
Investigation of the Molecular Evolution of Treg Suppression Mechanisms Indicates a Convergent Origin

 , , ,
, , ,  ,
,
Abstract
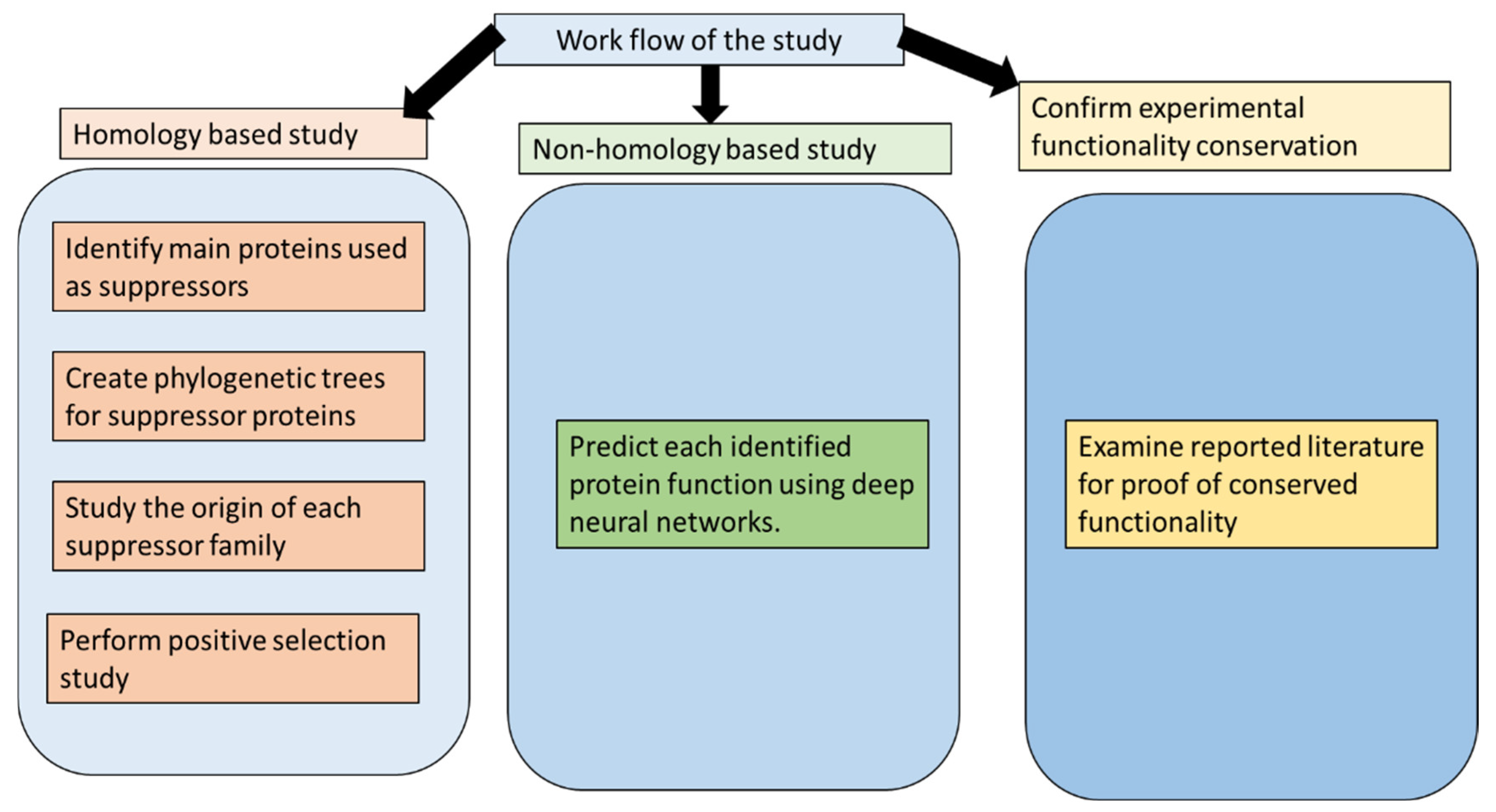
:1. Introduction
2. Methods
2.1. Database Search
2.2. Alignment and Phylogenetic Analysis
2.3. Ancestral Sequence Reconstruction (ASR)
2.4. HHsearch
2.5. Non-Homology Functional Prediction
2.6. Positive Selection
3. Results
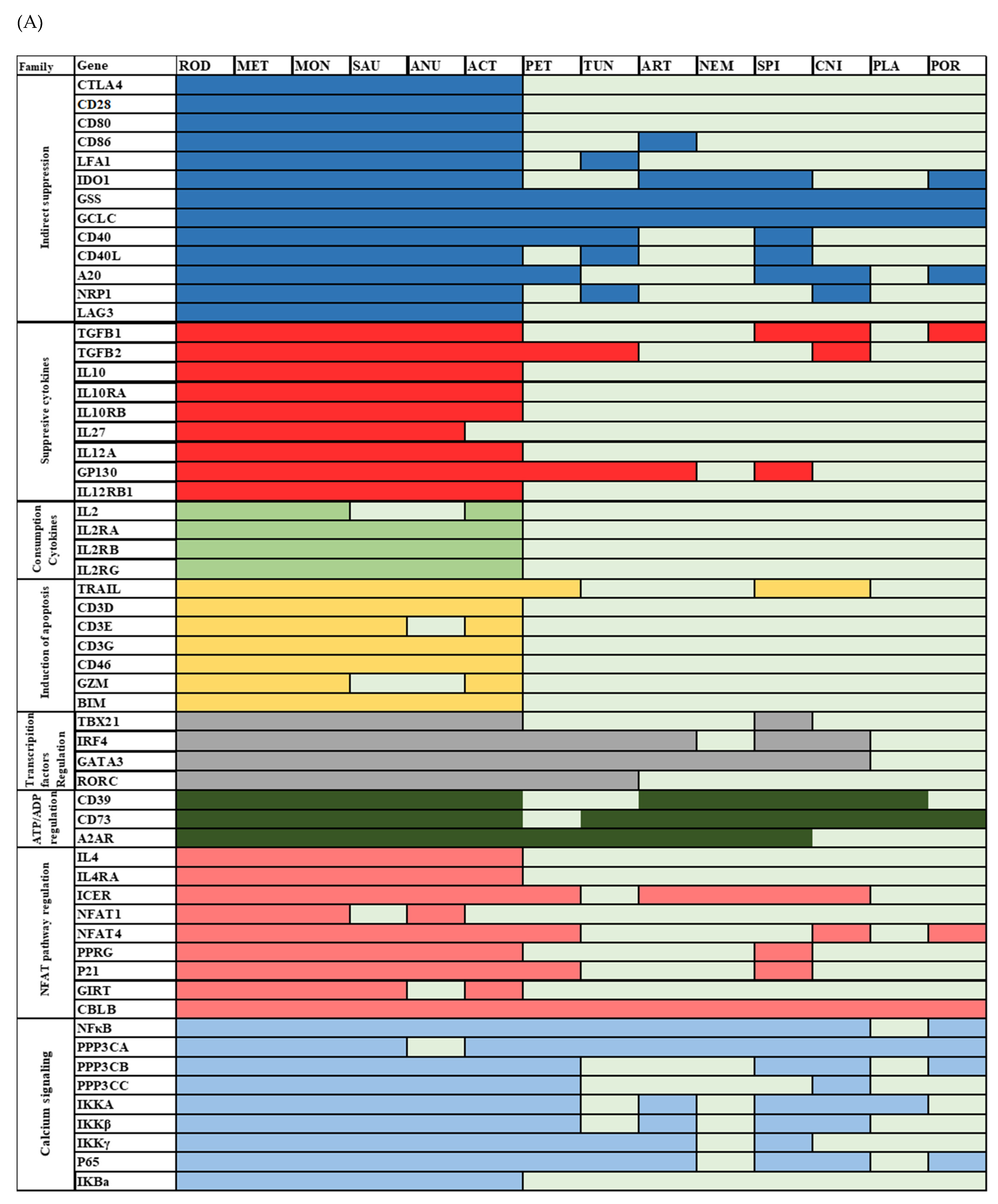
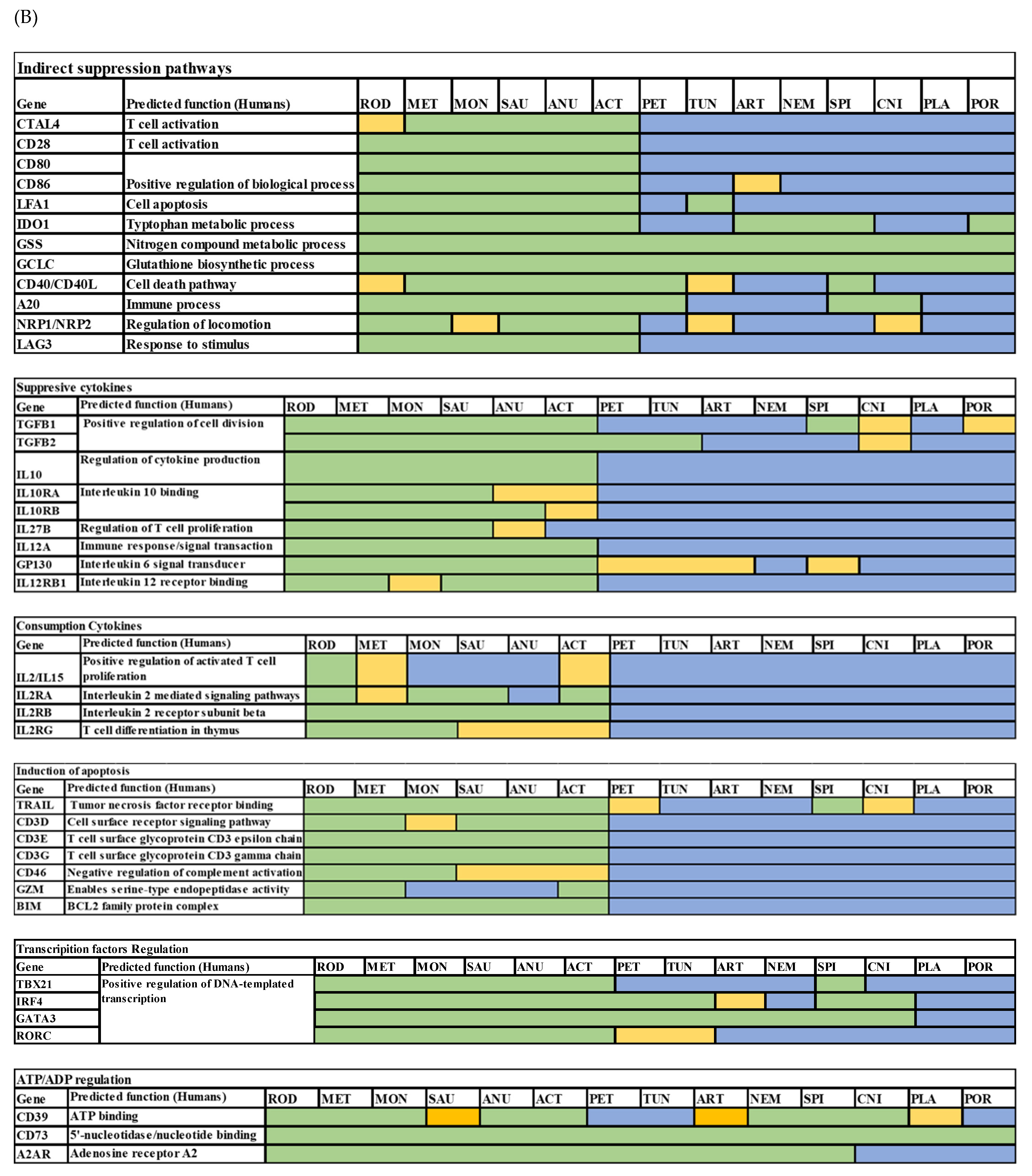
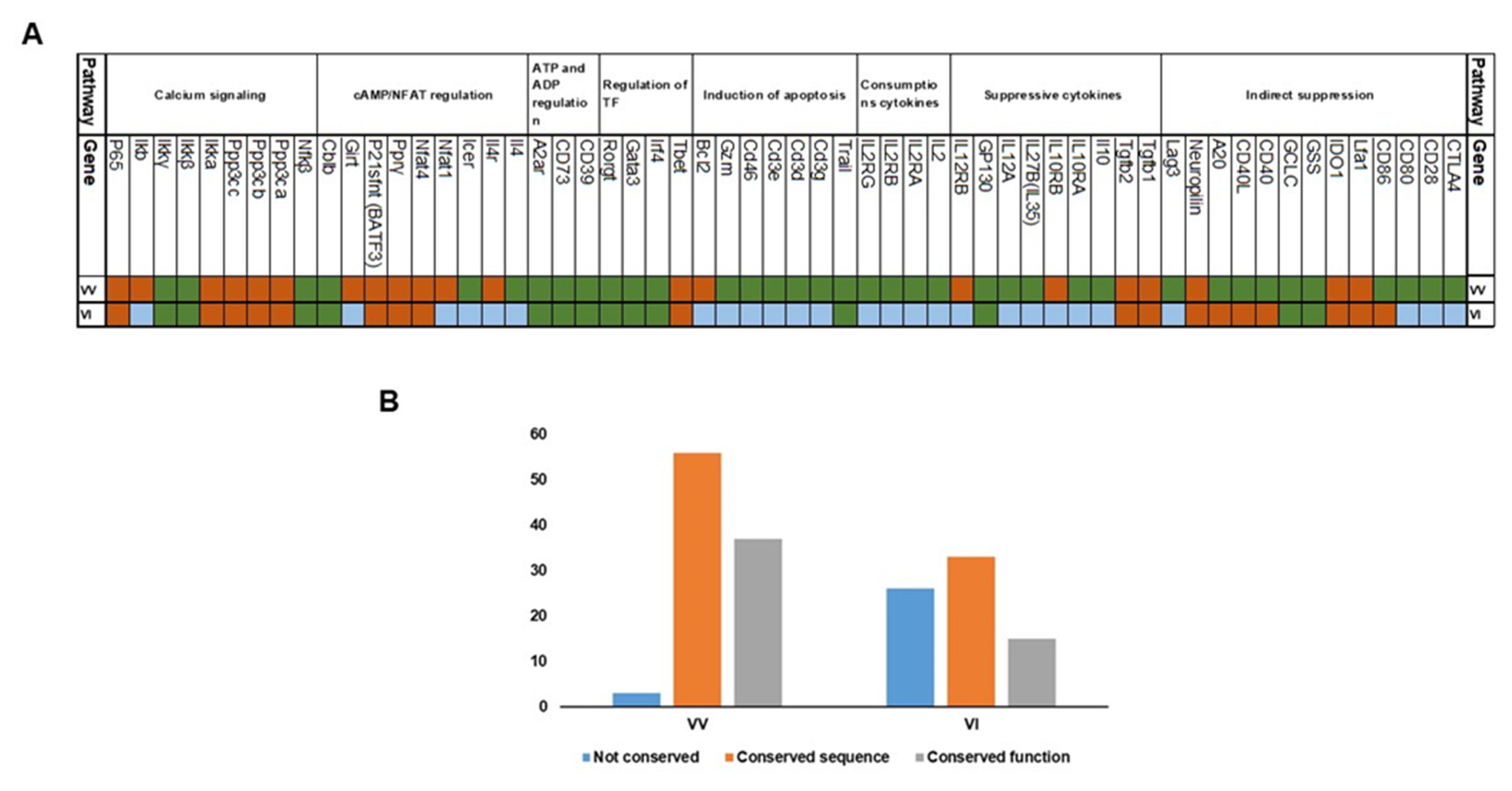
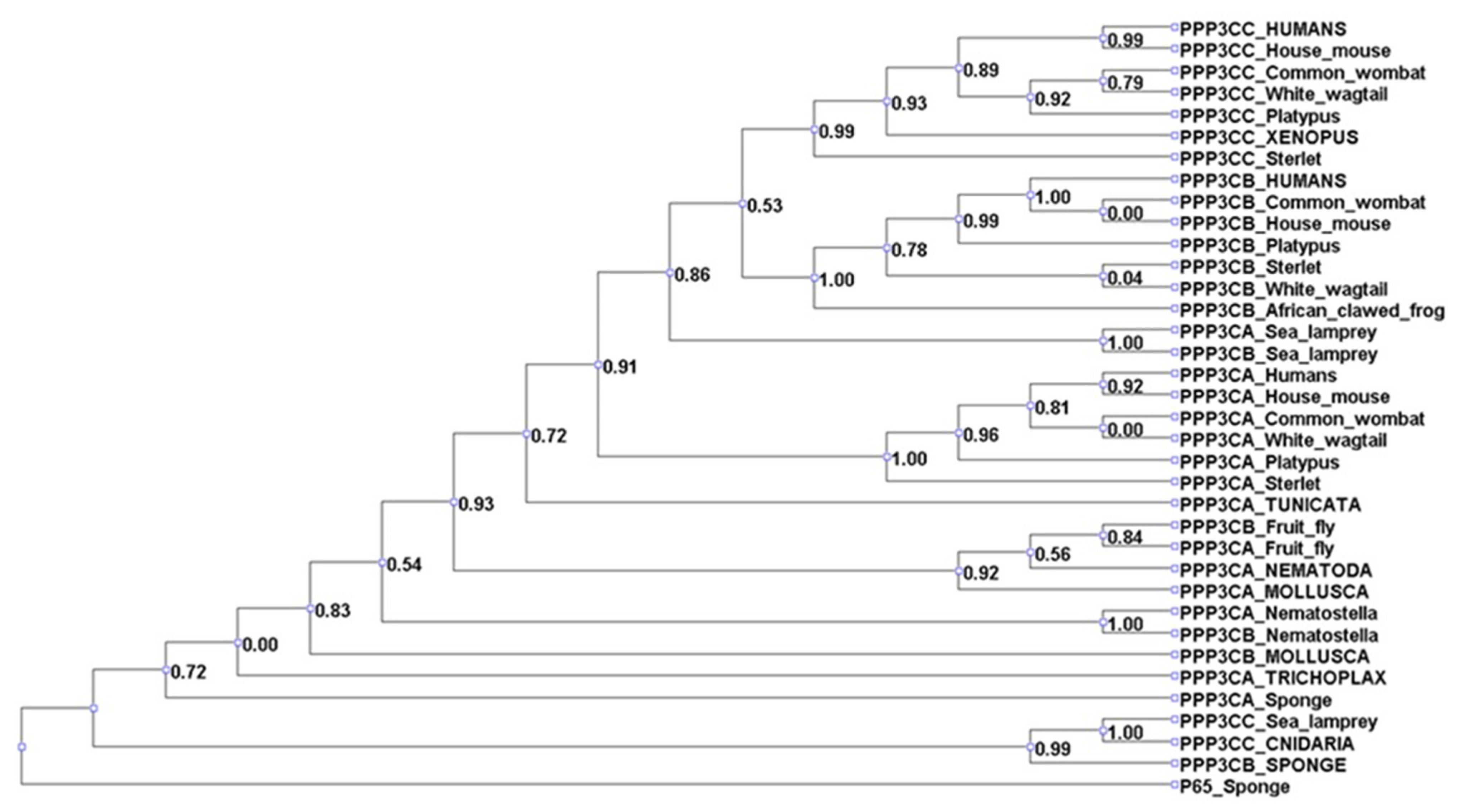
3.1. Analysis of Treg Suppressor Markers Reveals Multiple Origins of Suppression Mechanisms
3.2. Positive Selection Analysis Indicates That Tregs Employed Both Highly Conserved and Rapidly Evolving Mechanisms
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostroukhova, M.; Qi, Z.; Oriss, T.B.; Dixon-McCarthy, B.; Ray, P.; Ray, A. Treg-Mediated Immunosuppression Involves Activation of the Notch-HES1 Axis by Membrane-Bound TGF-β. J. Clin. Investig. 2006, 116, 996–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Kitani, A.; Strober, W. Cell Contact-Dependent Immunosuppression by CD4+CD25+ Regulatory T Cells Is Mediated by Cell Surface-Bound Transforming Growth Factor β. J. Exp. Med. 2001, 194, 629–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kühn, R.; Löhler, J.; Rennick, D.; Rajewsky, K.; Müller, W. Interleukin-10-Deficient Mice Develop Chronic Enterocolitis. Cell 1993, 75, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.D.; Di Marco, F.; Hooley, J.; Pitts-Meek, S.; Bauer, M.; Ryan, A.M.; Sordat, B.; Gibbs, V.C.; Aguet, M. The Orphan Receptor CRF2-4 Is an Essential Subunit of the Interleukin 10 Receptor. J. Exp. Med. 1998, 187, 571–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, B.M.; Sullivan, J.A.; Burlingham, W.J. Interleukin 35: A Key Mediator of Suppression and the Propagation of Infectious Tolerance. Front. Immunol. 2013, 4, 315. [Google Scholar] [CrossRef] [Green Version]
- Thornton, A.M.; Shevach, E.M. CD4+CD25+ Immunoregulatory T Cells Suppress Polyclonal T Cell Activation in Vitro by Inhibiting Interleukin 2 Production. J. Exp. Med. 1998, 188, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Bodor, J.; Bodorova, J.; Gress, R.E. Suppression of T Cell Function: A Potential Role for Transcriptional Repressor ICER. J. Leukoc. Biol. 2000, 67, 774–779. [Google Scholar] [CrossRef]
- Bhaumik, S.; Mickael, M.E.; Moran, M.; Spell, M.; Basu, R. RORγt Promotes Foxp3 Expression by Antagonizing the Effector Program in Colonic Regulatory T Cells. J. Immunol. 2021, 207, 2027–2038. [Google Scholar] [CrossRef]
- Read, S.; Malmström, V.; Powrie, F. Cytotoxic T Lymphocyte-Associated Antigen 4 Plays an Essential Role in the Function of CD25+CD4+ Regulatory Cells That Control Intestinal Inflammation. J. Exp. Med. 2000, 192, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Sarris, M.; Andersen, K.G.; Randow, F.; Mayr, L.; Betz, A.G. Neuropilin-1 Expression on Regulatory T Cells Enhances Their Interactions with Dendritic Cells during Antigen Recognition. Immunity 2008, 28, 402–413. [Google Scholar] [CrossRef]
