
PADI4 Haplotypes Contribute to mRNA Expression, the Enzymatic Activity of Peptidyl Arginine Deaminase and Rheumatoid Arthritis Risk in Patients from Western Mexico
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. RA Patients and Control Subjects
2.2. Laboratory Assessment and Quantification of Qutoantibodies
2.3. PADI4 SNPs Genotyping
2.4. RNA Isolation and cDNA Synthesis
2.5. Quantification of PADI4 Allelic Contribution to the mRNA Expression of PADI4
2.6. Quantitative Real-Time PCR for PADI4 Expression Quantitative
2.7. Isolation of Leukocytes
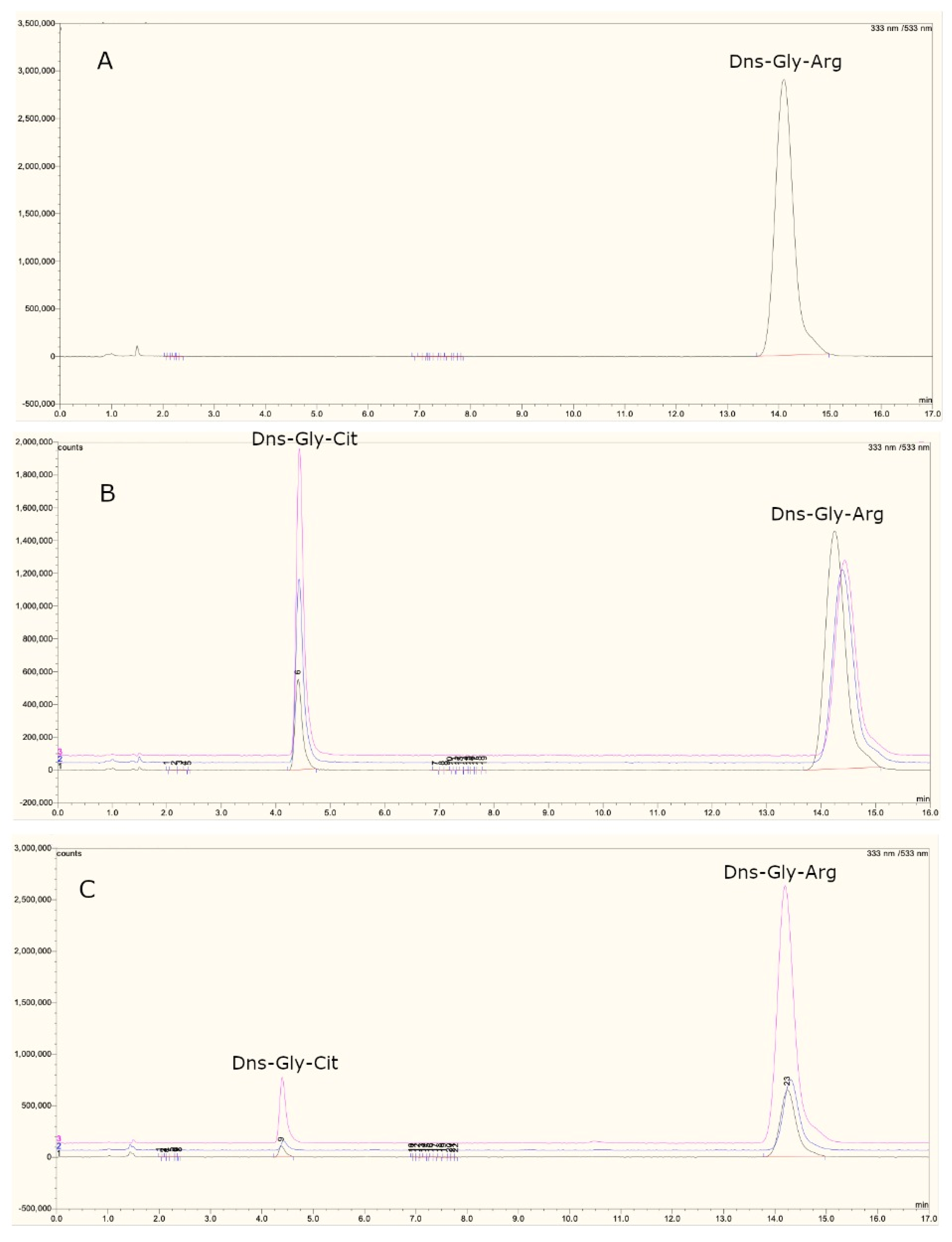
2.8. PAD4 Enzymatic Activity Analysis
2.9. Statistical Analysis
3. Results
3.1. Clinical and Demographic Characteristics of RA Patients
3.2. Genotype and Haplotype Frequencies of the PADI4 Gene in RA Patients and CS
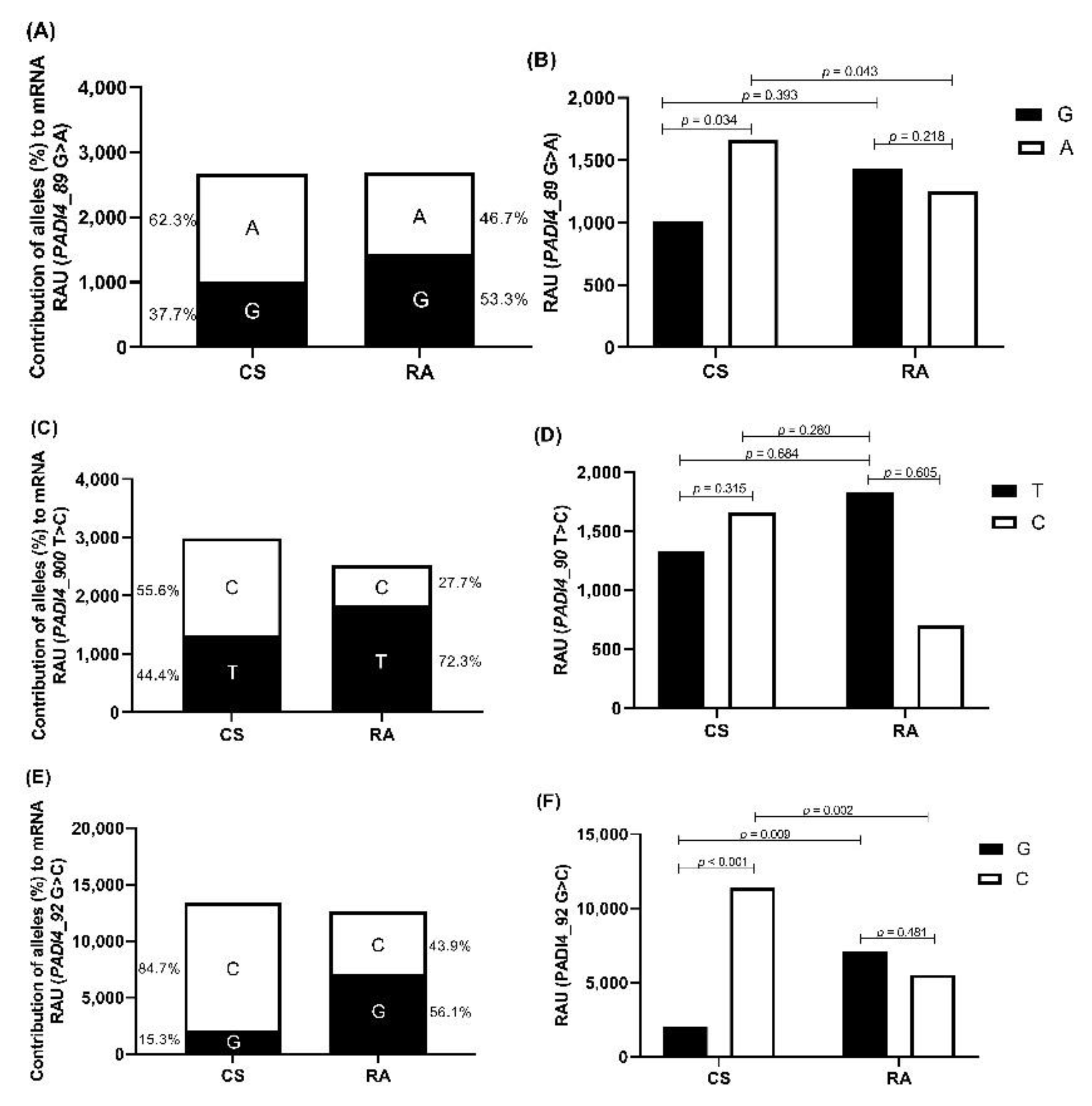
3.3. Allele-Specific Quantification of PADI4 mRNA Expression
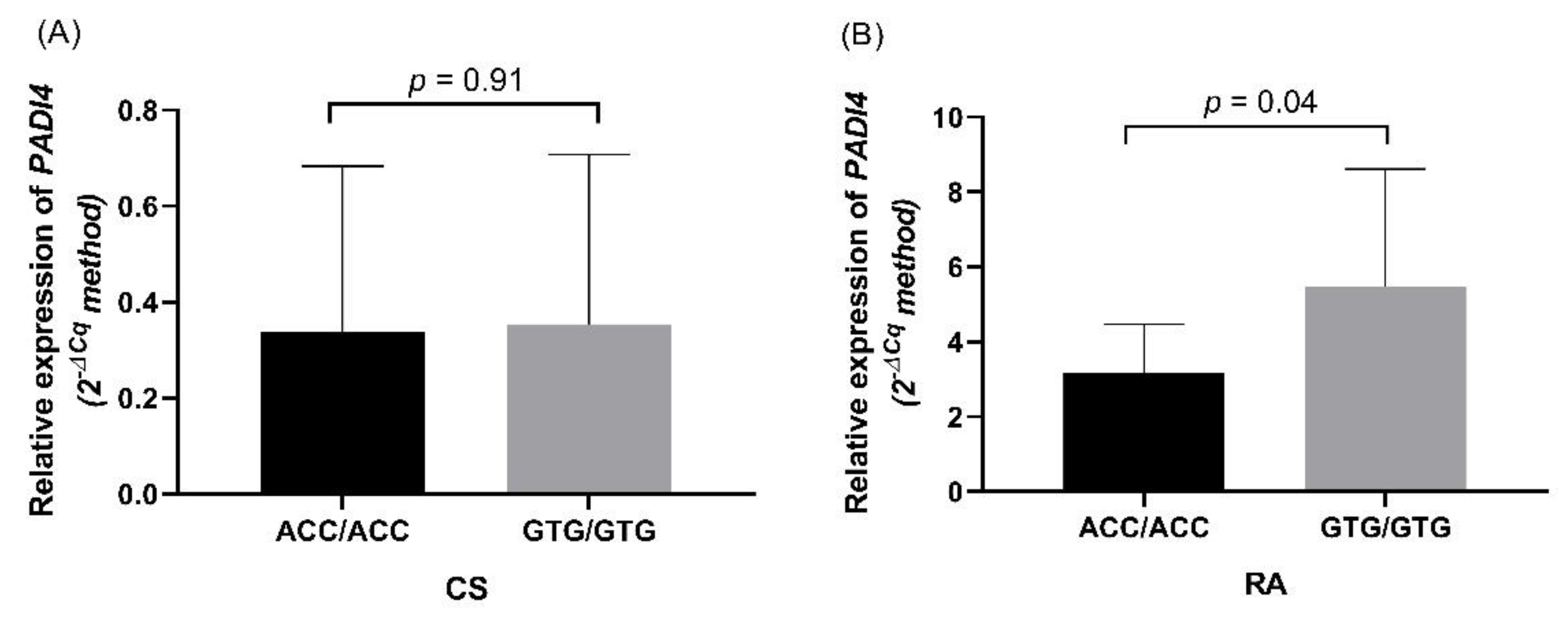
3.4. PADI4 Expression in RA Patients and CS
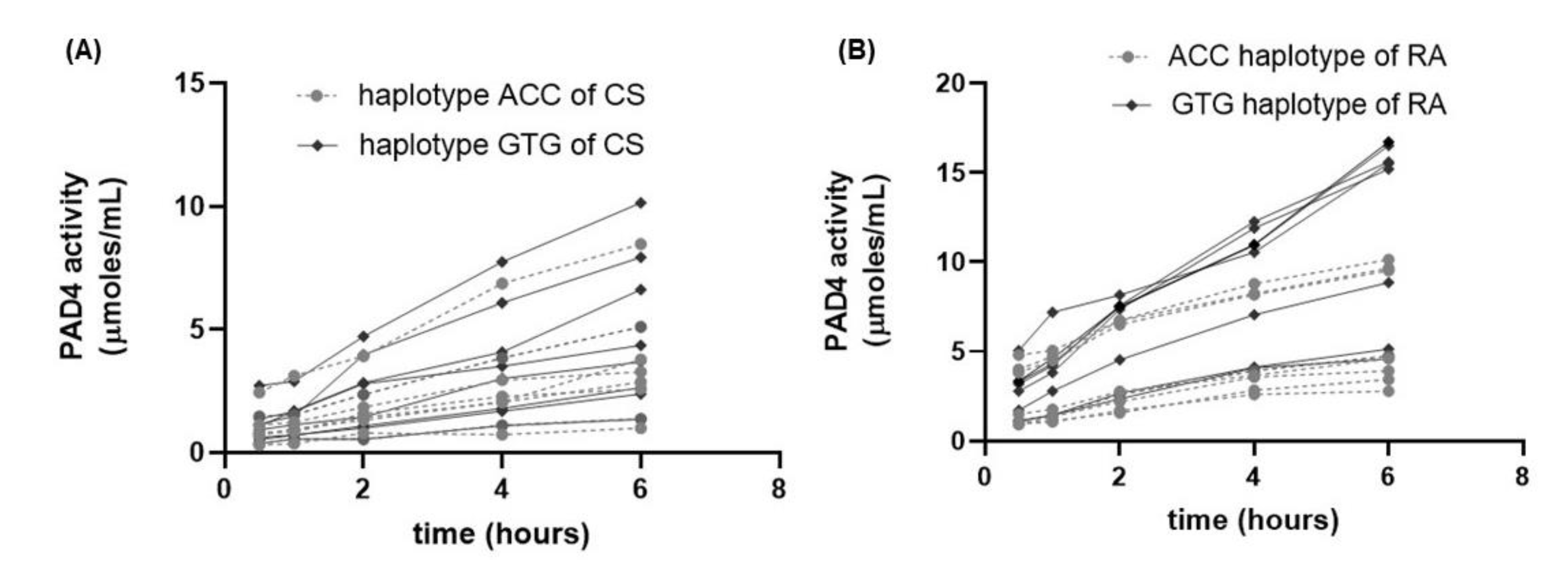
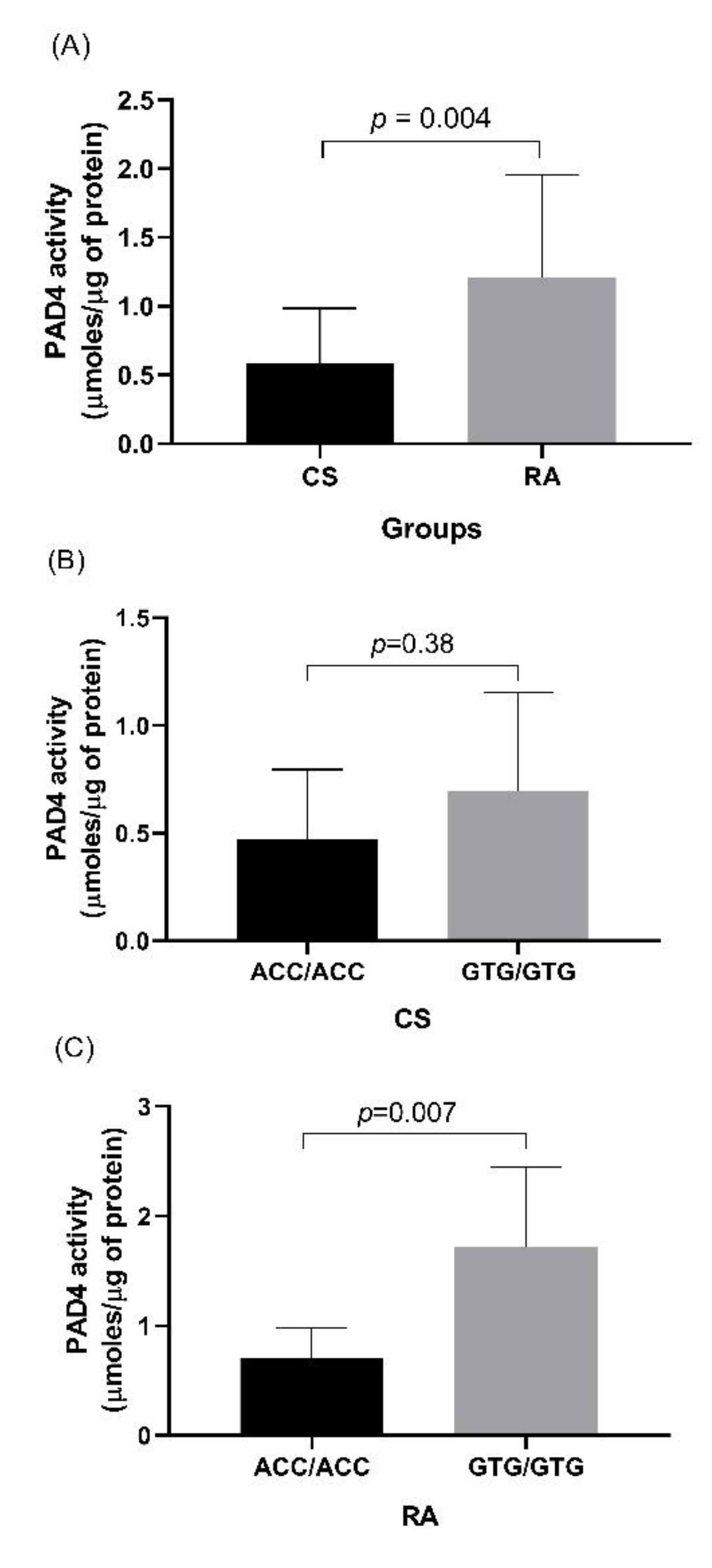
3.5. Evaluation of PAD4 Activity in CS and RA Patients
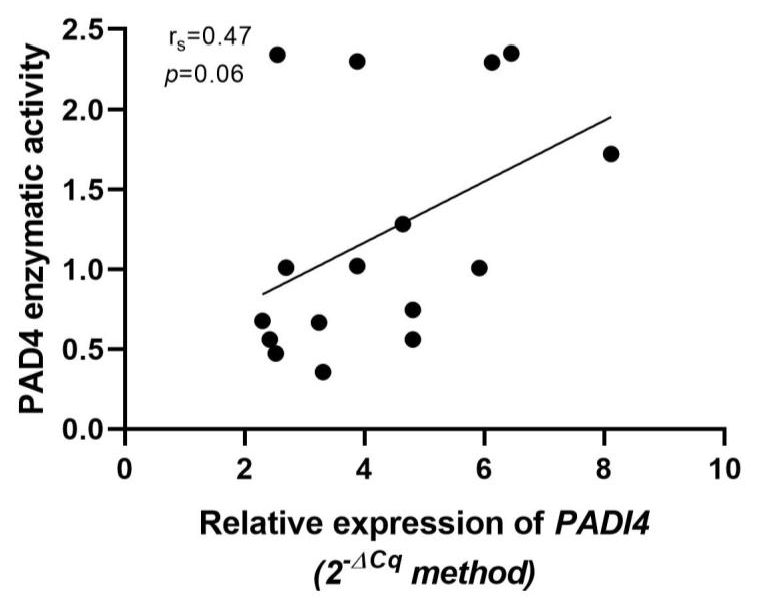
3.6. Association of Relative Expression of PADI4 Gene with PAD4 Enzymatic Activity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Firestein, G.S.; McInnes, I.B. Immunopathogenesis of Rheumatoid Arthritis. Immunity 2017, 46, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Hagiwara, T.; Yamada, M. Nuclear Localization of Peptidylarginine Deiminase V and Histone Deimination in Granulocytes. J. Biol. Chem. 2002, 277, 49562–49568. [Google Scholar] [CrossRef] [PubMed]
