
Corema album Leaves Mediate DNA Damage in Triple-Negative Breast Cancer Cells

 , ,
, , 
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Extract of C. album Leaves (ECAL)
2.3. Quantification of Total Phenolic Content
2.4. HPLC Analysis and Mass Spectrometry Analysis of Phenolic Content of ECAL
2.5. Cells Culture
2.6. Cell Viability
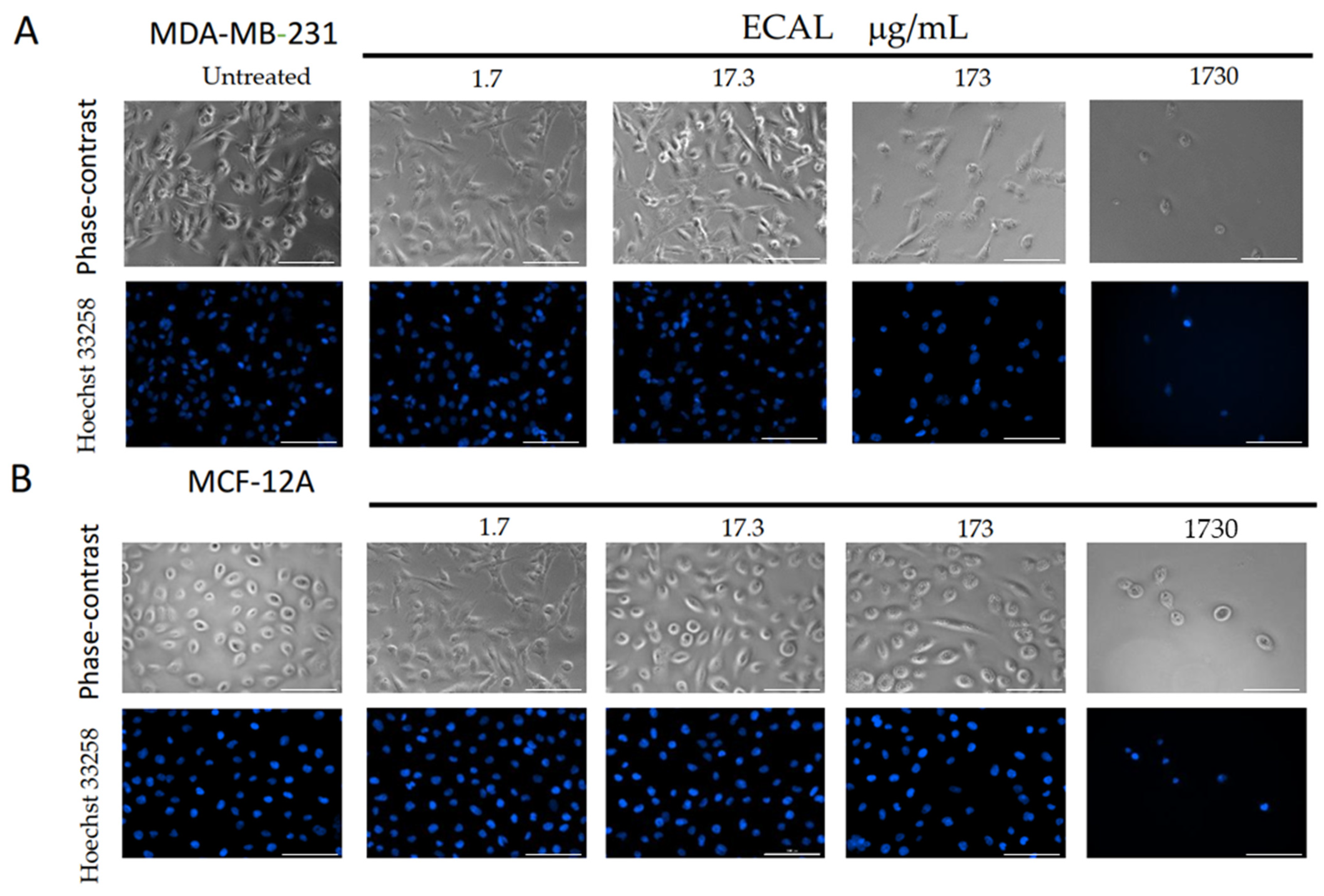
2.7. Cell Morphology
2.8. Tunel Assay
2.9. Western-Blot Analysis
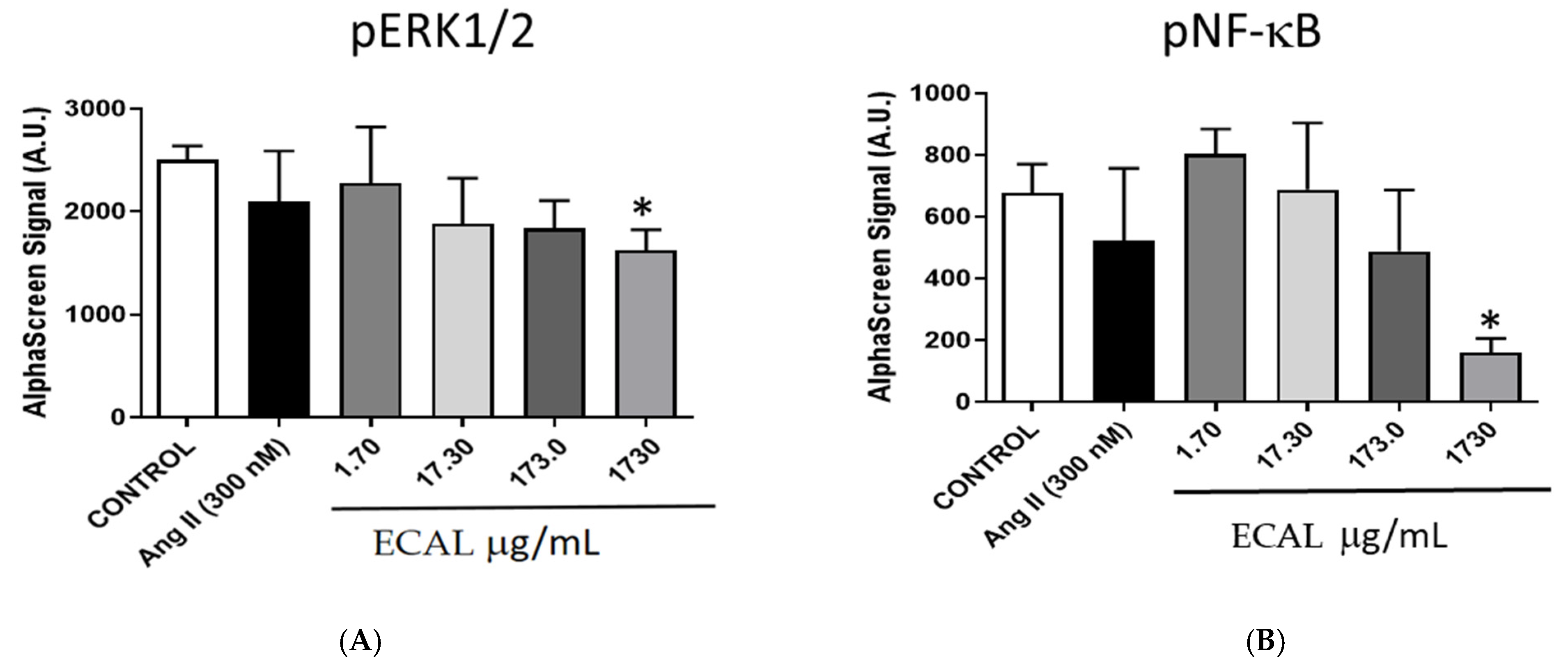
2.10. ERK1/2 Phosphorylation Assay
2.11. NF-κB Phosphorylation Assay
2.12. Statistical Analysis
3. Results
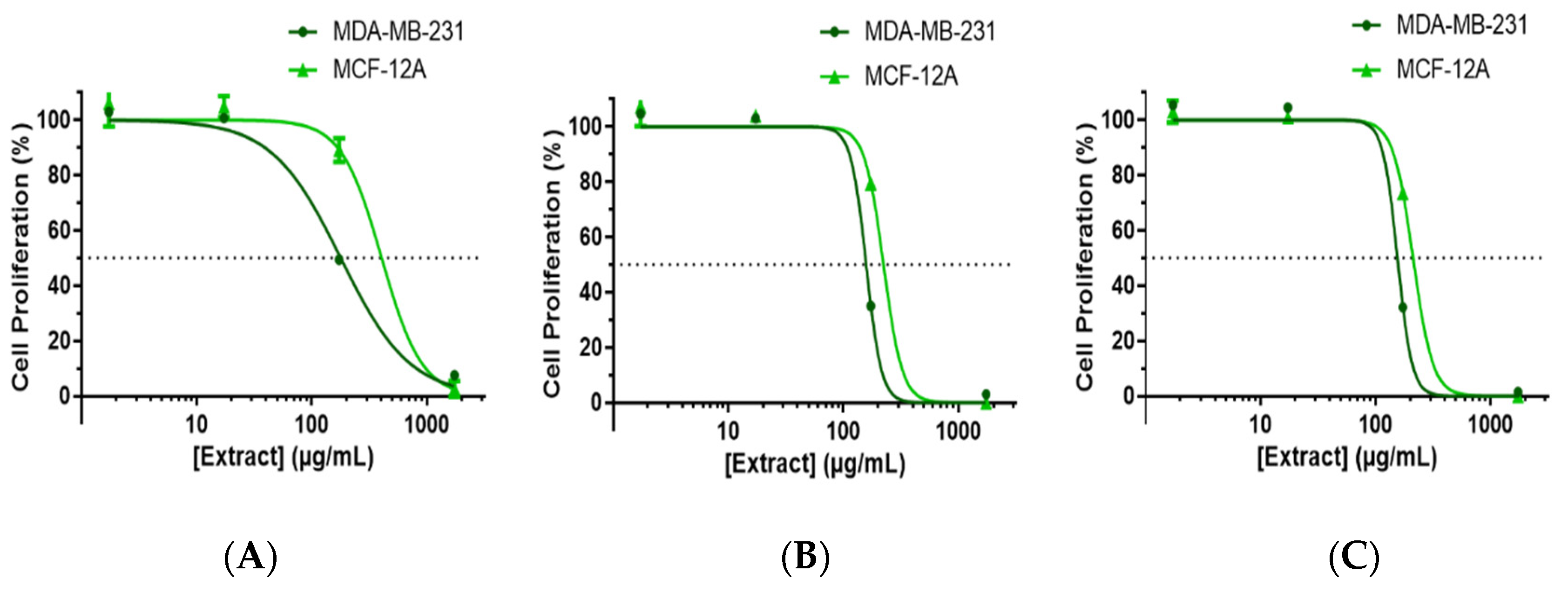
3.1. ECAL Supresses the Proliferation of Triple-Negative Breast Cancer
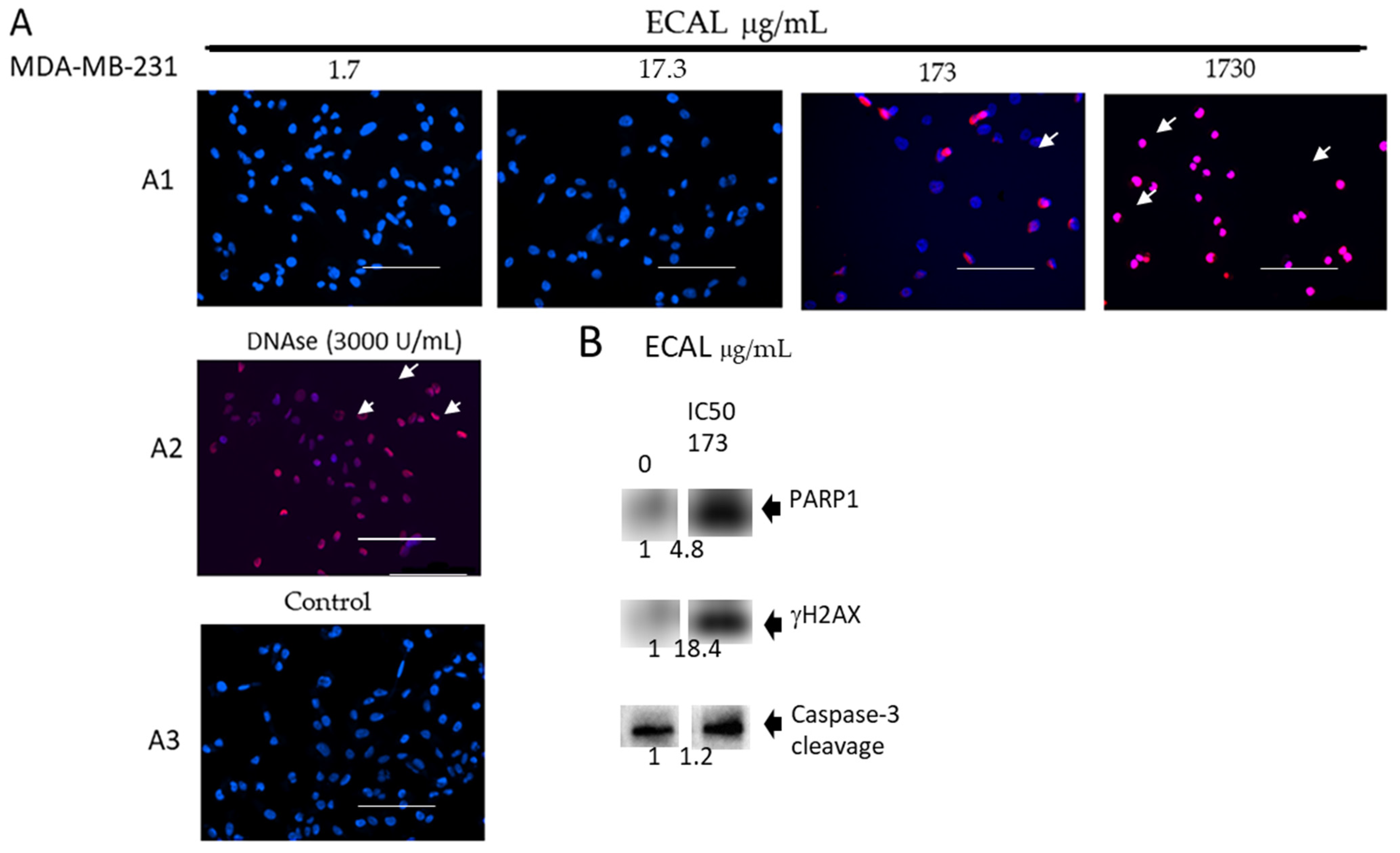
3.2. ECAL Induced Double-Stranded DNA Damage and Apoptosis via Caspase 3
3.3. Polyphenolic Content of ECAL
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S. Triple-Negative Breast Cancer Molecular Subtyping and Treatment Progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Siddharth, S.; Sharma, D. Triple Negative Breast Cancer: A Mountain Yet to Be Scaled Despite the Triumphs. Cancers 2021, 13, 3697. [Google Scholar] [CrossRef]
- Candelaria, R.P.; Adrada, B.E.; Wei, W.; Thompson, A.M.; Santiago, L.; Lane, D.L.; Huang, M.L.; Arribas, E.M.; Rauch, G.M.; Symmans, W.F.; et al. Imaging Features of Triple-Negative Breast Cancers According to Androgen Receptor Status. Eur. J. Radiol. 2019, 114, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A.; Bermúdez-Cruz, R.M. Natural Compounds that Target DNA Repair Pathways and Their Therapeutic Potential to Counteract Cancer Cells. Front. Oncol. 2020, 10, 2567. [Google Scholar] [CrossRef] [PubMed]
- Andrade, S.C.; Guiné, R.P.F.; Gonçalves, F.J.A. Evaluation of Phenolic Compounds, Antioxidant Activity and Bioaccessibility in White Crowberry (Corema album). J. Food Meas. Charact. 2017, 11, 1936–1946. [Google Scholar] [CrossRef]