- Liang, B.; Workman, C.; Lee, J.; Chew, C.; Dale, B.M.; Colonna, L.; Flores, M.; Li, N.; Schweighoffer, E.; Greenberg, S.; et al. Regulatory T Cells Inhibit Dendritic Cells by Lymphocyte Activation Gene-3 Engagement of MHC Class II. J. Immunol. 2008, 180, 5916–5926. [Google Scholar] [CrossRef] [Green Version]
- Song, X.T.; Kabler, K.E.; Shen, L.; Rollins, L.; Huang, X.F.; Chen, S.Y. A20 Is an Antigen Presentation Attenuator, and Its Inhibition Overcomes Regulatory T Cell-Mediated Suppression. Nat. Med. 2008, 14, 258–265. [Google Scholar] [CrossRef]
- Hänig, J.; Lutz, M.B. Suppression of Mature Dendritic Cell Function by Regulatory T Cells In Vivo Is Abrogated by CD40 Licensing. J. Immunol. 2008, 180, 1405–1413. [Google Scholar] [CrossRef] [Green Version]
- Litman, G.W.; Rast, J.P.; Fugmann, S.D. The Origins of Vertebrate Adaptive Immunity. Nat. Rev. Immunol. 2010, 10, 543–553. [Google Scholar] [CrossRef] [Green Version]
- Boehm, T.; McCurley, N.; Sutoh, Y.; Schorpp, M.; Kasahara, M.; Cooper, M.D. VLR-Based Adaptive Immunity. Annu. Rev. Immunol. 2012, 30, 203–220. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, T.; Shibasaki, Y.; Matsuura, Y. T Cells in Fish. Biology 2015, 4, 640–663. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Hirano, M.; Herrin, B.R.; Li, J.; Yu, C.; Sadlonova, A.; Cooper, M.D. Dual Nature of the Adaptive Immune System in Lampreys. Nature 2009, 459, 796–801. [Google Scholar] [CrossRef] [Green Version]
- Bhaumik, S.; Basu, R. Cellular and Molecular Dynamics of Th17 Differentiation and Its Developmental Plasticity in the Intestinal Immune Response. Front. Immunol. 2017, 8, 254. [Google Scholar] [CrossRef] [Green Version]
- Mickael, M.E.; Bhaumik, S.; Basu, R. Retinoid-Related Orphan Receptor RORγt in CD4+ T-Cell–Mediated Intestinal Homeostasis and Inflammation. Am. J. Pathol. 2020, 190, 1984–1999. [Google Scholar] [CrossRef]
- Lee, Y.K.; Mukasa, R.; Hatton, R.D.; Weaver, C.T. Developmental Plasticity of Th17 and Treg Cells. Curr. Opin. Immunol. 2009, 21, 274–280. [Google Scholar] [CrossRef]
- Schmidt, A.; Oberle, N.; Krammer, P.H. Molecular Mechanisms of Treg-Mediated T Cell Suppression. Front. Immunol. 2012, 3, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulmanov, M.; Khan, M.A.; Hoehndorf, R. DeepGO: Predicting Protein Functions from Sequence and Interactions Using a Deep Ontology-Aware Classifier. Bioinformatics 2018, 34, 660–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubick, N.; Brösamle, D.; Mickael, M.E. Molecular Evolution and Functional Divergence of the IgLON Family. Evol. Bioinform. 2018, 14, 1176934318775081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubick, N.; Klimovich, P.; Flournoy, P.H.; Bieńkowska, I.; Łazarczyk, M.; Sacharczuk, M.; Bhaumik, S.; Mickael, M.-E.; Basu, R. Interleukins and Interleukin Receptors Evolutionary History and Origin in Relation to CD4+ T Cell Evolution. Genes 2021, 12, 813. [Google Scholar] [CrossRef] [PubMed]