- Vossenaar, E.R. Expression and Activity of Citrullinating Peptidylarginine Deiminase Enzymes in Monocytes and Macrophages. Ann. Rheum. Dis. 2004, 63, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, H.; Sehnert, B.; Bockermann, R.; Engström, A.; Kalden, J.R.; Holmdahl, R. Humoral Immune Response to Citrullinated Collagen Type II Determinants in Early Rheumatoid Arthritis. Eur. J. Immunol. 2005, 35, 1643–1652. [Google Scholar] [CrossRef]
- Kinloch, A.; Tatzer, V.; Wait, R.; Peston, D.; Lundberg, K.; Donatien, P.; Moyes, D.; Taylor, P.C.; Venables, P.J. Identification of citrullinated α-enolase as a candidate autoantigen in rheumatoid arthritis. Arthritis Res. 2005, 7, R1421. [Google Scholar] [CrossRef]
- Pratesi, F.; Dioni, I.; Tommasi, C.; Alcaro, M.C.; Paolini, I.; Barbetti, F.; Boscaro, F.; Panza, F.; Puxeddu, I.; Rovero, P.; et al. Antibodies from Patients with Rheumatoid Arthritis Target Citrullinated Histone 4 Contained in Neutrophils Extracellular Traps. Ann. Rheum. Dis. 2014, 73, 1414–1422. [Google Scholar] [CrossRef]
- Wright, H.L.; Moots, R.J.; Edwards, S.W. The Multifactorial Role of Neutrophils in Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2014, 10, 593–601. [Google Scholar] [CrossRef]
- Suzuki, A.; Yamada, R.; Chang, X.; Tokuhiro, S.; Sawada, T.; Suzuki, M.; Nagasaki, M.; Nakayama-Hamada, M.; Kawaida, R.; Ono, M.; et al. Functional Haplotypes of PADI4, Encoding Citrullinating Enzyme Peptidylarginine Deiminase 4, Are Associated with Rheumatoid Arthritis. Nat. Genet. 2003, 34, 395–402. [Google Scholar] [CrossRef]
- Slack, J.L.; Jones, L.E.; Bhatia, M.M.; Thompson, P.R. Autodeimination of Protein Arginine Deiminase 4 Alters Protein–Protein Interactions but Not Activity. Biochemistry 2011, 50, 3997–4010. [Google Scholar] [CrossRef]