- Gras, A.; Garnatje, T.; Ibáñez, N.; López-Pujol, J.; Nualart, N.; Vallès, J. Medicinal Plant Uses and Names from the Herbarium of Francesc Bolòs (1773–1844). J. Ethnopharmacol. 2017, 204, 142–168. [Google Scholar] [CrossRef]
- De Oliveira, P.B.; Dale, A. Corema album (L.) D. Don, the White Crowberry—A New Crop. J. Berry Res. 2012, 2, 123–133. [Google Scholar] [CrossRef]
- Moreira da Silva, A.; Barroca, M.J.; Guiné, R.P.F. Knowledge and Consumption Habits Related with White Crowberries (Corema album L.). Appl. Sci. 2021, 11, 5463. [Google Scholar] [CrossRef]
- León-González, A.J.; Mateos, R.; Ramos, S.; Martín, M.Á.; Sarriá, B.; Martín-Cordero, C.; López-Lázaro, M.; Bravo, L.; Goya, L. Chemo-Protective Activity and Characterization of Phenolic Extracts from Corema album. Food Res. Int. 2012, 49, 728–738. [Google Scholar] [CrossRef]
- Marques, J.; Martin, D.; Amado, A.M.; Lysenko, V.; Osório, N.; Batista de Carvalho, L.A.E.; Marques, M.P.M.; Barroca, M.J.; Moreira da Silva, A. Novel Insights into Corema album Berries: Vibrational Profile and Biological Activity. Plants 2021, 10, 1761. [Google Scholar] [CrossRef] [PubMed]
- León-González, A.J.; Truchado, P.; Tomás-Barberán, F.A.; López-Lázaro, M.; Barradas, M.C.D.; Martín-Cordero, C. Phenolic Acids, Flavonols and Anthocyanins in Corema album (L.) D. Don Berries. J. Food Compos. Anal. 2013, 29, 58–63. [Google Scholar] [CrossRef]
- León-González, A.J.; Manson, M.M.; López-Lizaro, M.; Navarro, I.; Martín-Cordero, C. Induction of Apoptosis and Cell Cycle Arrest in Human Colon Carcinoma Cells by Corema album Leaves. Nat. Prod. Commun. 2014, 9, 55–56. [Google Scholar] [CrossRef] [PubMed]
- León-González, A.J.; López-Lázaro, M.; Espartero, J.L.; Martín-Cordero, C. Cytotoxic Activity of Dihydrochalcones Isolated from Corema album Leaves against HT-29 Colon Cancer Cells. Nat. Prod. Commun. 2013, 8, 1255–1256. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, H.; Yang, S. Phenolic Compounds and Its Antioxidant Activities in Ethanolic Extracts from Seven Cultivars of Chinese Jujube. Food Sci. Hum. Wellness 2014, 3, 183–190. [Google Scholar] [CrossRef]