- Mickael, M.E.; Rajput, A.; Steyn, J.; Wiemerslage, L.; Bürglin, T. An Optimised Phylogenetic Method Sheds More Light on the Main Branching Events of Rhodopsin-like Superfamily. Comp. Biochem. Physiol. Part D Genom. Proteom. 2016, 20, 85–94. [Google Scholar] [CrossRef]
- Mickael, M.-E.; Kubick, N.; Klimovich, P.; Flournoy, P.H.; Bieńkowska, I.; Sacharczuk, M. Paracellular and Transcellular Leukocytes Diapedesis Are Divergent but Interconnected Evolutionary Events. Genes 2021, 12, 254. [Google Scholar] [CrossRef]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Gabler, F.; Nam, S.Z.; Till, S.; Mirdita, M.; Steinegger, M.; Söding, J.; Lupas, A.N.; Alva, V. Protein Sequence Analysis Using the MPI Bioinformatics Toolkit. Curr. Protoc. Bioinform. 2020, 72, e108. [Google Scholar] [CrossRef]
- Törönen, P.; Medlar, A.; Holm, L. PANNZER2: A Rapid Functional Annotation Web Server. Nucleic Acids Res. 2018, 46, W84–W88. [Google Scholar] [CrossRef] [Green Version]
- Bernhofer, M.; Dallago, C.; Karl, T.; Satagopam, V.; Heinzinger, M.; Littmann, M.; Olenyi, T.; Qiu, J.; Schütze, K.; Yachdav, G.; et al. PredictProtein—Predicting Protein Structure and Function for 29 Years. Nucleic Acids Res. 2021, 49, W535–W540. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Jia, S.; Shao, X.; Liu, S.; Zhang, Q.; Song, J.; Wang, W.; Jin, Z. Interleukin-35 Modulates the Balance between Viral Specific CD4 + CD25 + CD127 Dim/- Regulatory T Cells and T Helper 17 Cells in Chronic Hepatitis B Virus Infection. Virol. J. 2019, 16, 48. [Google Scholar] [CrossRef]
- Staudt, V.; Bothur, E.; Klein, M.; Lingnau, K.; Reuter, S.; Grebe, N.; Gerlitzki, B.; Hoffmann, M.; Ulges, A.; Taube, C.; et al. Interferon-Regulatory Factor 4 Is Essential for the Developmental Program of T Helper 9 Cells. Immunity 2010, 33, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Patient, R.K.; McGhee, J.D. The GATA Family (Vertebrates and Invertebrates). Curr. Opin. Genet. Dev. 2002, 12, 416–422. [Google Scholar] [CrossRef]
- Chadwick, B.P.; Frischauf, A.-M. The CD39-like Gene Family: Identification of Three New Human Members (CD39L2, CD39L3, and CD39L4), Their Murine Homologues, and a Member of the Gene Family FromDrosophila Melanogaster. Genomics 1998, 50, 357–367. [Google Scholar] [CrossRef]
- Fenckova, M.; Hobizalova, R.; Fric, Z.F.; Dolezal, T. Functional Characterization of Ecto-5′-Nucleotidases and Apyrases in Drosophila Melanogaster. Insect Biochem. Mol. Biol. 2011, 41, 956–967. [Google Scholar] [CrossRef]
- Kucerova, L.; Broz, V.; Fleischmannova, J.; Santruckova, E.; Sidorov, R.; Dolezal, V.; Zurovec, M. Characterization of the Drosophila Adenosine Receptor: The Effect of Adenosine Analogs on CAMP Signaling in Drosophila Cells and Their Utility for in Vivo Experiments. J. Neurochem. 2012, 121, 383–395. [Google Scholar] [CrossRef]
- Bodor, J.; Fehervari, Z.; Diamond, B.; Sakaguchi, S. ICER/CREM-Mediated Transcriptional Attenuation of IL-2 and Its Role in Suppression by Regulatory T Cells. Eur. J. Immunol. 2007, 37, 884–895. [Google Scholar] [CrossRef]
