
Role of Stress on Driving the Intestinal Paracellular Permeability

 , and
, and
Abstract
:
1. Introduction
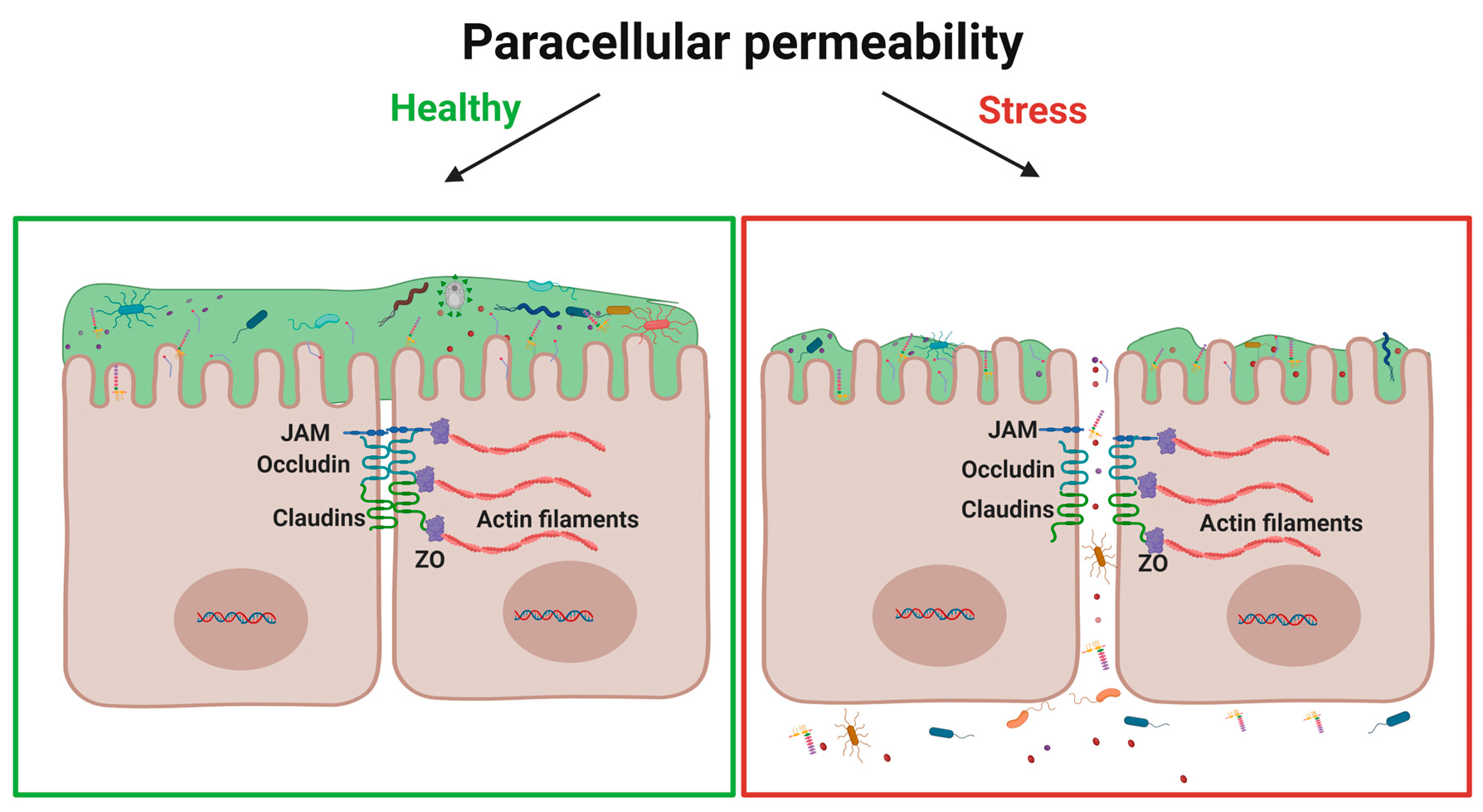
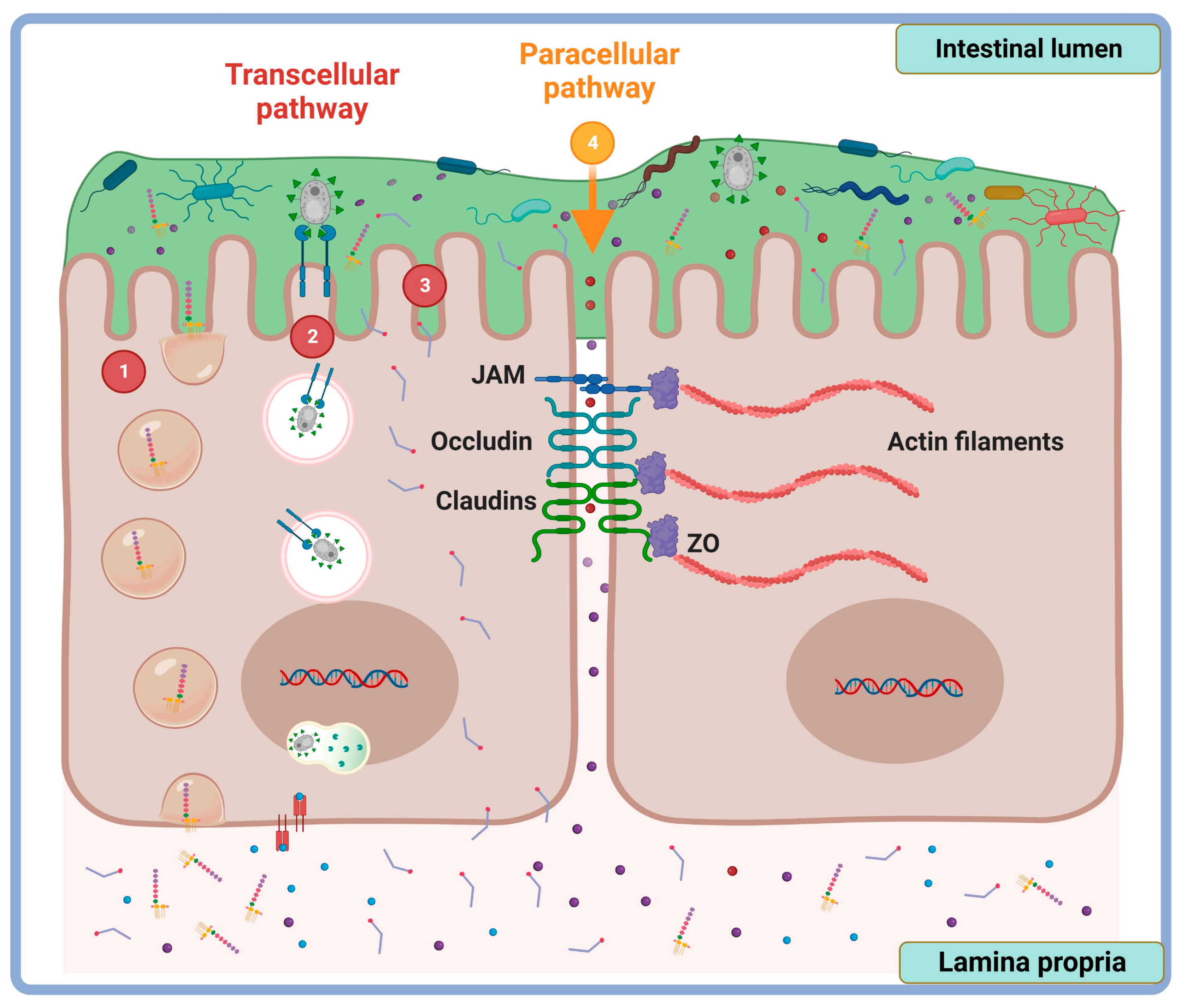
2. Tight Junction Proteins: Overview Structure and Function
3. Methods of Intestinal Permeability Evaluation
3.1. Semiquantitative Assessment of Intestinal Barrier Integrity
3.2. Assessment of Intestinal Permeability In Vivo
3.2.1. From the Intestinal Lumen toward the Blood
3.2.2. From the Blood to the Intestinal Lumen
3.3. Evaluation of Intestinal Permeability In Vitro
3.4. Assessment of Intestinal Barrier Function Ex Vivo
Ussing Chamber Technique
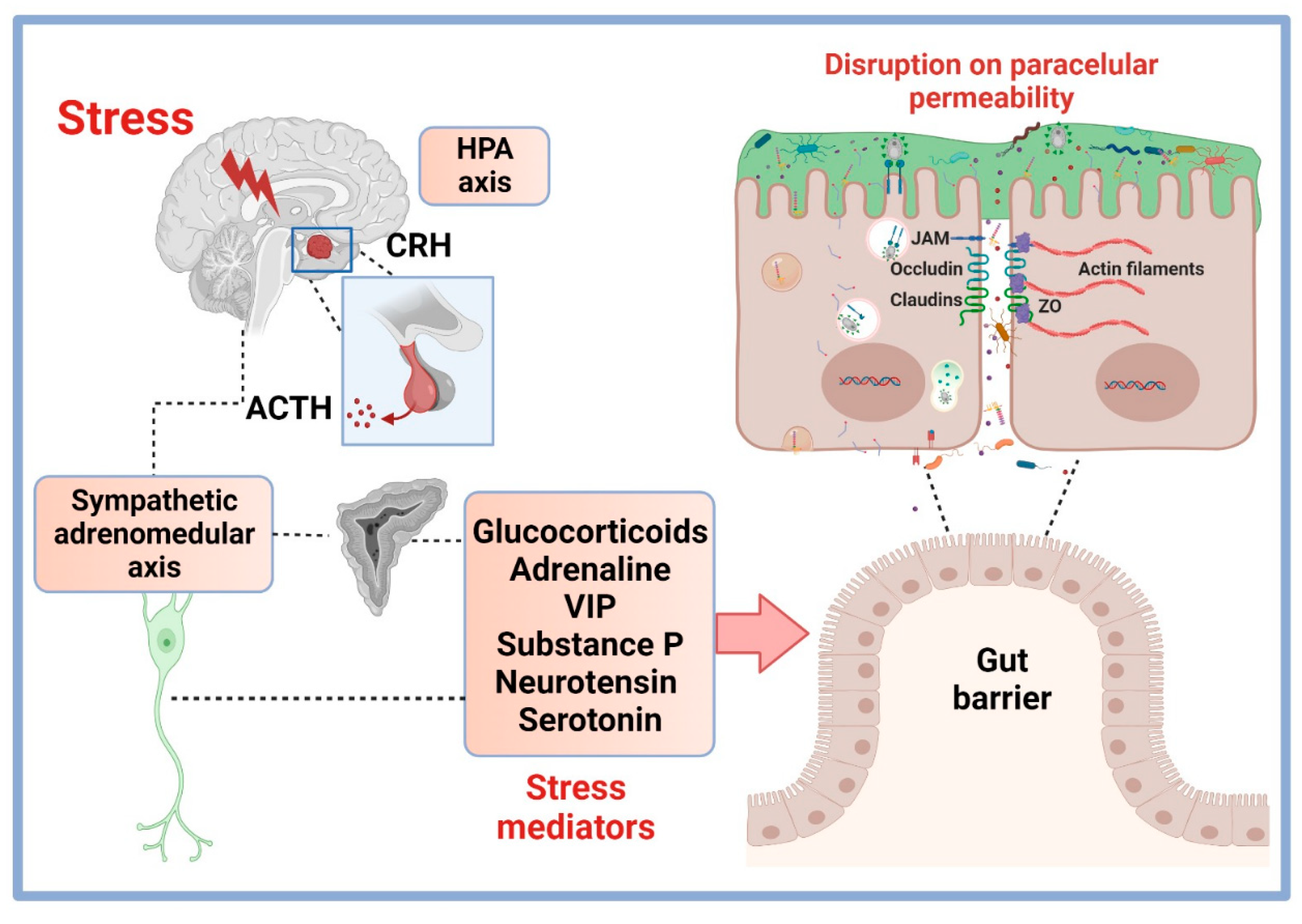
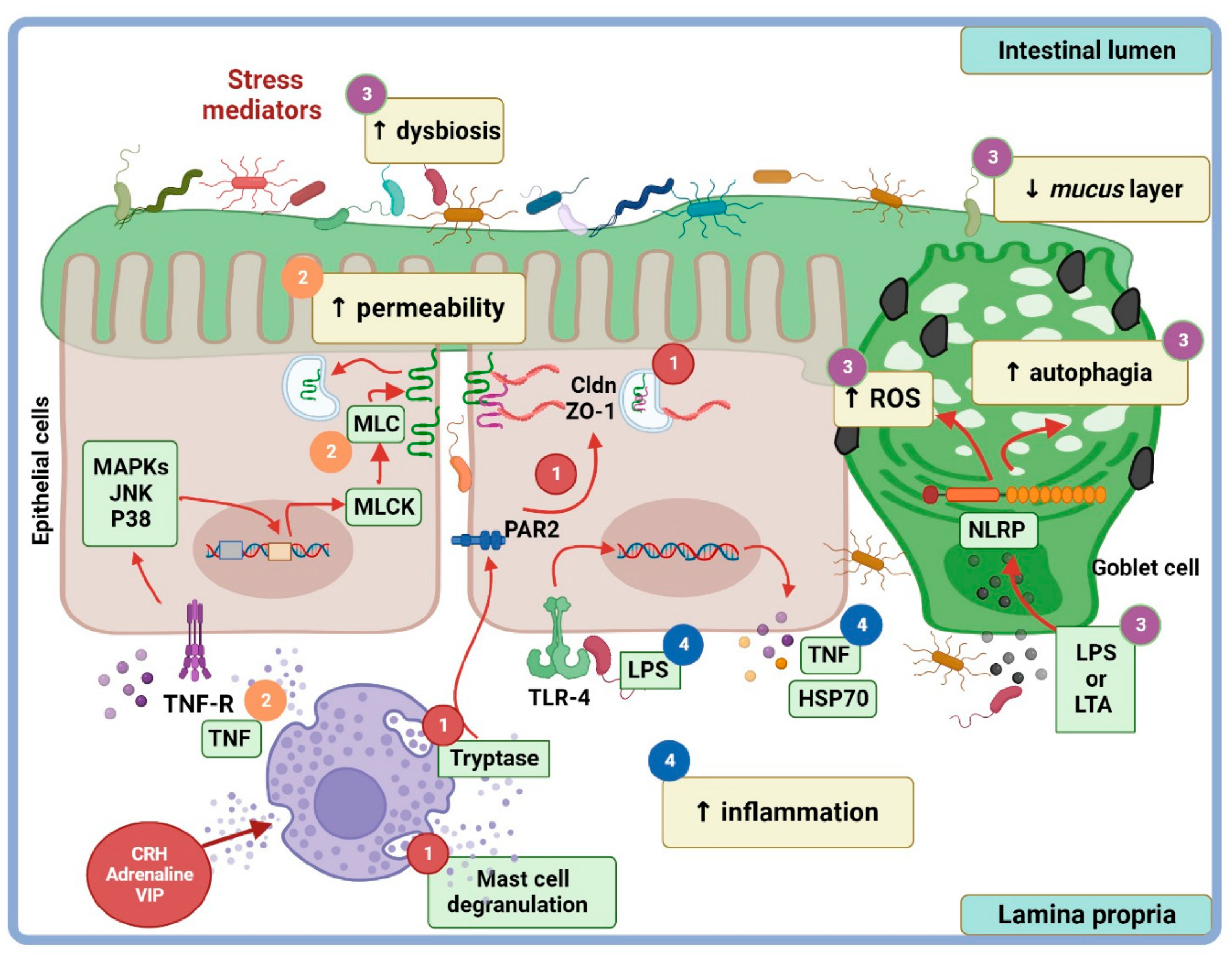
4. Stress Modulation Pathways on Permeability and TJP
4.1. HPA Axis Stress Hormones and Permeability
4.2. Additional Stress Pathways on Paracellular Permeability
5. Clinical Implications of Epithelial Junctions and Intestinal Permeability
6. Perspectives
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef]
- Lu, Z.; Ding, L.; Lu, Q.; Chen, Y.-H. Claudins in intestines: Distribution and functional significance in health. Tissue Barriers 2013, 1, e24978. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef]
- Li, X.J.; You, X.Y.; Wang, C.Y.; Li, X.L.; Sheng, Y.Y.; Zhuang, P.W.; Zhang, Y.J. Bidirectional Brain-gut-microbiota Axis in increased intestinal permeability induced by central nervous system injury. CNS Neurosci. Ther. 2020, 26, 783–790. [Google Scholar] [CrossRef]
- Rodiño-Janeiro, B.K.; Alonso-Cotoner, C.; Pigrau, M.; Lobo, B.; Vicario, M.; Santos, J. Role of corticotropin-releasing factor in gastrointestinal permeability. J. Neurogastroenterol. Motil. 2015, 21, 33–50. [Google Scholar] [CrossRef]
- Tena-Garitaonaindia, M.; Arredondo-Amador, M.; Mascaraque, C.; Asensio, M.; Marin, J.J.G.; Martínez-Augustin, O.; Sánchez de Medina, F. Modulation of intestinal barrier function by glucocorticoids: Lessons from preclinical models. Pharmacol. Res. 2022, 177, 106056. [Google Scholar] [CrossRef]
- Brzozowski, B.; Mazur-Bialy, A.; Pajdo, R.; Kwiecien, S.; Bilski, J.; Zwolinska-Wcislo, M.; Mach, T.; Brzozowski, T. Mechanisms by wich Stress Affects the Experimental and Clinical Inflammatory Bowel Disease (IBD): Role of Brain-Gut Axis. Curr. Neuropharmacol. 2016, 14, 892–900. [Google Scholar] [CrossRef]
- Camilleri, M. Human intestinal barrier: Effects of stressors, diet, prebiotics, and probiotics. Clin. Transl. Gastroenterol. 2021, 12, e00308. [Google Scholar] [CrossRef]
- Graziani, C.; Talocco, C.; De Sire, R.; Petito, V.; Lopetuso, L.R.; Gervasoni, J.; Persichilli, S.; Franceschi, F.; Ojetti, V.; Gasbarrini, A.S.F. Intestinal permeability in physiological and pathological conditions: Major determinants and assessment modalities. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 795–810. [Google Scholar] [PubMed]
- Salvo-Romero, E.; Alonso-Cotoner, C.; Pardo-Camacho, C.; Casado-Bedmar, M.; Vicario, M. The intestinal barrier function and its involvement in digestive disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Dodiya, H.B.; Forsyth, C.B.; Voigt, R.M.; Engen, P.A.; Patel, J.; Shaikh, M.; Green, S.J.; Naqib, A.; Roy, A.; Kordower, J.H.; et al. Chronic stress-induced gut dysfunction exacerbates Parkinson’s disease phenotype and pathology in a rotenone-induced mouse model of Parkinson’s disease. Neurobiol. Dis. 2020, 135, 104352. [Google Scholar] [CrossRef]
- Kelly, J.R.; Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G.; Hyland, N.P. Breaking down the barriers: The gut microbiome, intestinal permeability and stress-related psychiatric disorders. Front. Cell. Neurosci. 2015, 9, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Ait-Belgnaoui, A.; Bradesi, S.; Fioramonti, J.; Theodorou, V.; Bueno, L. Acute stress-induced hypersensitivity to colonic distension depends upon increase in paracellular permeability: Role of myosin light chain kinase. Pain 2005, 113, 141–147. [Google Scholar] [CrossRef]
- Cameron, H.L.; Perdue, M.H. Stress impairs murine intestinal barrier function: Improvement by glucagon-like peptide-2. J. Pharmacol. Exp. Ther. 2005, 314, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Ferrier, L.; Mazelin, L.; Cenac, N.; Desreumaux, P.; Janin, A.; Emilie, D.; Colombel, J.F.; Garcia-Villar, R.; Fioramonti, J.; Bueno, L. Stress-induced disruption of colonic epithelial barrier: Role of interferon-γ and myosin light chain kinase in mice. Gastroenterology 2003, 125, 795–804. [Google Scholar] [CrossRef]