- Ikari, K.; Kuwahara, M.; Nakamura, T.; Momohara, S.; Hara, M.; Yamanaka, H.; Tomatsu, T.; Kamatani, N. Association BetweenPADI4 and Rheumatoid Arthritis: A Replication Study. Arthritis Rheum. 2005, 52, 3054–3057. [Google Scholar] [CrossRef]
- Kang, C.P.; Lee, H.-S.; Ju, H.; Cho, H.; Kang, C.; Bae, S.-C. A Functional Haplotype of ThePADI4 Gene Associated with Increased Rheumatoid Arthritis Susceptibility in Koreans. Arthritis Rheum. 2006, 54, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Plenge, R.M.; Padyukov, L.; Remmers, E.F.; Purcell, S.; Lee, A.T.; Karlson, E.W.; Wolfe, F.; Kastner, D.L.; Alfredsson, L.; Altshuler, D.; et al. Replication of Putative Candidate-Gene Associations with Rheumatoid Arthritis in >4000 Samples from North America and Sweden: Association of Susceptibility with PTPN22, CTLA4, and PADI4. Am. J. Hum. Genet. 2005, 77, 1044–1060. [Google Scholar] [CrossRef] [PubMed]
- Baños-Hernández, C.J.; Navarro-Zarza, J.E.; Parra-Rojas, I.; Vázquez-Villamar, M.; Ramón Padilla-Gutiérrez, J.; Valle, Y.; Reyes-Castillo, Z.; Magdalena Torres-Carrillo, N.; García-Arellano, S.; Brennan-Bourdon, L.M.; et al. PADI4 Polymorphisms and the Functional Haplotype Are Associated with Increased Rheumatoid Arthritis Susceptibility: A Replication Study in a Southern Mexican Population. Hum. Immunol. 2017, 78, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Guzmán, I.P.; Reyes-Castillo, Z.; Muñoz-Barrios, S.; Ruiz-Noa, Y.; Martínez-Bonilla, G.E.; Parra-Rojas, I.; Palafox-Sánchez, C.A.; Muñoz-Valle, J.F. Polymorphisms and Functional Haplotype in PADI4: Further Evidence for Contribution on Rheumatoid Arthritis Susceptibility and Anti-Cyclic Citrullinated Peptide