- Van Der Merwe, P.A.; Bodian, D.L.; Daenke, S.; Linsley, P.; Davis, S.J. CD80 (B7-1) Binds Both CD28 and CTLA-4 with a Low Affinity and Very Fast Kinetics. J. Exp. Med. 1997, 185, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Oderup, C.; Cederbom, L.; Makowska, A.; Cilio, C.M.; Ivars, F. Cytotoxic T Lymphocyte Antigen-4-Dependent down-Modulation of Costimulatory Molecules on Dendritic Cells in CD4+ CD25+ Regulatory T-Cell-Mediated Suppression. Immunology 2006, 118, 240–249. [Google Scholar] [CrossRef]
- Lefort, C.T.; Ley, K. Neutrophil Arrest by LFA-1 Activation. Front. Immunol. 2012, 3, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, M.W.; Guo, H.F.; Li, X.; Linkugel, A.D.; Vander Kooi, C.W. Function of Members of the Neuropilin Family as Essential Pleiotropic Cell Surface Receptors. Biochemistry 2012, 51, 9437–9446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bots, M.; Medema, J.P. Granzymes at a Glance. J. Cell Sci. 2006, 119, 5011–5014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, A.; Luo, C.; Hogan, P.G. Transcription Factors of the NFAT Family: Regulation and Function. Annu. Rev. Immunol. 1997, 15, 707–747. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H. 5′-Nucleotidase: Molecular Structure and Functional Aspects. Biochem. J. 1992, 285, 345–365. [Google Scholar] [CrossRef] [Green Version]
- Robson, S.C.; Sévigny, J.; Zimmermann, H. The E-NTPDase Family of Ectonucleotidases: Structure Function Relationships and Pathophysiological Significance. Purinergic Signal. 2006, 2, 409–430. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Yan, X.; Wang, G.; Liu, H.; Gan, X.; Zhang, T.; Wang, J.; Li, L. Evolutionary Pattern and Regulation Analysis to Support Why Diversity Functions Existed within PPAR Gene Family Members. BioMed Res. Int. 2015, 2015, 613910. [Google Scholar] [CrossRef] [Green Version]
- Bernard, D.; Riteau, B.; Hansen, J.D.; Phillips, R.B.; Michel, F.; Boudinot, P.; Benmansour, A. Costimulatory Receptors in a Teleost Fish: Typical CD28, Elusive CTLA4. J. Immunol. 2006, 176, 4191–4200. [Google Scholar] [CrossRef] [Green Version]
- Muñiz Ortiz, J.G.; Opoka, R.; Kane, D.; Cartwright, I.L. Investigating Arsenic Susceptibility from a Genetic Perspective in Drosophila Reveals a Key Role for Glutathione Synthetase. Toxicol. Sci. 2009, 107, 416–426. [Google Scholar] [CrossRef] [Green Version]
- Mendoza-Coronel, E.; Castañón-Arreola, M. Comparative Evaluation of in Vitro Human Macrophage Models for Mycobacterial Infection Study. Pathog. Dis. 2016, 74, ftw052. [Google Scholar] [CrossRef]
- Hui, S.P.; Sheng, D.Z.; Sugimoto, K.; Gonzalez-Rajal, A.; Nakagawa, S.; Hesselson, D.; Kikuchi, K. Zebrafish Regulatory T Cells Mediate Organ-Specific Regenerative Programs. Dev. Cell 2017, 43, 659–672.e5. [Google Scholar] [CrossRef] [Green Version]