- Steinmetz, R.; Wagoner, H.A.; Zeng, P.; Hammond, J.R.; Hannon, T.S.; Meyers, J.L.; Pescovitz, O.H. Mechanisms Regulating the Constitutive Activation of the Extracellular Signal-Regulated Kinase (ERK) Signaling Pathway in Ovarian Cancer and the Effect of Ribonucleic Acid Interference for ERK1/2 on Cancer Cell Proliferation. Mol. Endocrinol. 2004, 18, 2570–2582. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Ravindran, J.; Aggarwal, B.B. NF-ΚB and Cancer: How Intimate Is This Relationship. Mol. Cell. Biochem. 2010, 336, 25–37. [Google Scholar] [CrossRef]
- Wang, M.; Chen, S.; Ao, D. Targeting DNA Repair Pathway in Cancer: Mechanisms and Clinical Application. MedComm 2021, 2, 654–691. [Google Scholar] [CrossRef]
- Oliveira, I.; Nunes, A.; Lima, A.; Borralho, P.; Rodrigues, C.; Ferreira, R.; Ribeiro, A. New Lectins from Mediterranean Flora. Activity against HT29 Colon Cancer Cells. Int. J. Mol. Sci. 2019, 20, 3059. [Google Scholar] [CrossRef]
- Brito, C.; Bertotti, T.; Primitivo, M.J.; Neves, M.; Pires, C.L.; Cruz, P.F.; Martins, P.A.T.; Rodrigues, A.C.; Moreno, M.J.; Brito, R.M.M.; et al. Corema album Spp: Edible Wild Crowberries with a High Content in Minerals and Organic Acids. Food Chem. 2021, 345, 128732. [Google Scholar] [CrossRef]
- Fresco, P.; Borges, F.; Marques, M.; Diniz, C. The Anticancer Properties of Dietary Polyphenols and Its Relation with Apoptosis. Curr. Pharm. Des. 2010, 16, 114–134. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, A.; Briand, C.; Grütter, M.G. Crystal Structure of Caspase-2, Apical Initiator of the Intrinsic Apoptotic Pathway. J. Biol. Chem. 2003, 278, 42441–42447. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, N.; Crown, J.; Stunell, H.; Hill, A.D.K.; McDermott, E.; O’Higgins, N.; Duffy, M.J. Caspase 3 in Breast Cancer. Clin. Cancer Res. 2003, 9, 738–742. [Google Scholar] [PubMed]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase Functions in Cell Death and Disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef]