- Keita, Å.V.; Söderholm, J.D.; Ericson, A.C. Stress-induced barrier disruption of rat follicle-associated epithelium involves corticotropin-releasing hormone, acetylcholine, substance P, and mast cells. Neurogastroenterol. Motil. 2010, 22, 770-e222. [Google Scholar] [CrossRef]
- Zheng, P.Y.; Feng, B.S.; Oluwole, C.; Struiksma, S.; Chen, X.; Li, P.; Tang, S.G.; Yang, P.C. Psychological stress induces eosinophils to produce corticotrophin releasing hormone in the intestine. Gut 2009, 58, 1473–1479. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, D.K.; Kim, Y.B.; Lee, K.J. Effect of acute stress on immune cell counts and the expression of tight junction proteins in the duodenal mucosa of rats. Gut Liver 2013, 7, 190–196. [Google Scholar] [CrossRef]
- Lin, R.; Wang, Z.; Cao, J.; Gao, T.; Dong, Y.; Chen, Y. Role of melatonin in intestinal mucosal injury induced by restraint stress in mice. Pharm. Biol. 2020, 58, 342–351. [Google Scholar] [CrossRef]
- Mazzon, E.; Cuzzocrea, S. Role of TNF-in ileum tight junction alteration in mouse model of restraint stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1268–G1280. [Google Scholar] [CrossRef] [PubMed]
- Cummins, P.M. Occludin: One Protein, Many Forms. Mol. Cell. Biol. 2012, 32, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Schoultz, I.; Keita, Å.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef]
- Richter, J.F.; Hildner, M.; Schmauder, R.; Turner, J.R.; Schumann, M.; Reiche, J. Occludin knockdown is not sufficient to induce transepithelial macromolecule passage. Tissue Barriers 2019, 7, 1612661. [Google Scholar] [CrossRef] [PubMed]
- Morita, K.; Furuse, M.; Fujimoto, K.; Tsukita, S. Claudin multigene family encoding four-transmembrane domain protein components of tight junction strands. Cell Biol. 1999, 96, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.M.; Amasheh, S.; Richter, J.F.; Milatz, S.; Gü, D.; Westphal, J.K.; Huber, O.; Schulzke, J.D.; Fromm, M. Tricellulin Forms a Barrier to Macromolecules in Tricellular Tight Junctions without Affecting Ion Permeability. Mol. Biol. Cell 2009, 20, 3713–3724. [Google Scholar] [CrossRef]
- Overgaard, C.E.; Daugherty, B.L.; Mitchell, L.A.; Koval, M. Claudins: Control of barrier function and regulation in response to oxidant stress. Antioxid. Redox Signal. 2011, 15, 1179–1193. [Google Scholar] [CrossRef]
- Günzel, D.; Amasheh, S.; Pfaffenbach, S.; Richter, J.F.; Kausalya, J.P.; Hunziker, W.; Fromm, M. Claudin-16 affects transcellular Cl- secretion in MDCK cells. J. Physiol. 2009, 587, 3777–3793. [Google Scholar] [CrossRef]
- Günzel, D.; Fromm, M. Claudins and other tight junction proteins. Compr. Physiol. 2012, 2, 1819–1852. [Google Scholar] [CrossRef]
- Krause, G.; Winkler, L.; Mueller, S.L.; Haseloff, R.F.; Piontek, J.; Blasig, I.E. Structure and function of claudins. Biochim. Biophys. Acta-Biomembr. 2008, 1778, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Markov, A.G.; Aschenbach, J.R.; Amasheh, S. Claudin clusters as determinants of epithelial barrier function. IUBMB Life 2015, 67, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Amasheh, S.; Fromm, M.; Günzel, D. Claudins of intestine and nephron-a correlation of molecular tight junction structure and barrier function. Acta Physiol. 2011, 201, 133–140. [Google Scholar] [CrossRef]
- Holmes, J.L.; Van Itallie, C.M.; Rasmussen, J.E.; Anderson, J.M. Claudin profiling in the mouse during postnatal intestinal development and along the gastrointestinal tract reveals complex expression patterns. Gene Expr. Patterns 2006, 6, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.A.; Nelson, W.J.; Chavez, N. Cell–cell junctions organize structural and signaling networks. Cold Spring Harb. Perspect. Biol. 2018, 10, a029181. [Google Scholar] [CrossRef] [PubMed]
- Nusrat, A.; Turner, J.R.; Madara, J.L. Molecular Physiology and Pathophysiology of Tight Junctions IV. Regulation of tight junctions by extracellular stimuli: Nutrients, cytokines, and immune cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G851–G857. [Google Scholar] [CrossRef]