- Burkhardt, N.B.; Elleder, D.; Schusser, B.; Krchlíková, V.; Göbel, T.W.; Härtle, S.; Kaspers, B. The Discovery of Chicken Foxp3 Demands Redefinition of Avian Regulatory T Cells. J. Immunol. 2022, 208, 1128–1138. [Google Scholar] [CrossRef]
- Dijkstra, J.M. TH2 and Treg Candidate Genes in Elephant Shark. Nature 2014, 511, E7–E9. [Google Scholar] [CrossRef]
- Venkatesh, B.; Lee, A.P.; Ravi, V.; Maurya, A.K.; Lian, M.M.; Swann, J.B.; Ohta, Y.; Flajnik, M.F.; Sutoh, Y.; Kasahara, M.; et al. Elephant Shark Genome Provides Unique Insights into Gnathostome Evolution. Nature 2014, 505, 174–179. [Google Scholar] [CrossRef] [Green Version]
- Chung-Davidson, Y.W.; Ren, J.; Yeh, C.Y.; Bussy, U.; Huerta, B.; Davidson, P.J.; Whyard, S.; Li, W. TGF-β Signaling Plays a Pivotal Role During Developmental Biliary Atresia in Sea Lamprey (Petromyzon Marinus). Hepatol. Commun. 2020, 4, 219–234. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Guo, J.; Cheng, X.; Li, W.; Lyu, S.; Chen, X.; Li, Q.; Wang, H. Molecular Evolution of Transforming Growth Factor-β (TGF-β) Gene Family and the Functional Characterization of Lamprey TGF-Β2. Front. Immunol. 2022, 13, 2318. [Google Scholar] [CrossRef]
- Detournay, O.; Schnitzler, C.E.; Poole, A.; Weis, V.M. Regulation of Cnidarian–Dinoflagellate Mutualisms: Evidence That Activation of a Host TGFβ Innate Immune Pathway Promotes Tolerance of the Symbiont. Dev. Comp. Immunol. 2012, 38, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Liongue, C.; Ward, A.C. Evolution of Class I Cytokine Receptors. BMC Evol. Biol. 2007, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Yang, C.; Liu, J.; Huang, T.; Lin, L.; Gu, L.; Li, Z.; Chen, M. Functional Characterization of a Putative Tumor Necrosis Factor Superfamily Member 10 in Blood Clam (Tegillarca Granosa). Dev. Comp. Immunol. 2021, 124, 104172. [Google Scholar] [CrossRef]
- Das, S.; Li, J.; Hirano, M.; Sutoh, Y.; Herrin, B.R.; Cooper, M.D. Evolution of Two Prototypic T Cell Lineages. Cell. Immunol. 2015, 296, 87–94. [Google Scholar] [CrossRef]
- Chen, Y.C.; Liu, T.; Yu, C.H.; Chiang, T.Y.; Hwang, C.C. Effects of GC Bias in Next-Generation-Sequencing Data on De Novo Genome Assembly. PLoS ONE 2013, 8, e62856. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.J.; Kuraku, S.; Holt, C.; Sauka-Spengler, T.; Jiang, N.; Campbell, M.S.; Yandell, M.D.; Manousaki, T.; Meyer, A.; Bloom, O.E.; et al. Sequencing of the Sea Lamprey (Petromyzon Marinus) Genome Provides Insights into Vertebrate Evolution. Nat. Genet. 2013, 45, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. Evolutionary Rate at the Molecular Level. Nature 1968, 217, 624–626. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Genes | |
|---|---|---|
| Indirect pathway | Inhibit DC | CTLA4, CD28, CD80, CD86, LFAI-1, A20, CD40-CD40L, neuropilin-1 and LAG3 |
| Direct pathways | Produce Suppressive cytokines | TGFβ, IL10 and IL35 |
| Consumption of cytokines | IL2, IL2Rα, IL2Rβ and IL2Rγ | |
| Induction of apoptosis | TRAIL, CD3, CD46, CD25 and BIM | |
| Regulation of TF | IRF4, GATA3, FoxP3 and RORγt | |
| ATP and ADP regulation | CD73, CD39, APRT, A2A receptors, and P2RY11 | |