Antibodies in a Western Mexican Population. Immunol. Lett. 2015, 163, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Damgaard, D.; Senolt, L.; Nielsen, M.F.; Pruijn, G.J.; Nielsen, C.H. Demonstration of Extracellular Peptidylarginine Deiminase (PAD) Activity in Synovial Fluid of Patients with Rheumatoid Arthritis Using a Novel Assay for Citrullination of Fibrinogen. Arthritis Res. 2014, 16, 498. [Google Scholar] [CrossRef]
- Damgaard, D.; Senolt, L.; Nielsen, C.H. Increased Levels of Peptidylarginine Deiminase 2 in Synovial Fluid from Anti-CCP-Positive Rheumatoid Arthritis Patients: Association with Disease Activity and Inflammatory Markers. Rheumatology 2016, 55, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Prevoo, M.L.; van ’t Hof, M.A.; Kuper, H.H.; van Leeuwen, M.A.; van de Putte, L.B.; van Riel, P.L. Modified Disease Activity Scores That Include Twenty-Eight-Joint Counts. Development and Validation in a Prospective Longitudinal Study of Patients with Rheumatoid Arthritis. Arthritis Rheum. 1995, 38, 44–48. [Google Scholar] [CrossRef]
- Cardiel, M.H.; Abello-Banfi, M.; Ruiz-Mercado, R.; Alarcon-Segovia, D. How to Measure Health Status in Rheumatoid Arthritis in Non-English Speaking Patients: Validation of a Spanish Version of the Health Assessment Questionnaire Disability Index (Spanish HAQ-DI). Clin. Exp. Rheumatol. 1993, 11, 117–121. [Google Scholar]
- Kaijzel, E.; Bayley, J.-P.; van Krugten, M.; Smith, L.; van de Linde, P.; Bakker, A.; Breedveld, F.; Huizinga, T.; Verweij, C. Allele-Specific Quantification of Tumor Necrosis Factor α (TNF) Transcription and the Role of Promoter Polymorphisms in Rheumatoid Arthritis Patients and Healthy Individuals. Genes Immun. 2001, 2, 135–144. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chikuma, T.; Yamada, M.; Tsuda, A.; Yamamoto, M.; Nakashima, K.; Yajima, R.; Kato, T. A Highly Sensitive High-Performance Liquid Chromatography– Fluorometric Method for the Assay of Peptidylarginine Deiminase Activity. Anal. Biochem. 2000, 285, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Klarenbeek, N.B.; Kerstens, P.J.S.M.; Huizinga, T.W.J.; Dijkmans, B.A.C.; Allaart, C.F. Recent Advances in the Management of Rheumatoid Arthritis. BMJ 2010, 341, c6942. [Google Scholar] [CrossRef] [PubMed]
- Perricone, C.; Ceccarelli, F.; Valesini, G. An Overview on the Genetic of Rheumatoid Arthritis: A Never-Ending Story. Autoimmun. Rev. 2011, 10, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Silman, A.J.; Pearson, J.E. Epidemiology and genetics of rheumatoid arthritis. Arthritis Res. 2002, 4, S265. [Google Scholar] [CrossRef]
- Bang, S.-Y.; Lee, K.-H.; Cho, S.-K.; Lee, H.-S.; Lee, K.W.; Bae, S.-C. Smoking Increases Rheumatoid Arthritis Susceptibility in Individuals Carrying the HLA-DRB1 Shared Epitope, Regardless of RF and ACPA. Arthritis Rheum. 2010, 62, 369–377. [Google Scholar] [CrossRef]
- Fan, L.Y.; Wang, W.J.; Wang, Q.; Zong, M.; Yang, L.; Zhang, H.; Sun, L.S.; Lu, T.B.; Han, J. A Functional Haplotype and Expression of the PADI4 Gene Associated with Increased Rheumatoid Arthritis Susceptibility in Chinese. Tissue Antigens 2008, 72, 469–473. [Google Scholar] [CrossRef]
- Hung, H.-C.; Lin, C.-Y.; Liao, Y.-F.; Hsu, P.-C.; Tsay, G.J.; Liu, G.-Y. The Functional Haplotype of Peptidylarginine Deiminase IV (S55G, A82V and A112G) Associated with Susceptibility to Rheumatoid Arthritis Dominates Apoptosis of Acute T Leukemia Jurkat Cells. Apoptosis 2007, 12, 475–487. [Google Scholar] [CrossRef]
