
Overexpression of KvCHX Enhances Salt Tolerance in Arabidopsis thaliana Seedlings
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Treatments
2.2. Isolation and Sequence Analysis of KvCHX, Protein Structure, Phylogenetic Tree, and Multiple Alignment
2.3. Vector Construction and Transformation
2.4. Confirmation of Transgenic Arabidopsis Plant
2.5. Western Blot Analysis of Transgenic Arabidopsis Plant
2.6. Subcellular Localization Assay
2.7. Salt Stress Treatments Involving the Transgenic Arabidopsis Plant
2.8. K+ Deficient Treatments of Transgenic Arabidopsis Plant
2.9. Determination of Potassium and Sodium Ion Content
3. Results
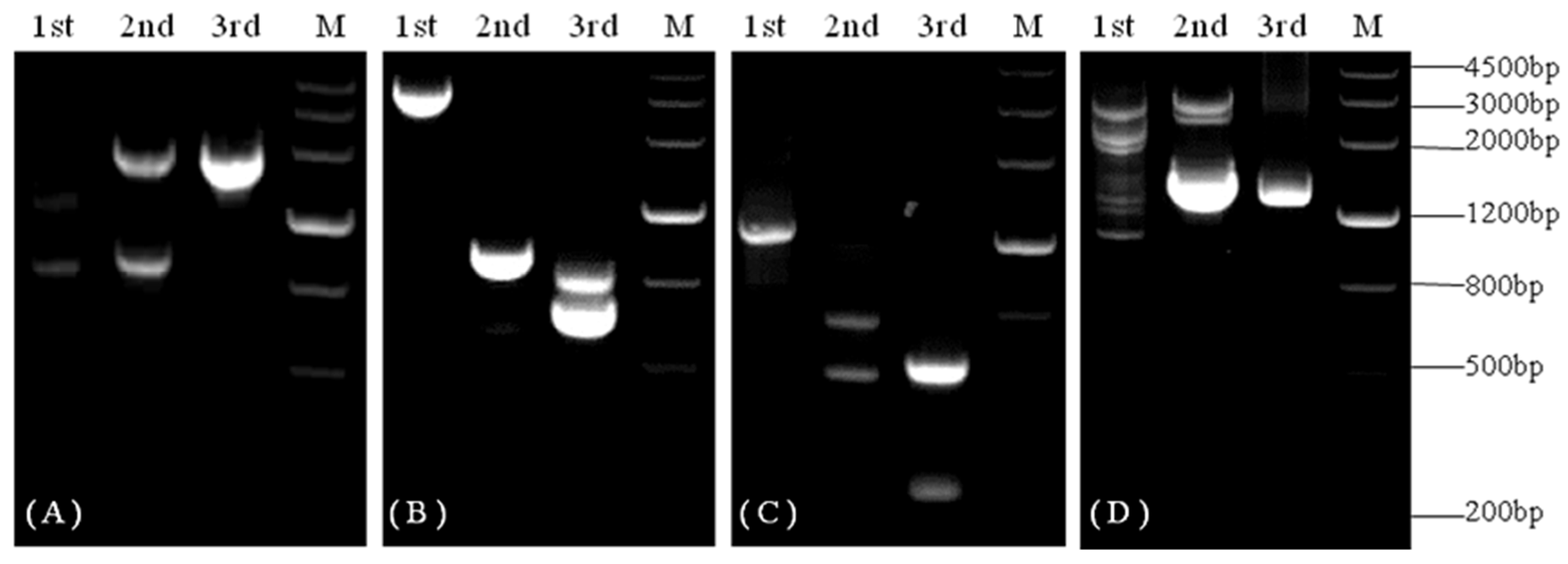
3.1. Isolation and Sequence Analysis of KvCHX
3.2. Analysis of KvCHX Gene Structure
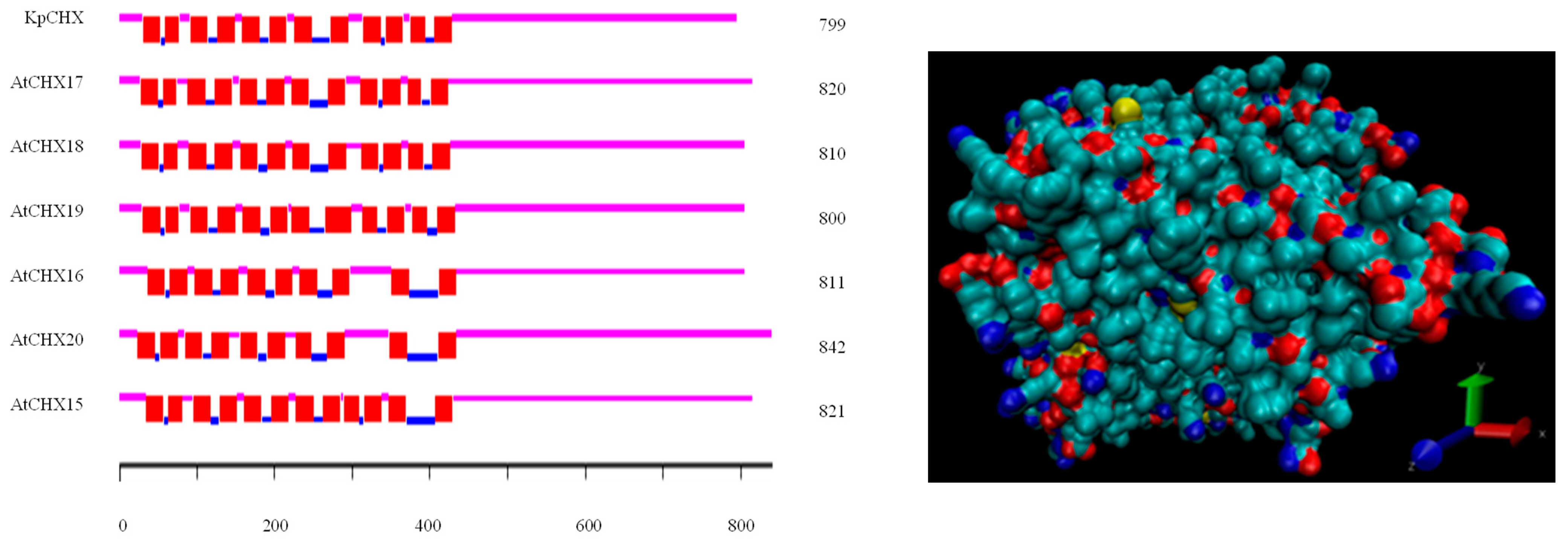
3.3. Structure and Function Analysis of KvCHX Protein
3.4. Phylogenetic Analysis and Multiple Alignment of CHXs
3.5. Expression and Subcellular Localization of Transgenic A. Thaliana
3.6. Overexpression of KvCHX Enhances Tolerance to Salt Stress in Transgenic Arabidopsis
3.7. KvCHX-Overexpressing Plants Grew Better Than WT under K+ Deficient Conditions
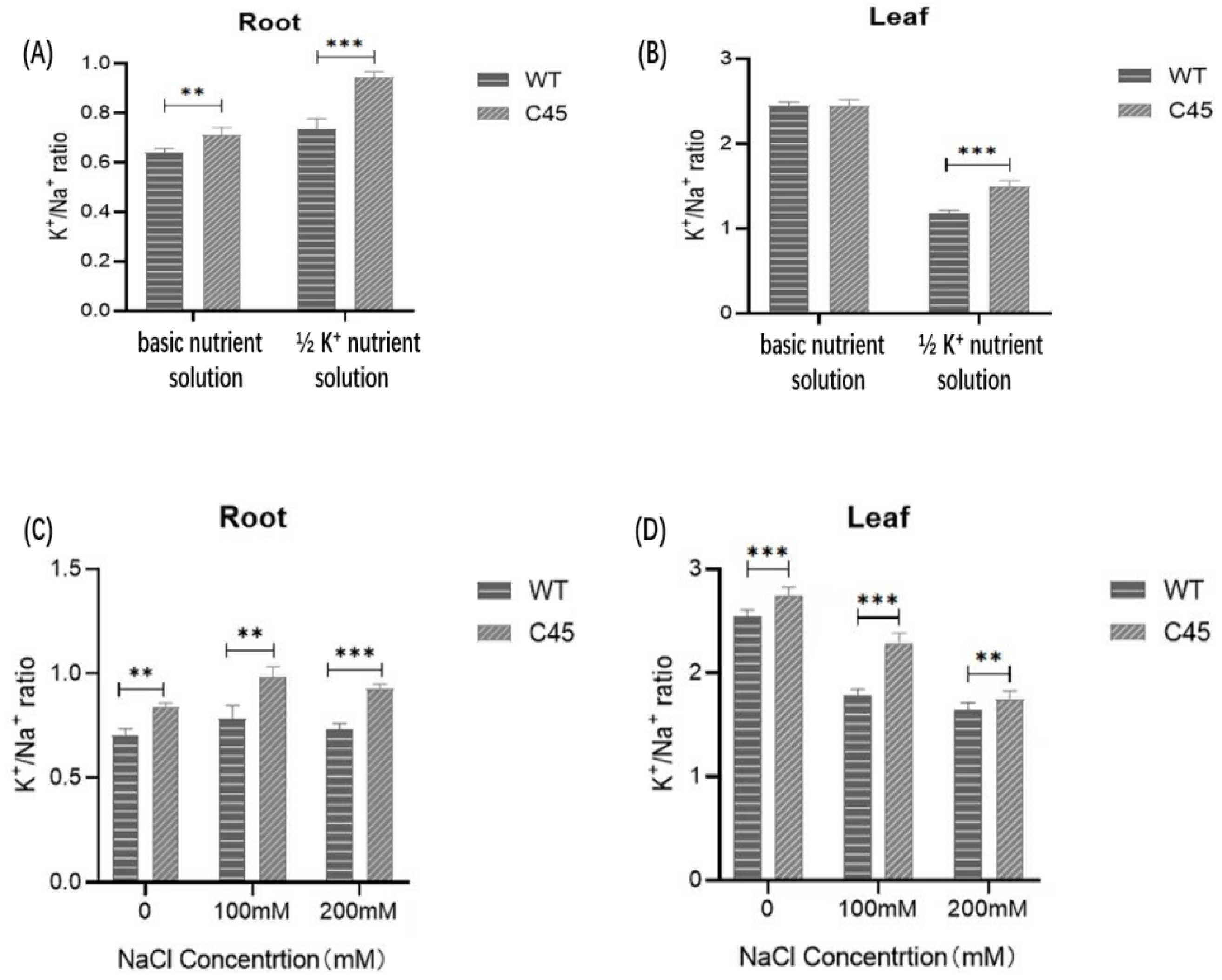
3.8. Determination of Ion Contents of Transgenic Arabidopsis Lines under Deficient K+ Treatment and Treatment with NaCl
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Leonardis, A.M.; Marone, D.; Mazzucotelli, E.; Neffar, F.; Rizza, F.; Di Fonzo, N.; Cattivelli, L.; Mastrangelo, A.M. Durum wheat genes up-regulated in the early phases of cold stress are modulated by drought in a developmental and genotype dependent manner. Plant Sci. 2007, 172, 1005–1016. [Google Scholar] [CrossRef]