- Sullivan, R.J.; Infante, J.R.; Janku, F.; Wong, D.J.L.; Sosman, J.A.; Keedy, V.; Patel, M.R.; Shapiro, G.I.; Mier, J.W.; Tolcher, A.W.; et al. First-in-Class ERK1/2 Inhibitor Ulixertinib (BVD-523) in Patients with MAPK Mutant Advanced Solid Tumors: Results of a Phase I Dose-Escalation and Expansion Study. Cancer Discov. 2018, 8, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Chang, P.; Lippard, S.J. Recognition of Platinum−DNA Damage by Poly(ADP-Ribose) Polymerase-1. Biochemistry 2010, 49, 6177–6183. [Google Scholar] [CrossRef]
- Ikeda, M.; Kurose, A.; Takatori, E.; Sugiyama, T.; Traganos, F.; Darzynkiewicz, Z.; Sawai, T. DNA Damage Detected with ΓH2AX in Endometrioid Adenocarcinoma Cell Lines. Int. J. Oncol. 2010, 36, 1081–1088. [Google Scholar] [CrossRef]
- Kowalczyk, T.; Sitarek, P.; Skała, E.; Toma, M.; Wielanek, M.; Pytel, D.; Wieczfińska, J.; Szemraj, J.; Śliwiński, T. Induction of Apoptosis by in Vitro and in Vivo Plant Extracts Derived from Menyanthes trifoliata L. in Human Cancer Cells. Cytotechnology 2019, 71, 165–180. [Google Scholar] [CrossRef]
- Fouzat, A.; Hussein, O.J.; Gupta, I.; Al-Farsi, H.F.; Khalil, A.; Al Moustafa, A.-E. Elaeagnus Angustifolia Plant Extract Induces Apoptosis via P53 and Signal Transducer and Activator of Transcription 3 Signaling Pathways in Triple-Negative Breast Cancer Cells. Front. Nutr. 2022, 9, 871667. [Google Scholar] [CrossRef]
- Karimi, M.; Conserva, F.; Mahmoudi, S.; Bergman, J.; Wiman, K.G.; Bykov, V.J.N. Extract from Asteraceae Brachylaena Ramiflora Induces Apoptosis Preferentially in Mutant P53-Expressing Human Tumor Cells. Carcinogenesis 2010, 31, 1045–1053. [Google Scholar] [CrossRef]