- Jonsson, M.; Kantyka, T.; Falkowski, K.; Aliko, A.; Aga, A.; Lillegraven, S.; Sexton, J.; Fevang, B.; Mydel, P.; Haavardsholm, E. Peptidylarginine Deiminase 4 (PAD4) Activity in Early Rheumatoid Arthritis. Scand. J. Rheumatol. 2020, 49, 87–95. [Google Scholar] [CrossRef]
- Krishnamurthy, A.; Joshua, V.; Haj Hensvold, A.; Jin, T.; Sun, M.; Vivar, N.; Ytterberg, A.J.; Engström, M.; Fernandes-Cerqueira, C.; Amara, K.; et al. Identification of a Novel Chemokine-Dependent Molecular Mechanism Underlying Rheumatoid Arthritis-Associated Autoantibody-Mediated Bone Loss. Ann. Rheum. Dis. 2016, 75, 721–729. [Google Scholar] [CrossRef]
- Chang, X. Localization of Peptidylarginine Deiminase 4 (PADI4) and Citrullinated Protein in Synovial Tissue of Rheumatoid Arthritis. Rheumatology 2005, 44, 40–50. [Google Scholar] [CrossRef]
- Asaga, H.; Nakashima, K.; Senshu, T.; Ishigami, A.; Yamada, M. Immunocytochemical Localization of Peptidylarginine Deiminase in Human Eosinophils and Neutrophils. J. Leukoc. Biol. 2001, 70, 46–51. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographics | RA (n = 100) |
| Age (years) a | 47 ± 14 |
| Gender | |
| Female % (n) Male % (n) | 87 (87) 13 (13) |
| Smoking | |
| Smokers % (n) | 39 (39) |
| Non-smokers % (n) | 61 (61) |
| Clinical assessment | |
| Disease evolution (years) b | 5 (1–10) |
| Early RA ≤ 1 year of evolution % (n) | 29.2 (28) |
| Established RA > 1 year of evolution % (n) | 70.8 (68) |
| Age of disease onset a | 41 ± 12 |
| DAS28 clinical activity b | 5.0 (3.5–5.8) |
| HAQ-DI functional disability b | 0.6 (0.2–1) |
| RF (U/ mL) b | 89.3 (28.3–300) |
| Positive RF % (n) | 54 (54) |