- Yan, K.; Shao, H.; Shao, C.; Chen, P.; Zhao, S.; Brestic, M.; Chen, X. Physiological adaptive mechanisms of plants grown in saline soil and implications for sustainable saline agriculture in coastal zone. Acta Physiol. Plant. 2013, 35, 2867–2878. [Google Scholar] [CrossRef]
- Tang, X.; Mu, X.; Shao, H.; Wang, H.; Brestic, M. Global plant-responding mechanisms to salt stress: Physiological and molecular levels and implications in biotechnology. Crit. Rev. Biotechnol. 2015, 35, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- James, R.A.; Blake, C.; Byrt, C.S.; Munns, R. Major genes for Na+ exclusion, Nax1 and Nax2 (wheat HKT1;4 and HKT1;5), decrease Na+ accumulation in bread wheat leaves under saline and waterlogged conditions. J. Exp. Bot. 2011, 62, 2939–2947. [Google Scholar] [CrossRef]
- Szczerba, M.W.; Britto, D.T.; Kronzucker, H.J. K+ transport in plants: Physiology and molecular biology. J. Plant Physiol. 2009, 166, 447–466. [Google Scholar] [CrossRef]
- Elumalai, R.P.; Nagpal, P.; Reed, J.W. A mutation in the Arabidopsis KT2/KUP2 potassium transporter gene affects shoot cell expansion. Plant Cell 2002, 14, 119–131. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Sanders, D. Mechanisms of potassium absorption by higher plant roots. Physiol. Plant. 1996, 96, 158–168. [Google Scholar] [CrossRef]
- Philippar, K.; Fuchs, I.; Luthen, H.; Hoth, S.; Bauer, C.S.; Haga, K.; Thiel, G.; Ljung, K.; Sandberg, G.; Bottger, M.; et al. Auxin-induced K+ channel expression represents an essential step in coleoptile growth and gravitropism. Proc. Natl. Acad. Sci. USA 1999, 96, 12186–12191. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, F.; Amtmann, A. K+ Nutrition and Na+ Toxicity: The Basis of Cellular K+/Na+ Ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef]