- Efenberger-Szmechtyk, M.; Nowak, A.; Nowak, A. Cytotoxic and Dna-Damaging Effects of Aronia Melanocarpa, Cornus Mas, and Chaenomeles Superba Leaf Extracts on the Human Colon Adenocarcinoma Cell Line Caco-2. Antioxidants 2020, 9, 1030. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.M.; Salvador, M.R.; Machado, C.S.; Lang, K.L. In Vitro Evaluation of the Cytotoxic and Genotoxic Effects of Ethanolic Extracts of Echinodorus Macrophyllus. Braz. J. Health Pharm. 2021, 3, 12–21. [Google Scholar] [CrossRef]
- Pal, A.; Sengupta, S.; Kundu, R. Tiliacora Racemosa Leaves Induce Oxidative Stress Mediated DNA Damage Leading to G2/M Phase Arrest and Apoptosis in Cervical Cancer Cells SiHa. J. Ethnopharmacol. 2021, 269, 113686. [Google Scholar] [CrossRef] [PubMed]
- Kühnel, F.; Zender, L.; Paul, Y.; Tietze, M.K.; Trautwein, C.; Manns, M.; Kubicka, S. NFκB Mediates Apoptosis through Transcriptional Activation of Fas (CD95) in Adenoviral Hepatitis. J. Biol. Chem. 2000, 275, 6421–6427. [Google Scholar] [CrossRef]
- Altieri, D.C. Targeting Survivin in Cancer. Cancer Lett. 2013, 332, 225–228. [Google Scholar] [CrossRef]
- Jin, R.; Yamashita, H.; Yu, X.; Wang, J.; Franco, O.E.; Wang, Y.; Hayward, S.W.; Matusik, R.J. Inhibition of NF-Kappa B Signaling Restores Responsiveness of Castrate-Resistant Prostate Cancer Cells to Anti-Androgen Treatment by Decreasing Androgen Receptor-Variant Expression. Oncogene 2015, 34, 3700–3710. [Google Scholar] [CrossRef]
- Abbaspour Babaei, M.; Zaman Huri, H.; Kamalidehghan, B.; Yeap, S.K.; Ahmadipour, F. Apoptotic Induction and Inhibition of NF-ΚB Signaling Pathway in Human Prostatic Cancer PC3 Cells by Natural Compound 2,2′-Oxybis (4-Allyl-1-Methoxybenzene), Biseugenol B, from Litsea Costalis: An in Vitro Study. Onco. Targets. Ther. 2017, 10, 277–294. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Cusack, J.C.; Liu, R.; Baldwin, A.S. Control of Inducible Chemoresistance: Enhanced Anti-Tumor Therapy through Increased Apoptosis by Inhibition of NF-ΚB. Nat. Med. 1999, 5, 412–417. [Google Scholar] [CrossRef]