| Negative RF %(n) | 46 (46) |
| Anti-CCP antibodies (U/ mL) b | 100.2 (2.7–185.1) |
| Positive anti-CCP % (n) | 72.7 (72) |
| Negative anti-CCP % (n) | 27.8 (27) |
| Anti-PADI4 antibodies (U/ Ml) b | 2026.9 (188.9–3359.6) |
| ESR (mm/h) b | 39 (23–46) |
| hsCRP (mg/L) b | 24.7 (12.3–41) |
| Polymorphism | CS % (n = 100) | RA % (n = 100) | OR (IC 95%) | p |
|---|---|---|---|---|
| PADI4_89 G>A | ||||
| AA a | 34 (34) | 20 (20) | 1 | |
| GA | 52 (52) | 53 (53) | 1.73 (0.84–3.6) | 0.107 |
| GG | 14 (14) | 27 (27) | 3.28 (1.3–8.4) | 0.005 |
| Allele | ||||
| A a | 60 (120) | 46.5 (93) | 1 | |
| G | 40 (80) | 53.5 (107) | 1.72 (1.14–2.62) | 0.007 |
| PADI4_90 T>C | ||||
| CC a | 33 (33) | 19 (19) | 1 | |
| TC | 51 (51) | 55 (55) | 1.51 (0.69–3.37) | 0.270 |
| TT | 16 (16) | 26 (26) | 2.82 (1.12–7.14) | 0.014 |
| Allele | ||||
| C a | 58.5 (117) | 46.5 (93) | 1 | |
| T | 41.5 (83) | 53.5 (107) | 1.62 (1.07–2.46) | 0.016 |
| PADI4_92 G>C | ||||
| CC a | 37 (37) | 21 (21) | 1 | |
| GC | 48 (48) | 54 (54) | 1.98 (0.97–4.07) | 0.041 |
| GG | 15 (15) | 25 (25) | 2.94 (1.18–7.38) | 0.010 |
| Allele | ||||
| C a | 61 (122) | 52 (104) | 1 | |
| G | 39 (78) | 48 (96) | 1.69 (1.11–2.57) | 0.009 |
| Haplotype | CS % (n) | RA % (n) | OR (IC 95%) | P |
|---|---|---|---|---|
| GTG | 32.5 (13) | 57.9 (22) | 2.86 (1.03–7.99) | 0.024 |
| ACC a | 67.5 (27) | 42.1 (16) | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matuz-Flores, M.G.; Rosas-Rodríguez, J.A.; Tortoledo-Ortiz, O.; Muñoz-Barrios, S.; Martínez-Bonilla, G.E.; Hernández-Bello, J.; Baños-Hernández, C.J.; Pacheco-Tena, C.; Sánchez-Zuno, G.A.; Panduro-Espinoza, B.; et al. PADI4 Haplotypes Contribute to mRNA Expression, the Enzymatic Activity of Peptidyl Arginine Deaminase and Rheumatoid Arthritis Risk in Patients from Western Mexico. Curr. Issues Mol. Biol. 2022, 44, 4268-4281. https://doi.org/10.3390/cimb44090293
