Kang, J.; Rowden, J.; Stone, P.; Crane, J.; et al. Effect of Lactobacillus rhamnosus HN001 in pregnancy on postpartum symptoms of depression and anxiety: A randomised double-blind placebo-controlled trial. EBioMedicine 2017, 24, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Knuesel, T.; Mohajeri, M.H. The role of the gut microbiota in the development and progression of major depressive and bipolar disorder. Nutrients 2021, 14, 37. [Google Scholar] [CrossRef]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Duranti, S.; Ruiz, L.; Lugli, G.A.; Tames, H.; Milani, C.; Mancabelli, L.; Mancino, W.; Longhi, G.; Carnevali, L.; Sgoifo, A.; et al. Bifidobacterium adolescentis as a key member of the human gut microbiota in the production of GABA. Sci. Rep. 2020, 10, 14112. [Google Scholar] [CrossRef]
- Han, M.; Liao, W.; Wu, S.; Gong, X.; Bai, C. Use of streptococcus thermophilus for the in situ production of γ-aminobutyric acid-enriched fermented milk. J. Dairy Sci. 2020, 103, 98–105. [Google Scholar] [CrossRef] [Green Version]
- Franciosi, E.; Carafa, I.; Nardin, T.; Schiavon, S.; Poznanski, E.; Cavazza, A.; Larcher, R.; Tuohy, K.M. Biodiversity and γ-aminobutyric acid production by lactic acid bacteria isolated from traditional alpine raw cow’s milk cheeses. BioMed Res. Int. 2015, 2015, 625740. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Zhao, Z.; Zhao, Y.; Wang, Z.; Wang, C.; Yang, G.; Li, S. Lactobacillus plantarum DP189 Prevents cognitive dysfunction in D-Galactose/AlCl3 induced mouse model of Alzheimer’s disease via modulating gut microbiota and PI3K/Akt/GSK-3β signaling pathway. Nutr. Neurosci. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhang, S.; Zhang, X.; Zhou, H.; Wen, T.; Wang, J. Depression-like symptoms due to Dcf1 deficiency are alleviated by intestinal transplantation of Lactobacillus murine and lactobacillus reuteri. Biochem. Biophys. Res. Commun. 2022, 593, 137–143. [Google Scholar] [CrossRef]
- Sharafi, S.; Nateghi, L. Optimization of gamma-aminobutyric acid production by probiotic bacteria through response surface methodology. Iran. J. Microbiol. 2020, 12, 584–591. [Google Scholar] [CrossRef]
- Kelly, J.R.; Allen, A.P.; Temko, A.; Hutch, W.; Kennedy, P.J.; Farid, N.; Murphy, E.; Boylan, G.; Bienenstock, J.; Cryan, J.F.; et al. Lost in Translation? The potential psychobiotic Lactobacillus Rhamnosus (JB-1) fails to modulate stress or cognitive performance in healthy male subjects. Brain Behav. Immun. 2017, 61, 50–59. [Google Scholar] [CrossRef]
- Bercik, P.; Verdu, E.F.; Foster, J.A.; Macri, J.; Potter, M.; Huang, X.; Malinowski, P.; Jackson, W.; Blennerhassett, P.; Neufeld, K.A.; et al. Chronic gastrointestinal inflammation induces anxiety-like behavior and alters central nervous system biochemistry in mice. Gastroenterology 2010, 139, 2102–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Genera | Species | Physiological Function | Therapeutic Agency | References |
|---|---|---|---|---|
| Lactobacillus | Lactobacillus rhamnosus | Intestinal flora regulation Antibacterial compound production Neurotransmitter modulation Immune response regulation | Depression | [46,47,48,49] |
| Obesity | ||||
| Lactobacillus reuteri | Immune response regulation Intestinal barrier function Intestinal, oral and vaginal flora regulation | Obesity Gastrointestinal disorders Acute infectious diarrhea | [47,48] | |
| Bifidobacterium | Bifidobacterium longum | Immune response regulation Carbohydrate metabolism in infants Lactose metabolism | Acute rotavirus Diarrhea Antibiotic-associated diarrhea Lactose intolerance | [50,51] |
| Bifidobacterium breve | Pathogen growt hinhibition Immune response regulation Intestinal flora regulation Lipid metabolism | Celiac disease Obesity Necrotizing enterocolitis | [50,52] | |
| Streptococcus | Streptococcus thermophilus | Immune response regulation Lactose metabolism Intestinal toxin regulation Immune response regulation | Lactose intolerance Uremia | [53,54] |