- Antoon, J.W.; White, M.D.; Slaughter, E.M.; Driver, J.L.; Khalili, H.S.; Elliott, S.; Smith, C.D.; Burow, M.E.; Beckman, B.S. Targeting NFĸB Mediated Breast Cancer Chemoresistance through Selective Inhibition of Sphingosine Kinase-2. Cancer Biol. Ther. 2011, 11, 678–689. [Google Scholar] [CrossRef]
- Roth, B.J.; Dreicer, R.; Einhorn, L.H.; Neuberg, D.; Johnson, D.H.; Smith, J.L.; Hudes, G.R.; Schultz, S.M.; Loehrer, P.J. Significant Activity of Paclitaxel in Advanced Transitional-Cell Carcinoma of the Urothelium: A Phase II Trial of the Eastern Cooperative Oncology Group. J. Clin. Oncol. 1994, 12, 2264–2270. [Google Scholar] [CrossRef]
- Das, R.; Mehta, D.K.; Dhanawat, M. Medicinal Plants in Cancer Treatment: Contribution of Nuclear Factor-Kappa B (NF-KB) Inhibitors. Mini-Rev. Med. Chem. 2022, 22, 1938–1962. [Google Scholar] [CrossRef] [PubMed]
- Weaver, A.N.; Yang, E.S. Beyond DNA Repair: Additional Functions of PARP-1 in Cancer. Front. Oncol. 2013, 3, 290. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, B.; Liu, Y.; Yu, X.; Cheng, G. Dual Effects of Active ERK in Cancer: A Potential Target for Enhancing Radiosensitivity (Review). Oncol. Lett. 2020, 20, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Mao, R.; Wang, J.; Ding, L.; Jiang, S.; Gao, C.; Kang, H.; Chen, X.; Sun, X.; Xu, J. ERK1/2 Promoted Proliferation and Inhibited Apoptosis of Human Cervical Cancer Cells and Regulated the Expression of c-Fos and c-Jun Proteins. Med. Oncol. 2015, 32, 57. [Google Scholar] [CrossRef]
- Zhou, G.; Yang, J.; Song, P. Correlation of ERK/MAPK Signaling Pathway with Proliferation and Apoptosis of Colon Cancer Cells. Oncol. Lett. 2018, 17, 2266–2270. [Google Scholar] [CrossRef]
- Yan, Z.; Ohuchida, K.; Fei, S.; Zheng, B.; Guan, W.; Feng, H.; Kibe, S.; Ando, Y.; Koikawa, K.; Abe, T.; et al. Inhibition of ERK1/2 in Cancer-Associated Pancreatic Stellate Cells Suppresses Cancer-Stromal Interaction and Metastasis. J. Exp. Clin. Cancer Res. 2019, 38, 221. [Google Scholar] [CrossRef]