- Takeichi, M. Dynamic contacts: Rearranging adherens junctions to drive epithelial remodelling. Nat. Rev. Mol. Cell Biol. 2014, 15, 397–410. [Google Scholar] [CrossRef]
- Goodenough, D.A.; Paul, D.L. Gap junctions. Cold Spring Harb. Perspect. Biol. 2009, 1, a002576. [Google Scholar] [CrossRef]
- Hamilton, J.A. Fatty acid transport: Difficult or easy? James. J. Lipid Res. 1998, 39, 467–481. [Google Scholar] [CrossRef]
- Moraes, T.F.; Reithmeier, R.A.F. Membrane transport metabolons. Biochim. Biophys. Acta-Biomembr. 2012, 1818, 2687–2706. [Google Scholar] [CrossRef]
- Doherty, G.J.; McMahon, H.T. Mechanisms of endocytosis. Annu. Rev. Biochem. 2009, 78, 857–902. [Google Scholar] [CrossRef] [PubMed]
- Hollander, D.; Kaunitz, J.D. The “Leaky Gut”: Tight Junctions but Loose Associations? Dig. Dis. Sci. 2020, 65, 1277–1287. [Google Scholar] [CrossRef] [PubMed]
- Pappenheimer, J.R.; Reiss, K.Z. Contribution of Solvent Drag through Intercellular Junctions to Absorption of Nutrients by the Small Intestine of the Rat. J. Membr. Biol. 1987, 100, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Grootjans, J. Non-invasive assessment of barrier integrity and function of the human gut. World J. Gastrointest. Surg. 2010, 2, 61. [Google Scholar] [CrossRef]
- Wang, L.; Llorente, C.; Hartmann, P.; Yang, A.M.; Chen, P.; Schnabl, B. Methods to determine intestinal permeability and bacterial translocation during liver disease. J. Immunol. Methods 2015, 421, 44–53. [Google Scholar] [CrossRef]
- Miglietta, S.; Borghini, R.; Relucenti, M.; Sorrentino, V.; Chen, R.; Li, X.; Fazi, F.; Donato, G.; Familiari, G.; Petrozza, V.; et al. New Insights into Intestinal Permeability in Irritable Bowel Syndrome-Like Disorders: Histological and Ultrastructural Findings of Duodenal Biopsies. Cells 2021, 10, 2593. [Google Scholar] [CrossRef]
- Zeissig, S.; Bürgel, N.; Günzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61–72. [Google Scholar] [CrossRef]
- Boschetti, E.; Accarino, A.; Malagelada, C.; Malagelada, J.R.; Cogliandro, R.F.; Gori, A.; Tugnoli, V.; Giancola, F.; Bianco, F.; Bonora, E.; et al. Gut epithelial and vascular barrier abnormalities in patients with chronic intestinal pseudo-obstruction. Neurogastroenterol. Motil. 2019, 31, e13652. [Google Scholar] [CrossRef]
- Tabat, M.W.; Marques, T.M.; Markgren, M.; Löfvendahl, L.; Brummer, R.J.; Wall, R. Acute effects of butyrate on induced hyperpermeability and tight junction protein expression in human colonic tissues. Biomolecules 2020, 10, 766. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, X.; Andersson, R. Role of Intestinal Permeability in Monitoring Mucosal Barrier Function History, Methodology, and Significance of Pathophysiology. Dig. Surg. 1998, 15, 386–397. [Google Scholar] [CrossRef]
- Thomson, A.; Smart, K.; Somerville, M.S.; Lauder, S.N.; Appanna, G.; Horwood, J.; Sunder Raj, L.; Srivastava, B.; Durai, D.; Scurr, M.J.; et al. The Ussing chamber system for measuring intestinal permeability in health and disease. BMC Gastroenterol. 2019, 19, 98. [Google Scholar] [CrossRef] [PubMed]
- Sequeira, I.R.; Lentle, R.G.; Kruger, M.C.; Hurst, R.D. Standardising the lactulose mannitol test of gut permeability to minimise error and promote comparability. PLoS ONE 2014, 9, e99256. [Google Scholar] [CrossRef] [PubMed]
- Dastych, M.; Dastych, M.; Novotná, H.; Číhalová, J. Lactulose/mannitol test and specificity, sensitivity, and area under curve of intestinal permeability parameters in patients with liver cirrhosis and Crohn’s disease. Dig. Dis. Sci. 2008, 53, 2789–2792. [Google Scholar] [CrossRef] [PubMed]
- Vanuytsel, T.; Tack, J.; Farre, R. The Role of Intestinal Permeability in Gastrointestinal Disorders and Current Methods of Evaluation. Front. Nutr. 2021, 8, 717925. [Google Scholar] [CrossRef] [PubMed]
- Vojdani, A. For the Assessment of Intestinal Permeability, Size Matters. Altern. Ther. Health Med. 2013, 19, 12–24. [Google Scholar] [PubMed]
- Schäfer, T.V.; Ivnitsky, J.J.; Rejniuk, V.L. Permeability of Intestinal and Blood—Tissue Barriers in Rats for Evans Blue Dye under Conditions of Acute Intoxication with Cyclophosphamide. Bull. Exp. Biol. Med. 2019, 168, 38–40. [Google Scholar] [CrossRef]