- Sze, H.; Chanroj, S. Plant Endomembrane Dynamics: Studies of K+/H+ Antiporters Provide Insights on the Effects of pH and Ion Homeostasis. Plant Physiol. 2018, 177, 875–895. [Google Scholar] [CrossRef] [PubMed]
- Padmanaban, S.; Chanroj, S.; Kwak, J.M.; Li, X.; Ward, J.M.; Sze, H. Participation of endomembrane cation/H+ exchanger AtCHX20 in osmoregulation of guard cells. Plant Physiol. 2007, 144, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Guan, R.; Bose, J.; Henderson, S.W.; Wege, S.; Qiu, L.; Gilliham, M. Soybean CHX-type ion transport protein GmSALT3 confers leaf Na+ exclusion via a root derived mechanism, and Cl− exclusion via a shoot derived process. Plant Cell Environ. 2021, 44, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Xia, Y.; Ma, B.L.; Feng, C.; Qin, P. Culture of Seashore Mallow under Different Salinity Levels Using Plastic Nutrient-Rich Matrices and Transplantation. Agron. J. 2010, 102, 395–402. [Google Scholar] [CrossRef]
- Godfrey, R.K.; Wooten, J.W. Aquatic and Wetland Plants of Southeastern United States. Vol. II: Dicotyledons; University of Georgia Press: Athens, Greece, 1981. [Google Scholar]
- Islam, M.N.; Wilson, C.A.; Watkins, T.R. Nutritional evaluation of seashore mallow seed, Kosteletzkya virginica. J. Agric. Food Chem. 1982, 30, 1195–1198. [Google Scholar] [CrossRef]
- Moser, B.R.; Dien, B.S.; Seliskar, D.M.; Gallagher, J.L. Seashore mallow (Kosteletzkya pentacarpos) as a salt-tolerant feedstock for production of biodiesel and ethanol. Renew. Energy 2013, 50, 833–839. [Google Scholar] [CrossRef]
- Wang, H.; Tang, X.; Wang, H.; Shao, H. Physiological responses of Kosteletzkya virginica to coastal wetland soil. Sci. World J. 2015, 2015, 354581. [Google Scholar] [CrossRef]