- Gkouveris, I.; Nikitakis, N.; Karanikou, M.; Rassidakis, G.; Sklavounou, A. Erk1/2 Activation and Modulation of STAT3 Signaling in Oral Cancer. Oncol. Rep. 2014, 32, 2175–2182. [Google Scholar] [CrossRef]
- Chin, H.M.; Lai, D.K.; Falchook, G.S. Extracellular Signal-Regulated Kinase (ERK) Inhibitors in Oncology Clinical Trials. J. Immunother. Precis. Oncol. 2019, 2, 10–16. [Google Scholar] [CrossRef]
- Liu, F.; Yang, X.; Geng, M.; Huang, M. Targeting ERK, an Achilles’ Heel of the MAPK Pathway, in Cancer Therapy. Acta Pharm. Sin. B 2018, 8, 552–562. [Google Scholar] [CrossRef]
- León-González, A.J.; Navarro, I.; Acero, N.; Muñoz-Mingarro, D.; Martín-Cordero, C. The Fruit of Corema album (L.) D. Don, a Singular White Berry with Potential Benefits in Nutrition and Health. Phytochem. Rev. 2022, 21, 525–536. [Google Scholar] [CrossRef]
- Macedo, D.; Tavares, L.; McDougall, G.J.; Vicente Miranda, H.; Stewart, D.; Ferreira, R.B.; Tenreiro, S.; Outeiro, T.F.; Santos, C.N. (Poly)Phenols Protect from α-Synuclein Toxicity by Reducing Oxidative Stress and Promoting Autophagy. Hum. Mol. Genet. 2015, 24, 1717–1732. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Xue, L. Kaempferol Suppresses Proliferation and Induces Cell Cycle Arrest, Apoptosis, and DNA Damage in Breast Cancer Cells. Oncol. Res. 2019, 27, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Fresco, P.; Borges, F.; Diniz, C.; Marques, M.P.M. New Insights on the Anticancer Properties of Dietary Polyphenols. Med. Res. Rev. 2006, 26, 747–766. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Compound | MW |
|---|---|---|
| Hydroxybenzoic acids | 4-Hydroxybenzoic acid hexoside | 300 |
| Protocatechuic acid hexoside | 316 | |
| Hydroxycinnamic acids | Coumaroyl glucose | 326 |
| Sinapoyl glucose | 386 | |
| Vanillyl glucose | 330 | |
| Feruloyl glucose | 356 | |
| Flavanols | (+)-Catechin | 29,068 |
| (−)-Epicatechin | 290,268 | |
| (+)-Catechin 3-O-glucose | 452,409 | |
| Procyanidin dimer type A | 576 | |
| Procyanidin trimer type A | 864 | |
| Procyanidin trimer type A | 862 | |