- Jung, H.P.; Dong, I.P.; Hong, J.K.; Yong, K.C.; Chong, I.S.; Woo, K.J.; Byung, I.K.; Kyoung, H.W.; Soon, M.P. The relationship between small-intestinal bacterial overgrowth and intestinal permeability in patients with irritable bowel syndrome. Gut Liver 2009, 3, 174–179. [Google Scholar] [CrossRef]
- Clayburgh, D.R.; Barrett, T.A.; Tang, Y.; Meddings, J.B.; Van Eldik, L.J.; Watterson, D.M.; Clarke, L.L.; Mrsny, R.J.; Turner, J.R. Epithelial myosin light chain kinase-dependent barrier dysfunction mediates T cell activation-induced diarrhea in vivo. J. Clin. Investig. 2005, 115, 2702–2715. [Google Scholar] [CrossRef]
- Radu, M.; Chernoff, J. An in vivo Assay to Test Blood Vessel Permeability. J. Vis. Exp. 2013, 73, e50062. [Google Scholar] [CrossRef]
- Dahlgren, D.; Roos, C.; Lundqvist, A.; Tannergren, C.; Langguth, P.; Sjöblom, M.; Sjögren, E.; Lennernäs, H. Preclinical Effect of Absorption Modifying Excipients on Rat Intestinal Transport of Model Compounds and the Mucosal Barrier Marker 51Cr-EDTA. Mol. Pharm. 2017, 14, 4243–4251. [Google Scholar] [CrossRef]
- Ghiselli, F.; Rossi, B.; Piva, A.; Grilli, E. Assessing Intestinal Health. In Vitro and Ex vivo Gut Barrier Models of Farm Animals: Benefits and Limitations. Front. Vet. Sci. 2021, 8, 723387. [Google Scholar] [CrossRef] [PubMed]
- Balimane, P.V.; Chong, S. Cell culture-based models for intestinal permeability: A critique. Drug Discov. Today 2005, 10, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Zweibaum, A.; Triadou, N.; Kedinger, M.; Augeron, C.; Robine-Léon, S.; Pinto, M.; Rousset, M.H.K. Sucrase-isomaltase: A marker of foetal and malignant epithelial cells of the human colon. Int. J. Cancer 1983, 32, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Juuti-Uusitalo, K.M.; Kaukinen, K.; Mäki, M.; Tuimala, J.; Kainulainen, H. Gene expression in TGFbeta-induced epithelial cell differentiation in a three-dimensional intestinal epithelial cell differentiation model. BMC Genom. 2006, 7, 279. [Google Scholar] [CrossRef]
- Devriese, S.; Van den Bossche, L.; Van Welden, S.; Holvoet, T.; Pinheiro, I.; Hindryckx, P.; De Vos, M.; Laukens, D. T84 monolayers are superior to Caco-2 as a model system of colonocytes. Histochem. Cell Biol. 2017, 148, 85–93. [Google Scholar] [CrossRef]
- Langerholc, T.; Maragkoudakis, P.A.; Wollgast, J.; Gradisnik, L.; Cencic, A. Novel and established intestinal cell line models-An indispensable tool in food science and nutrition. Trends Food Sci. Technol. 2011, 22, S11–S20. [Google Scholar] [CrossRef]
- CenciČ, A.; Langerholc, T. Functional cell models of the gut and their applications in food microbiology—A review. Int. J. Food Microbiol. 2010, 141, S4–S14. [Google Scholar] [CrossRef]
- Westerhout, J.; Wortelboer, H.; Verhoeckx, K. Ussing Chamber. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 263–273. ISBN 9783319161044. [Google Scholar]
- Mateer, S.W.; Cardona, J.; Marks, E.; Goggin, B.J.; Hua, S.; Keely, S. Ex vivo intestinal sacs to assess mucosal permeability in models of gastrointestinal disease. J. Vis. Exp. 2016, 2016, 53250. [Google Scholar] [CrossRef]
- Wilcz-Villega, E.M.; McClean, S.; O’Sullivan, M.A. Mast cell tryptase reduces junctional adhesion molecule-A (JAM-A) expression in intestinal epithelial cells: Implications for the mechanisms of barrier dysfunction in irritable bowel syndrome. Am. J. Gastroenterol. 2013, 108, 1140–1151. [Google Scholar] [CrossRef]
- Wallon, C.; Braaf, Y.; Wolving, M.; Olaison, G.; Söderholm, J.D. Endoscopic biopsies in Ussing chambers evaluated for studies of macromolecular permeability in the human colon. Scand. J. Gastroenterol. 2005, 40, 586–595. [Google Scholar] [CrossRef]
- Katinios, G.; Casado-Bedmar, M.; Walter, S.A.; Vicario, M.; González-Castro, A.M.; Bednarska, O.; Söderholm, J.D.; Hjortswang, H.; Keita, Å.V. Increased Colonic Epithelial Permeability and Mucosal Eosinophilia in Ulcerative Colitis in Remission Compared with Irritable Bowel Syndrome and Health. Inflamm. Bowel Dis. 2020, 26, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Piche, T.; Barbara, G.; Aubert, P.; Des Varannes, S.B.; Dainese, R.; Nano, J.L.; Cremon, C.; Stanghellini, V.; De Giorgio, R.; Galmiche, J.P.; et al. Impaired Intestinal barrier integrity in the colon of patients with irritable bowel syndrome: Involvement of soluble mediators. Gut 2009, 58, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Lance, S.; Delbro, D.S.; Jennische, E.; Delbro, D.S. Evans Blue Permeation of Intestinal Mucosa in the Rat. Scand. J. Gastroenterol. 1994, 29, 38–46. [Google Scholar]