| cAMP/NFAT regulation | IL4, ICER, PPRγ, p21SNFT, GITR and CBLB | |
| Calcium signaling | NFκB, PPP3CA, PPP3CB, PPP3CC, IKKα, IKKβ, IKKγ, IκB IKBA, and p65 |
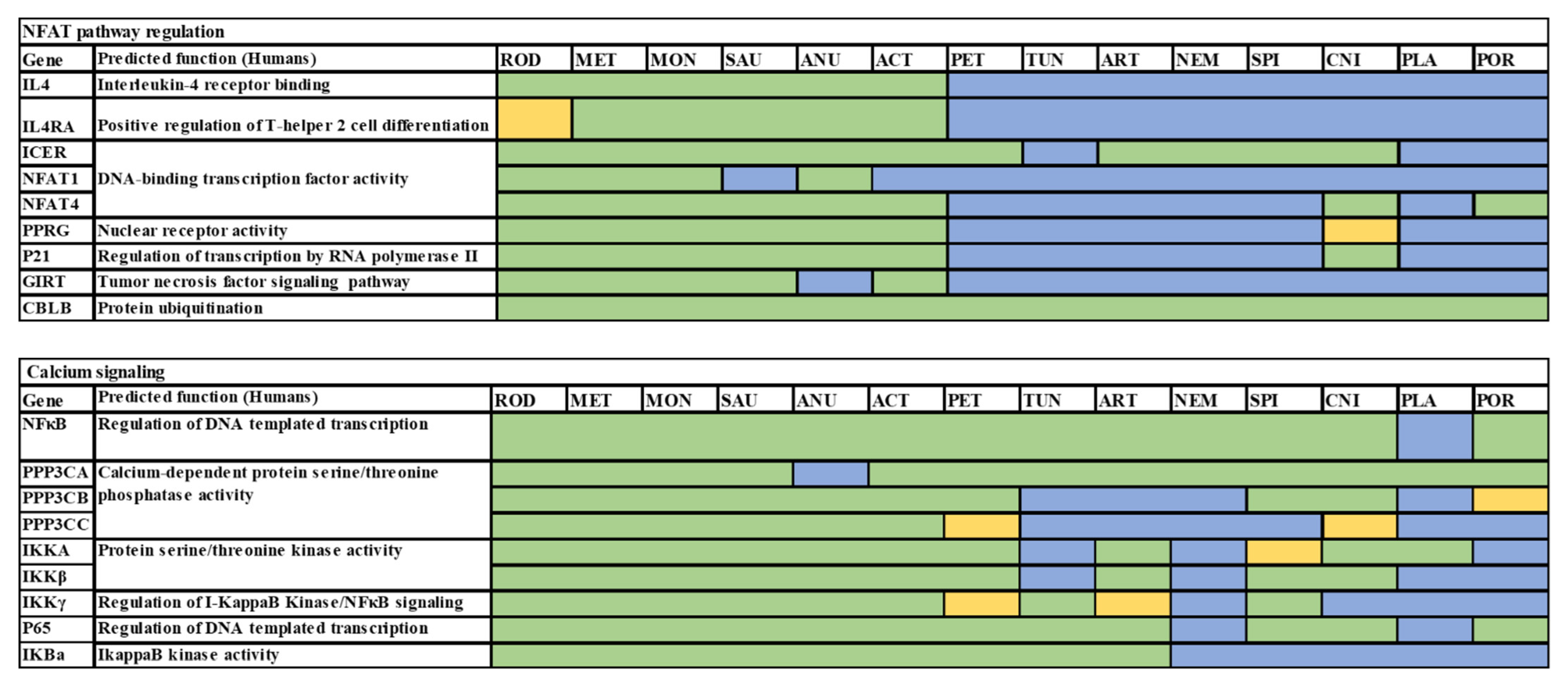
| Gene | Conserved Function between Vertebrates and Invertebrates | Conserved Function within Vertebrates | |
|---|---|---|---|
| 1 | CTLA4 | No evidence for conservation of function | PMID: 16547256 |
| 2 | CD28 | No evidence for conservation of function | PMID: 16547256 |
| 3 | CD80 | No evidence for conservation of function | PMID: 30107249 PMID: 19535623 |
| 4 | CD86 | No evidence for conservation of function | PMID: 30107249 PMID: 19535623 |
| 5 | Lfa1 | No evidence for conservation of function | |
| 6 | IDO1 | No evidence for conservation of function | |
| 7 | GSS | Conserved function in Drosophila PMID: 18779381 | PMID: 21802407 |
| 8 | GCLC | Conserved function in Drosophila PMID: 19036725 | PMID: 28158580 |
| 9 | CD40 | No evidence for conservation of function | PMID: 35667467 |
| 10 | CD40L | No evidence for conservation of function | PMID: 35667467 |
| 11 | A20 | No evidence for conservation of function | PMID: 33154446 |
| 12 | Neuropilin | No evidence for conservation of function | |
| 13 | Lag3 | No evidence for conservation of function | PMID: 16951357 |
| 14 | Tgfb1 | No evidence for conservation of function | |
| 15 | Tgfb2 | No evidence for conservation of function | |
| 16 | Il10 | No evidence for conservation of function | PMID: 25416810 |
| 17 | IL10RA | No evidence for conservation of function | PMID: 25416810 |
| 18 | IL10RB | No evidence for conservation of function | No evidence for conservation of function |
| 19 | IL27B(IL35) | No evidence for conservation of function | PMID: 30590066 |
| 20 | IL12A | No evidence for conservation of function | PMID: 30204772 (similar) |
| 21 | GP130 | Conserved in Drosophila PMID: 14504285 | PMID: 18687405 |
| 22 | IL12RB | No evidence for conservation of function | No evidence for conservation of function |
| 23 | IL2 | No evidence for conservation of function | PMID: 30093902 |
| 24 | IL2RA | No evidence for conservation of function | PMID: 30093902 |
| 25 | IL2RB | No evidence for conservation of function | PMID: 35218892 |
| 26 | IL2RG | No evidence for conservation of function | PMID: 35218892 |
| 27 | Trail | Seemed to be homologs with Drosophila gene Eiger, PMID: 12065414. Evidence for conserved function of inducing apoptosis. | PMID: 33483333 |
| 28 | Cd3g | No evidence for conservation of function | PMID: 19744563 |
| 29 | Cd3d | No evidence for conservation of function | PMID: 19744563 |
| 30 | Cd3e | No evidence for conservation of function | PMID: 19744563 |
| 31 | Cd46 | No evidence for conservation of function | PMID: 25452563 |
| 32 | Gzm | No evidence for conservation of function | PMID: 31736981 |
| 33 | Bim (Bcl2l11) | No evidence for conservation of function | No evidence for conservation of function |
| 34 | Tbet | Some function conserved in ectoderms | PMID: 25016582 |