| Bacillus | Bacillus coagulans | Lactose metabolism Protein and carbohydrate metabolism Toxin regulation Bowel motility Cholesterol metabolism Intestinal flora regulation | Irritable bowel syndrome Antibiotic-associated diarrhea Obesity Inflammatory bowel disease Colorectal cancer | [47,55] |
| Saccharomyces | Saccharomyces cerevisiae | Immune response regulation Pathogen growth inhibition | Inflammatory bowel disease Anti-biotic associated diarrhea Vaginal infection | [47,56] |
| Enterococcus | Enterococcus faecium | Cholesterol modulation Immune response regulation Intestinal flora regulation | Intestinal disease Anti-biotic associated diarrhea Irritable bowel syndrome | [56,57] |
| Escherichia | Escherichia coli | Intestinal barrier Immune response regulation | Intestinal inflammatory diseaseInfectious diarrhea | [58,59] |
| Genera | GABA-Producing Species/Strains | Probiotic GABAergic Effects/Correlations | References |
|---|---|---|---|
| Lactobacillus | Lactobacillus | ↓ Depression | [4,20,21] |
| rhamnosus JB-1 | |||
| Lactobacillus | ↓ Anxiety-like | [64,65] | |
| rhamnosus GG | Behavior | ||
| Lactobacillus | ↓ Osteoarthritis pain | [66] | |
| rhamnosus LR-2 | Severity | ||
| Lactobacillus | N/A (In vitro) | [67] | |
| rhamnosus YS9 | |||
| Lactobacillus rhamnosus SP1 | N/A (In vitro) | [68] | |
| Lactobacillus plantarum T6B10 | N/A (In vitro) | [68] | |
| L. plantarum DP189 | ↓ Gut dysbiosis, | ||
| ↓ Cognitive dysfunction in Alzheimer’s disease | [75] | ||
| Lactobacillus reuteri | ↓ Depression | [76] | |
| Lactobacillus murine | ↓ Depression | [76] | |
| Lactobacillus | ↑ Insulin secretion, | ||
| brevis DPC6108 | ↓ Metabolic | [22,71] | |
| Dysfunction, | |||
| ↓ Depression-like behavior | |||
| Lactobacillus | ↑ Insulin secretion, | [22,71] | |
| brevis DSM32386 | ↓ Metabolic | ||
| Dysfunction, | |||
| Lactobacillus | ↓ Depression-like behavior | [77] | |
| brevis TD10 | N/A (in vitro) | ||
| Bifidobacterium | Bifidobacterium | N/A (in vitro) | [71] |
| dentium NCFB 2243 | |||
| Bifidobacterium | N/A (in vitro) | [71] | |
| dentium DPC6333 | |||
| Bifidobacterium infantis UCC35624 | N/A (in vitro) | [71] | |
| Bifidobacterium | N/A | [72] | |
| adolescentis PRL2019 | |||
| Bifidobacterium adolescentis HD17T2H | N/A | [72] | |
| Bifidobacterium adolescentis DPC6044 | N/A (in vitro) | [71] | |
| Streptococcus | Streptococcus thermophilus GABA | N/A (in vitro) | [73,74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tette, F.-M.; Kwofie, S.K.; Wilson, M.D. Therapeutic Anti-Depressant Potential of Microbial GABA Produced by Lactobacillus rhamnosus Strains for GABAergic Signaling Restoration and Inhibition of Addiction-Induced HPA Axis Hyperactivity. Curr. Issues Mol. Biol. 2022, 44, 1434-1451. https://doi.org/10.3390/cimb44040096
Tette F-M, Kwofie SK, Wilson MD. Therapeutic Anti-Depressant Potential of Microbial GABA Produced by Lactobacillus rhamnosus Strains for GABAergic Signaling Restoration and Inhibition of Addiction-Induced HPA Axis Hyperactivity. Current Issues in Molecular Biology. 2022; 44(4):1434-1451. https://doi.org/10.3390/cimb44040096
Chicago/Turabian StyleTette, Fernanda-Marie, Samuel K. Kwofie, and Michael D. Wilson. 2022. "Therapeutic Anti-Depressant Potential of Microbial GABA Produced by Lactobacillus rhamnosus Strains for GABAergic Signaling Restoration and Inhibition of Addiction-Induced HPA Axis Hyperactivity" Current Issues in Molecular Biology 44, no. 4: 1434-1451. https://doi.org/10.3390/cimb44040096
APA StyleTette, F. -M., Kwofie, S. K., & Wilson, M. D. (2022). Therapeutic Anti-Depressant Potential of Microbial GABA Produced by Lactobacillus rhamnosus Strains for GABAergic Signaling Restoration and Inhibition of Addiction-Induced HPA Axis Hyperactivity. Current Issues in Molecular Biology, 44(4), 1434-1451. https://doi.org/10.3390/cimb44040096