- Wilms, E.; Troost, F.J.; Elizalde, M.; Winkens, B.; de Vos, P.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M. Intestinal barrier function is maintained with aging–a comprehensive study in healthy subjects and irritable bowel syndrome patients. Sci. Rep. 2020, 10, 475. [Google Scholar] [CrossRef]
- Godoy, L.D.; Rossignoli, M.T.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E.H. A comprehensive overview on stress neurobiology: Basic concepts and clinical implications. Front. Behav. Neurosci. 2018, 12, 127. [Google Scholar] [CrossRef]
- Belda, X.; Fuentes, S.; Daviu, N.; Nadal, R.; Armario, A. Stress-induced sensitization: The hypothalamic-pituitary-adrenal axis and beyond. Stress 2015, 18, 269–279. [Google Scholar] [CrossRef]
- Goldstein, D.S. Stress and the “extended” autonomic system. Auton. Neurosci. Basic Clin. 2021, 236, 102889. [Google Scholar] [CrossRef]
- Lauffer, A.; Vanuytsel, T.; Vanormelingen, C.; Vanheel, H.; Salim Rasoel, S.; Tóth, J.; Tack, J.; Fornari, F.; Farré, R. Subacute stress and chronic stress interact to decrease intestinal barrier function in rats. Stress 2016, 19, 225–234. [Google Scholar] [CrossRef]
- Cima, I.; Corazza, N.; Dick, B.; Fuhrer, A.; Herren, S.; Jakob, S.; Ayuni, E.; Mueller, C.; Brunner, T. Intestinal epithelial cells synthesize glucocorticoids and regulate T cell activation. J. Exp. Med. 2004, 200, 1635–1646. [Google Scholar] [CrossRef]
- Zheng, G.; Wu, S.P.; Hu, Y.; Smith, D.E.; Wiley, J.W.; Hong, S. Corticosterone mediates stress-related increased intestinal permeability in a region-specific manner. Neurogastroenterol. Motil. 2013, 25, e127–e139. [Google Scholar] [CrossRef]
- Zheng, G.; Victor Fon, G.; Meixner, W.; Creekmore, A.; Zong, Y.; Michael Dame, M.K.; Colacino, J.; Dedhia, P.H.; Hong, S.; Wiley, J.W. Chronic stress and intestinal barrier dysfunction: Glucocorticoid receptor and transcription repressor HES1 regulate tight junction protein Claudin-1 promoter. Sci. Rep. 2017, 7, 4502. [Google Scholar] [CrossRef] [PubMed]
- Boivin, M.A.; Ye, D.; Kennedy, J.C.; Al-Sadi, R.; Shepela, C.; Ma, T.Y. Mechanism of glucocorticoid regulation of the intestinal tight junction barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Gluth, M.; Weege, F.; Pape, U.-F.; Wiedenmann, B.; Baumgart, D.C.; Theuring, F. Glucocorticoids regulate barrier function and claudin expression in intestinal epithelial cells via MKP-1. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shepherd, E.G.; Nelin, L.D. MAPK phosphatases-Regulating the immune response. Nat. Rev. Immunol. 2007, 7, 202–212. [Google Scholar] [CrossRef]
- Matsuo, K.; Zhang, X.; Ono, Y.; Nagatomi, R. Acute stress-induced colonic tissue HSP70 expression requires commensal bacterial components and intrinsic glucocorticoid. Brain Behav. Immun. 2009, 23, 108–115. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, Z.Q.; Liu, X.Y.; Yang, L.; Geng, X.R.; Yang, G.; Liu, Z.G.; Zheng, P.Y.; Yang, P.C. Stress-Derived Corticotropin Releasing Factor Breaches Epithelial Endotoxin Tolerance. PLoS ONE 2013, 8, e65760. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of Commensal Microflora by Toll-Like Receptors Is Required for Intestinal Homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef]
- Rumio, C.; Besusso, D.; Arnaboldi, F.; Palazzo, M.; Selleri, S.; Gariboldi, S.; Akira, S.; Uematsu, S.; Bignami, P.; Ceriani, V.; et al. Activation of smooth muscle and myenteric plexus cells of jejunum via Toll-like receptor 4. J. Cell. Physiol. 2006, 208, 47–54. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal barrier dysfunction, LPS translocation, and disease development. J. Endocr. Soc. 2020, 4, bvz039. [Google Scholar] [CrossRef]
- Nébot-Vivinus, M.; Harkat, C.; Bzioueche, H.; Cartier, C.; Plichon-Dainese, R.; Moussa, L.; Eutamene, H.; Pishvaie, D.; Holowacz, S.; Seyrig, C.; et al. Multispecies probiotic protects gut barrier function in experimental models. World J. Gastroenterol. 2014, 20, 6832–6843. [Google Scholar] [CrossRef]