| 35 | Irf4 | Conserved function not proven | PMID: 19638535 |
| 36 | Gata3 | Conserved function not proven | PMID: 19638535 PMID: 12100886 |
| 37 | Rorgt | Conserved function not proven | PMID: 16990136 |
| 38 | Cd39 | Conserved function in Drosophila and even in plants | PMID: 9676430 |
| 39 | Cd73 | Conserved function in Drosophila | PMID: 21996016 |
| 40 | A2ar | Seems to have a conserved in Drosophila | PMID: 32108870 |
| 41 | P2ry11 | No evidence for conservation of function | |
| 42 | Il4 | No evidence for conservation of function | PMID: 32641385 |
| 43 | Il4r | No evidence for conservation of function | |
| 44 | Icer | Well conserved in invertebrates on a functional level | PMID: 19434522 |
| 45 | Nfat1 | No evidence for conservation of function | |
| 46 | Nfat4 | No evidence of conserved function | |
| 47 | Pprγ | No evidence of conserved function | |
| 48 | P2 1sfnt (BATF3) | No evidence of conserved function | |
| 49 | Girt | No evidence for conservation of function | |
| 50 | Cblb | Seems Conserved function in both C.elegans and drosophila PMID: 10542134 | PMID 16227975 |
| 51 | Nfkβ | Conserved function in Drosophila | PMID: 20457557 |
| 52 | Ppp3ca | No evidence of conserved function | |
| 53 | Ppp3cb | No evidence of conserved function | |
| 54 | Ppp3cc | No evidence of conserved function | |
| 55 | Ikka | No evidence of conserved function | |
| 56 | Ikkβ | Conserved function | PMID: 34452932 |
| 57 | Ikkγ | Conserved function | PMID: 11017107 |
| 58 | Ikb | No evidence for conservation of function | No evidence of conserved function |
| 59 | P65 | No evidence of conserved function | |
| Molecule | Main Family | Putative Origin | Blastp E-Value | HHsearch Probability (%) |
|---|---|---|---|---|
| CTLA4 | Immunoglobulin superfamily | IGv domain-containing protein (Callorhinchus milii) | 1 × 10−17 | 99.5 |
| LAG3 | IG-like proetin (Nothobranchius kuhntae) | NA | 99 | |
| CD40L | Tumor necrosis factor (TNF) superfamily | Tumor necrosis factor ligand superfamily member 10-like (Lingula unguis) | 3 × 10−16 | 97.9 |
| TRAIL | ||||
| GITR CD40R | Tumor Necrosis Factor Receptor Superfamily | TNFRSF21-like (Monosiga brevicollis) (Choanoflagellata) | 1 × 10−09 | NA |
| A20 (TNFAIP3) | Tumor Necrosis Factor, α Induced Protein family | Tumor necrosis factor α-induced protein 3like (Amphimedon queenslandica) | 1 × 10−40 | 99.8 |
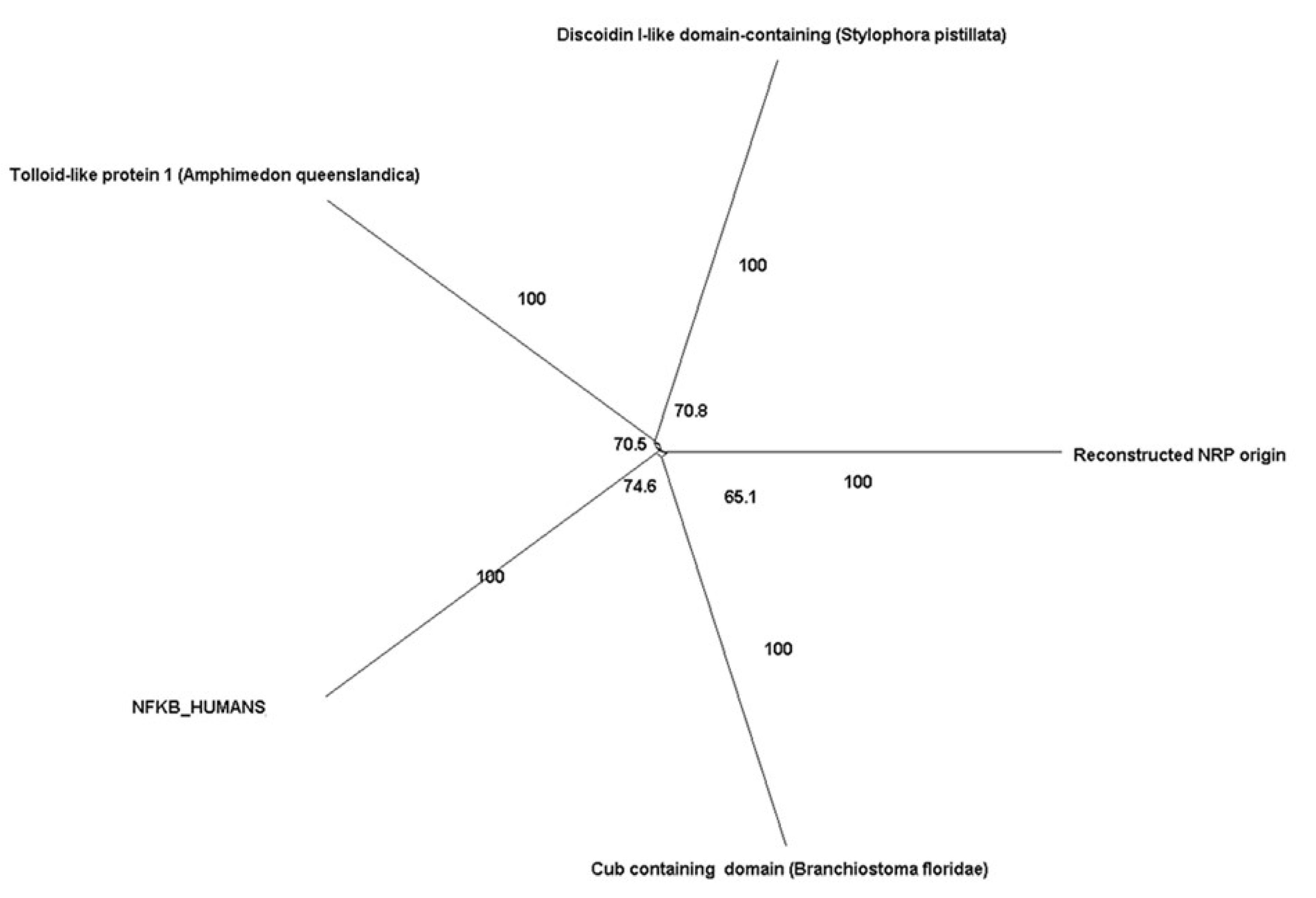
| Neuropilin-1 | Neuropilin family | Tolloid-like protein 1 (Amphimedon queenslandica) | 6 × 10−20 | NA |
| Cub-containing domain (Branchiostoma floridae) | NA | 100 | ||
| Discoidin I-like domain-containing (Stylophora pistillata) | NA | 99 | ||
| GZM B (CTLA1) | Granzymes | Trypsin (Asbolus verrucosus) | 1 × 10−38 | 100 |
| Melanization protease 1 | 2 × 10−33 | NA | ||
| Corin, isoform C (Drosophila melanogaster) | 3 × 10−32 | NA | ||
| CD73 (NT5E) | Ecto-5’-nucleotidase | 5’-nucleotidase/apyrase family protein (Limimaricola hongkongensis) | 4 × 10−124 | 100 |