Matuz-Flores MG, Rosas-Rodríguez JA, Tortoledo-Ortiz O, Muñoz-Barrios S, Martínez-Bonilla GE, Hernández-Bello J, Baños-Hernández CJ, Pacheco-Tena C, Sánchez-Zuno GA, Panduro-Espinoza B, et al. PADI4 Haplotypes Contribute to mRNA Expression, the Enzymatic Activity of Peptidyl Arginine Deaminase and Rheumatoid Arthritis Risk in Patients from Western Mexico. Current Issues in Molecular Biology. 2022; 44(9):4268-4281. https://doi.org/10.3390/cimb44090293
Chicago/Turabian StyleMatuz-Flores, Mónica Guadalupe, Jesús Alfredo Rosas-Rodríguez, Orlando Tortoledo-Ortiz, Salvador Muñoz-Barrios, Gloria Esther Martínez-Bonilla, Jorge Hernández-Bello, Christian Johana Baños-Hernández, Cesar Pacheco-Tena, Gabriela Athziri Sánchez-Zuno, Beatriz Panduro-Espinoza, and et al. 2022. "PADI4 Haplotypes Contribute to mRNA Expression, the Enzymatic Activity of Peptidyl Arginine Deaminase and Rheumatoid Arthritis Risk in Patients from Western Mexico" Current Issues in Molecular Biology 44, no. 9: 4268-4281. https://doi.org/10.3390/cimb44090293
APA StyleMatuz-Flores, M. G., Rosas-Rodríguez, J. A., Tortoledo-Ortiz, O., Muñoz-Barrios, S., Martínez-Bonilla, G. E., Hernández-Bello, J., Baños-Hernández, C. J., Pacheco-Tena, C., Sánchez-Zuno, G. A., Panduro-Espinoza, B., & Muñoz-Valle, J. F. (2022). PADI4 Haplotypes Contribute to mRNA Expression, the Enzymatic Activity of Peptidyl Arginine Deaminase and Rheumatoid Arthritis Risk in Patients from Western Mexico. Current Issues in Molecular Biology, 44(9), 4268-4281. https://doi.org/10.3390/cimb44090293