- Yu, J.; Liu, T.; Guo, Q.; Wang, Z.; Chen, Y.; Dong, Y. Disruption of the Intestinal Mucosal Barrier Induced by High Fructose and Restraint Stress Is Regulated by the Intestinal Microbiota and Microbiota Metabolites. Microbiol. Spectr. 2023, 11, e0469822. [Google Scholar] [CrossRef] [PubMed]
- Almeida-da-Silva, C.L.C.; Savio, L.E.B.; Coutinho-Silva, R.; Ojcius, D.M. The role of NOD-like receptors in innate immunity. Front. Immunol. 2023, 14, 1122586. [Google Scholar] [CrossRef] [PubMed]
- Larabi, A.; Barnich, N.; Nguyen, H.T.T. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in IBD. Autophagy 2020, 16, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Mizugaki, A.; Inoue, Y.; Kato, H.; Murakami, H. Cystine reduces tight junction permeability and intestinal inflammation induced by oxidative stress in Caco-2 cells. Amino Acids 2021, 53, 1021–1032. [Google Scholar] [CrossRef]
- Geerlings, S.Y.; Kostopoulos, I.; de Vos, W.M.; Belzer, C. Akkermansia muciniphila in the human gastrointestinal tract: When, where, and how? Microorganisms 2018, 6, 75. [Google Scholar] [CrossRef]
- Grabacka, M.; Płonka, P.M.; Pierzchalska, M. The PPARα Regulation of the Gut Physiology in Regard to Interaction with Microbiota, Intestinal Immunity, Metabolism, and Permeability. Int. J. Mol. Sci. 2022, 23, 14156. [Google Scholar] [CrossRef]
- Mirza, A.Z.; Althagafi, I.I.; Shamshad, H. Role of PPAR receptor in different diseases and their ligands: Physiological importance and clinical implications. Eur. J. Med. Chem. 2019, 166, 502–513. [Google Scholar] [CrossRef]
- Ponferrada, Á.; Caso, J.R.; Alou, L.; Colón, A.; Sevillano, D.; Moro, M.A.; Lizasoain, I.; Menchén, P.; Gómez-Lus, M.L.; Lorenzo, P.; et al. The Role of PPARγ on Restoration of Colonic Homeostasis after Experimental Stress-Induced Inflammation and Dysfunction. Gastroenterology 2007, 132, 1791–1803. [Google Scholar] [CrossRef]
- Mazzon, E.; Crisafulli, C.; Galuppo, M.; Cuzzocrea, S. Role of peroxisome proliferator-activated receptor-in ileum tight junction alteration in mouse model of restraint stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, 488–505. [Google Scholar] [CrossRef]
- Sharkey, K.A.; Wiley, J.W. The Role of the Endocannabinoid System in the Brain–Gut Axis. Gastroenterology 2016, 151, 252–266. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, X.; Yang, C.; Zhao, S. Effect of monoacylglycerol lipase inhibition on intestinal permeability in chronic stress model. Biochem. Biophys. Res. Commun. 2020, 525, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Song, J.; Hou, X. Mast cells and irritable bowel syndrome: From the bench to the bedside. J. Neurogastroenterol. Motil. 2016, 22, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Chelombitko, M.A.; Fedorov, A.V.; Ilyinskaya, O.P.; Zinovkin, R.A.; Chernyak, B.V. Role of reactive oxygen species in mast cell degranulation. Biochemistry 2016, 81, 1564–1577. [Google Scholar] [CrossRef] [PubMed]
- Mazzon, E.; Sturniolo, G.; Puzzolo, D.; Frisina, N.; Fries, W. Effect of stress on the paracellular barrier in the rat ileum. Small Intest. 2002, 51, 507–513. [Google Scholar] [CrossRef]
- Hattay, P.; Prusator, D.K.; Tran, L.; Greenwood-Van Meerveld, B. Psychological stress-induced colonic barrier dysfunction: Role of immune-mediated mechanisms. Neurogastroenterol. Motil. 2017, 29, e13043. [Google Scholar] [CrossRef]
- Demaude, J.; Salvador-Cartier, C.; Fioramonti, J.; Ferrier, L.; Bueno, L. Phenotypic changes in colonocytes following acute stress or activation of mast cells in mice: Implications for delayed epithelial barrier dysfunction. Gut 2006, 55, 655–661. [Google Scholar] [CrossRef]
- Jacob, C.; Yang, P.C.; Darmoul, D.; Amadesi, S.; Saito, T.; Cottrell, G.S.; Coelho, A.M.; Singh, P.; Grady, E.F.; Perdue, M.; et al. Mast cell tryptase controls paracellular permeability of the intestine: Role of protease-activated receptor 2 and β-arrestins. J. Biol. Chem. 2005, 280, 31936–31948. [Google Scholar] [CrossRef]
- Cenac, N.; Chin, A.C.; Garcia-Villar, R.; Salvador-Cartier, C.; Ferrier, L.; Vergnolle, N.; Buret, A.G.; Fioramonti, J.; Bueno, L. PAR2 activation alters colonic paracellular permeability in mice via IFN-γ-dependent and -independent pathways. J. Physiol. 2004, 558, 913–925. [Google Scholar] [CrossRef]