| CD39 | E-NTPDase family of ectonucleotidases | GDA1/CD39 nucleoside phosphatase (Helicosporidium sp. ATCC 50920) | 7 × 10−61 | NA |
| BATF (p21SNFT) | Basic leucine zipper transcriptional factor ATF-like | jun dimerization protein 2-like (Actinia tenebrosa) | 4 × 10−06 | NA |
| ICER | Unknown | bZIP transcription factor domain containing protein (Acanthamoeba castellanii) | 7 × 10−10 | 100 |
| NFAT | NFATs | NFATc4 (Amphimedon queenslandica) | 5 × 10−23 | NA |
| Cbl-b | ubiquitin ligase | E3 ubiquitin ligase Cbl TKB (Salpingoeca rosetta) (Choanoflagellates) | 2 × 10−101 | NA |
| PPRγ | 5 hydroxyicosatetraenoic acid and 5-oxo- eicosatetraenoic acid family | retinoic acid receptor RXR-alpha-B isoform X1 (Nematostella vectensis) | 9 × 10−40 | NA |
| steroid hormone receptor ERR2-like isoform X2 (Acropora digitifera) | 1 × 10−34 | NA | ||
| Axonema Dynein heavy chain 6 (Exaiptasia diaphana) | 1 × 10−34 | NA |
| Molecule | Global (ω) Value | p-Value |
|---|---|---|
| CTLA4 | 0.42 | <0.0005 |
| CD40 | 0.27 | <0.00001 |
| CD40L | 0.39 | <0.00002 |
| A20 (TNFAIP3) | 0.96 | not significant at p < 0.05. |
| Neuropilin-1 | 1.13 | not significant at p < 0.05. |
| LAG3 | 1.50 | <0.00001 |
| TGFβ1 | 1.46 | <0.00001. |
| TRAIL | 0.44 | <0.00006 |
| GZM | 0.31 | <0.00001 |
| CD39 | 1.45 | <0.00001 |
| CD73 | 0.48 | <0.00001 |
| ICER | 0.63 | <0.01 |
| NFAT | 1.88 | <0.00001 |
| PPRγ | 0.61 | not significant at p < 0.05. |
| p21SNFT | 0.66 | not significant at p < 0.05. |
| GITR | 0.50 | <0.0001 |
| Cbl-b | 1.38 | <0.00001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhaumik, S.; Łazarczyk, M.; Kubick, N.; Klimovich, P.; Gurba, A.; Paszkiewicz, J.; Teodorowicz, P.; Kocki, T.; Horbańczuk, J.O.; Manda, G.; et al. Investigation of the Molecular Evolution of Treg Suppression Mechanisms Indicates a Convergent Origin. Curr. Issues Mol. Biol. 2023, 45, 628-648. https://doi.org/10.3390/cimb45010042
Bhaumik S, Łazarczyk M, Kubick N, Klimovich P, Gurba A, Paszkiewicz J, Teodorowicz P, Kocki T, Horbańczuk JO, Manda G, et al. Investigation of the Molecular Evolution of Treg Suppression Mechanisms Indicates a Convergent Origin. Current Issues in Molecular Biology. 2023; 45(1):628-648. https://doi.org/10.3390/cimb45010042
Chicago/Turabian StyleBhaumik, Suniti, Marzena Łazarczyk, Norwin Kubick, Pavel Klimovich, Agata Gurba, Justyna Paszkiewicz, Patrycja Teodorowicz, Tomasz Kocki, Jarosław Olav Horbańczuk, Gina Manda, and et al. 2023. "Investigation of the Molecular Evolution of Treg Suppression Mechanisms Indicates a Convergent Origin" Current Issues in Molecular Biology 45, no. 1: 628-648. https://doi.org/10.3390/cimb45010042
APA StyleBhaumik, S., Łazarczyk, M., Kubick, N., Klimovich, P., Gurba, A., Paszkiewicz, J., Teodorowicz, P., Kocki, T., Horbańczuk, J. O., Manda, G., Sacharczuk, M., & Mickael, M. -E. (2023). Investigation of the Molecular Evolution of Treg Suppression Mechanisms Indicates a Convergent Origin. Current Issues in Molecular Biology, 45(1), 628-648. https://doi.org/10.3390/cimb45010042