| Procyanidin tetramer type A | 1152 | |
| Procyanidin tetramer type A | 1150 | |
| Procyanidin pentamer type A | 1440 | |
| Procyanidin galhate | 880 | |
| Flavones or Flavonols | Myricetin hexoside | 480 |
| Myricetin dihexoside | 642 | |
| Rhamnetin hexoside | 478 | |
| Quercetin rhamnosyl hexoside | 610 | |
| Quercetin hexoside | 464 | |
| Kaempherol hexoside | 448 | |
| Myricetin methyl ether hexoside | 494 | |
| Myricetin xyloside | 450 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerquido, A.S.; Vojtek, M.; Ribeiro-Oliveira, R.; Gonçalves-Monteiro, S.; Barroca, M.J.; Moreira da Silva, A.; Viegas, O.; Freitas, V.; Sousa, J.B.; Ferreira, I.M.P.L.V.O.; et al. Corema album Leaves Mediate DNA Damage in Triple-Negative Breast Cancer Cells. Curr. Issues Mol. Biol. 2022, 44, 3598-3610. https://doi.org/10.3390/cimb44080246
Cerquido AS, Vojtek M, Ribeiro-Oliveira R, Gonçalves-Monteiro S, Barroca MJ, Moreira da Silva A, Viegas O, Freitas V, Sousa JB, Ferreira IMPLVO, et al. Corema album Leaves Mediate DNA Damage in Triple-Negative Breast Cancer Cells. Current Issues in Molecular Biology. 2022; 44(8):3598-3610. https://doi.org/10.3390/cimb44080246
Chicago/Turabian StyleCerquido, Ana Sofia, Martin Vojtek, Rita Ribeiro-Oliveira, Salomé Gonçalves-Monteiro, Maria João Barroca, Aida Moreira da Silva, Olga Viegas, Victor Freitas, Joana Beatriz Sousa, Isabel M. P. L. V. O. Ferreira, and et al. 2022. "Corema album Leaves Mediate DNA Damage in Triple-Negative Breast Cancer Cells" Current Issues in Molecular Biology 44, no. 8: 3598-3610. https://doi.org/10.3390/cimb44080246
APA StyleCerquido, A. S., Vojtek, M., Ribeiro-Oliveira, R., Gonçalves-Monteiro, S., Barroca, M. J., Moreira da Silva, A., Viegas, O., Freitas, V., Sousa, J. B., Ferreira, I. M. P. L. V. O., & Diniz, C. (2022). Corema album Leaves Mediate DNA Damage in Triple-Negative Breast Cancer Cells. Current Issues in Molecular Biology, 44(8), 3598-3610. https://doi.org/10.3390/cimb44080246