- Drucker, D.J. Biological actions and therapeutic potential of the glucagon-like peptides. Gastroenterology 2002, 122, 531–544. [Google Scholar] [CrossRef]
- Jamison, J.R. Irritable Bowel Syndrome. In Clinical Guide to Nutrition & Dietary Supplements in Disease Management; Churchill Livingstone: Saint Louis, MO, USA, 2003; Volume 117, pp. 329–332. [Google Scholar] [CrossRef]
- Ford, A.C.; Lacy, B.E.; Talley, N.J. Irritable Bowel Syndrome. Rev. Prat. 2017, 376, 2566–2578. [Google Scholar] [CrossRef]
- Buhner, S.; Li, Q.; Vignali, S.; Barbara, G.; De Giorgio, R.; Stanghellini, V.; Cremon, C.; Zeller, F.; Langer, R.; Daniel, H.; et al. Activation of Human Enteric Neurons by Supernatants of Colonic Biopsy Specimens from Patients with Irritable Bowel Syndrome. Gastroenterology 2009, 137, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Liebregts, T.; Adam, B.; Bredack, C.; Röth, A.; Heinzel, S.; Lester, S.; Downie-Doyle, S.; Smith, E.; Drew, P.; Talley, N.J.; et al. Immune Activation in Patients With Irritable Bowel Syndrome. Gastroenterology 2007, 132, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Vargas, N.N.; Pattison, L.A.; Zhao, P.; Lieu, T.M.; Latorre, R.; Jensen, D.D.; Castro, J.; Aurelio, L.; Le, G.T.; Flynn, B.; et al. Protease-activated receptor-2 in endosomes signals persistent pain of irritable bowel syndrome. Proc. Natl. Acad. Sci. USA 2018, 115, E7438–E7447. [Google Scholar] [CrossRef] [PubMed]
- Enck, P.; Aziz, Q.; Barbara, G.; Farmer, A.D.; Fukudo, S.; Mayer, E.A.; Niesler, B.; Quigley, E.M.M.; Rajilić-Stojanović, M.; Schemann, M.; et al. Irritable bowel syndrome. Nat. Rev. Dis. Prim. 2016, 2, 1–24. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; Scaldaferri, F.; Bruno, G.; Petito, V.; Franceschi, F.; Gasbarrini, A. The therapeutic management of gut barrier leaking. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1068–1076. [Google Scholar]
- Simrén, M.; Tack, J. New treatments and therapeutic targets for IBS and other functional bowel disorders. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 589–605. [Google Scholar] [CrossRef]
- Bonetto, S.; Fagoonee, S.; Battaglia, E.; Grassini, M.; Saracco, G.M.; Pellicano, R. Recent advances in the treatment of irritable bowel syndrome. Polish Arch. Intern. Med. 2021, 131, 709–715. [Google Scholar] [CrossRef]
- Zong, Y.; Zhu, S.; Zhang, S.; Zheng, G.; Wiley, J.W.; Hong, S. Chronic stress and intestinal permeability: Lubiprostone regulates glucocorticoid receptor-mediated changes in colon epithelial tight junction proteins, barrier function, and visceral pain in the rodent and human. Neurogastroenterol. Motil. 2019, 31, e13477. [Google Scholar] [CrossRef]
- Van Spaendonk, H.; Ceuleers, H.; Witters, L.; Patteet, E.; Joossens, J.; Augustyns, K.; Lambeir, A.M.; De Meester, I.; De Man, J.G.; De Winter, B.Y. Regulation of intestinal permeability: The role of proteases. World J. Gastroenterol. 2017, 23, 2106–2123. [Google Scholar] [CrossRef]
- Santos, J.; Yates, D.; Guilarte, M.; Vicario, M.; Alonso, C.; Perdue, M.H. Stress neuropeptides evoke epithelial responses via mast cell activation in the rat colon. Psychoneuroendocrinology 2008, 33, 1248–1256. [Google Scholar] [CrossRef]
- Scudamore, C.L.; Jepson, M.A.; Hirst, B.H.; Miller, H.R.P. The rat mucosal mast cell chymase, RMCP-II, alters epithelial cell monolayer permeability in association with altered distribution of the tight junction proteins ZO-1 and occludin. Eur. J. Cell Biol. 1998, 75, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Vicario, M.; Guilarte, M.; Alonso, C.; Yang, P.; Martínez, C.; Ramos, L.; Lobo, B.; González, A.; Guilà, M.; Pigrau, M.; et al. Chronological assessment of mast cell-mediated gut dysfunction and mucosal inflammation in a rat model of chronic psychosocial stress. Brain. Behav. Immun. 2010, 24, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, J.; Dong, L.; Shi, H.; Wang, Z.; Ding, H.; Shi, H.; Lu, X. A protease inhibitor against acute stress-induced visceral hypersensitivity and paracellular permeability in rats. Eur. J. Pharmacol. 2011, 654, 289–294. [Google Scholar] [CrossRef]
- Lu, L.; Yan, L.; Yuan, J.; Ye, Q.; Lin, J. Shuganyin decoction improves the intestinal barrier function in a rat model of irritable bowel syndrome induced by water-avoidance stress. Chin. Med. 2018, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Mehranfard, N.; Yazdi, A.; Rafiei, A.; Shakerin, Z.; Ghasemi, M. Honey protects against chronic unpredictable mild stress induced- intestinal barrier disintegration and hepatic inflammation. Mol. Biol. Rep. 2020, 47, 8475–8484. [Google Scholar] [CrossRef]
- Zoppi, S.; Madrigal, J.L.M.; Pérez-Nievas, B.G.; Marín-Jiménez, I.; Caso, J.R.; Alou, L.; García-Bueno, B.; Colón, A.; Manzanares, J.; Luisa Gómez-Lus, M.; et al. Endogenous cannabinoid system regulates intestinal barrier function in vivo through cannabinoid type 1 receptor activation. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, 565–571. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker (MW) 1, Administration Route and Permeability Pathway | Sample and Method of Detection | Comments |
|---|---|---|
| Sugars Mannitol (182.2 g/mol), oral route, TCP 2 L-rhamnose (164.16 g/mol), oral route, TCP Lactulose (342.3 g/mol), oral route, PCP 3 Cellobiose (342.3 g/mol), oral route, PCP [52] | Urine GC 4 and/or HPLC 5 | Combination of lactulose/mannitol (L/M) determinates both TCP and PCP; mannitol/L-rhamnose to evaluate TCP and lactulose/cellobiose to evaluate PCP [53]. L/M are degraded by the colonic microbiota, therefore is not recommended to evaluate the colonic permeability [54]. L/M test does not reflect damage of the TJPs 6 [55]. |
| Fluorochrome FITC 7-dextran conjugate (3–2000 kDa), oral route, PCP [45] | Plasma Fluorescence spectroscopy | Permeability to high size of FITC-dextran reflects damage to TJPs structure [55]. The standardization to detection is difficult due depends critically on the dextran molecular weight [45]. |
| Dye EB 8 (960.8 g/mol), jejunal loop injection, PCP [56] | Blood Spectrophotometry at λ = 620 nm | Extent of tissue damage can be visualized macroscopically. After administration, some tissues stain strongly stained so that are useless for histological evaluation [56]. |
| Polymers PEG 9 (200–8000 Da), oral route, PCP [57] | Urine LC-MS 7 method | PEG probe is suitable marker for gut permeability evaluation in humans [57]. |
| Marker (MW) 1, Administration Route and Permeability Pathway | Sample and Method of Detection | Comments |
|---|---|---|
| HAS 2 (67 kDa) (endogenous marker), PCP 3 [45] | Stool samples Standard ELISA 4 test | Blood levels must be normal. Not useful in end-stage liver disease and kidney disease [45]. |
| Alexa Fluor 488 conjugate BSA 5 (66 kDa), I.V. 6 infusion, PCP [58] | Intestinal tissue Fluorescent microplate reader | This intestinal permeability test is usually used to evaluate the effects of drugs on intestinal permeability [45]. |
| EB 7 (960.8 g/mol), I.V. infusion, PCP [56,59] | Intestinal tissue Spectrophotometry at λ = 620 nm | Relatively simple and affordable assay. EA dye is altered by the concentration of BSA in blood that passes through the epithelial tissue. |
| Radioactive chromium bound to EDTA 8 (51Cr-EDTA) (51.9961 g/mol), I.V. infusion, PCP [24,60] | Intestinal tissue After radiation of 0.12 mSv, 51Cr-EDTA is measured by gamma-counting | The strong binding of 51Cr with EDTA prevents its interaction with other ions. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molotla-Torres, D.E.; Guzmán-Mejía, F.; Godínez-Victoria, M.; Drago-Serrano, M.E. Role of Stress on Driving the Intestinal Paracellular Permeability. Curr. Issues Mol. Biol. 2023, 45, 9284-9305. https://doi.org/10.3390/cimb45110581
Molotla-Torres DE, Guzmán-Mejía F, Godínez-Victoria M, Drago-Serrano ME. Role of Stress on Driving the Intestinal Paracellular Permeability. Current Issues in Molecular Biology. 2023; 45(11):9284-9305. https://doi.org/10.3390/cimb45110581
Chicago/Turabian StyleMolotla-Torres, Daniel Efrain, Fabiola Guzmán-Mejía, Marycarmen Godínez-Victoria, and Maria Elisa Drago-Serrano. 2023. "Role of Stress on Driving the Intestinal Paracellular Permeability" Current Issues in Molecular Biology 45, no. 11: 9284-9305. https://doi.org/10.3390/cimb45110581
APA StyleMolotla-Torres, D. E., Guzmán-Mejía, F., Godínez-Victoria, M., & Drago-Serrano, M. E. (2023). Role of Stress on Driving the Intestinal Paracellular Permeability. Current Issues in Molecular Biology, 45(11), 9284-9305. https://doi.org/10.3390/cimb45110581