- Guo, Y.; Tian, Z.; Qin, G.; Yan, D.; Zhang, J.; Zhou, W.; Qin, P. Gene expression of halophyte Kosteletzkya virginica seedlings under salt stress at early stage. Genetica 2009, 137, 189–199. [Google Scholar] [CrossRef]
- Sze, H.; Padmanaban, S.; Cellier, F.; Honys, D.; Cheng, N.; Bock, K.W.; Conéjéro, G.; Li, X.; Twell, D.; Ward, J.M.; et al. Expression patterns of a novel AtCHX gene family highlight potential roles in osmotic adjustment and K+ homeostasis in pollen development. Plant Physiol. 2004, 136, 2532–2547. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.; Ramírez, O.; Saldaña, C.; Coria, R.; Peña, A. A Saccharomyces cerevisiae mutant lacking a K+/H+ exchanger. J. Bacteriol. 1998, 180, 5860–5865. [Google Scholar] [CrossRef] [PubMed]
- Cellier, F.; Conéjéro, G.; Ricaud, L.; Luu, D.T.; Lepetit, M.; Gosti, F.; Casse, F. Characterization of AtCHX17, a member of the cation/H+ exchangers, CHX family, from Arabidopsis thaliana suggests a role in K+ homeostasis. Plant J. 2004, 39, 834–846. [Google Scholar] [CrossRef] [PubMed]
- Boyes, D.C.; Zayed, A.M.; Ascenzi, R.; McCaskill, A.J.; Hoffman, N.E.; Davis, K.R.; Gorlach, J. Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell 2001, 13, 1499–1510. [Google Scholar] [CrossRef]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.D.; Grossniklaus, U. A gateway cloning vector set for high-throughput functional analysis of genes in planta. Plant Physiol. 2003, 133, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Hofgen, R.; Willmitzer, L. Storage of competent cells for Agrobacterium transformation. Nucleic Acids Res. 1988, 16, 9877. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Chen, S.; Tian, Z.Y.; Guo, Y.Q. Characterization of hexokinase gene family members in Glycine max and functional analysis of GmHXK2 under salt stress. Front. Genet. 2023, 23, 1135290. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Song, S.; Chen, Y.; Chen, J.; Dai, X.; Zhang, W. Physiological mechanisms underlying OsNAC5-dependent tolerance of rice plants to abiotic stress. Planta 2011, 234, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Szabolcs, I. Salt-Affected Soils; CRC Press Inc.: Boca Raton, FL, USA, 1989. [Google Scholar]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chen, S.; Zhao, Q.; Wang, T.; Yang, C.; Diaz, C.; Sun, G.; Dai, S. Physiological and proteomic analysis of salinity tolerance in Puccinellia tenuiflora. J. Proteome Res. 2011, 10, 3852–3870. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Das, P.; Parida, A.K.; Agarwal, P.K. Proteomics, metabolomics, and ionomics perspectives of salinity tolerance in halophytes Front. Plant Sci. 2015, 6, 537. [Google Scholar]
- Ye, C.; Yang, X.; Xia, X.; Yin, W. Comparative analysis of cation/proton antiporter superfamily in plants. Gene 2013, 521, 245–251. [Google Scholar] [CrossRef]
- Brett, C.L.; Donowitz, M.; Rao, R. Evolutionary origins of eukaryotic sodium/proton exchangers. Am. J. Physiol. Cell Physiol. 2005, 288, C223–C239. [Google Scholar] [CrossRef]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J.; et al. The Pfam protein families database. Nucleic Acids Res. 2012, 40, D290–D301. [Google Scholar] [CrossRef]
- Zhu, J. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Bose, J.; Rodrigo-Moreno, A.; Lai, D.; Xie, Y.; Shen, W.; Shabala, S. Rapid regulation of the plasma membrane H⁺-ATPase activity is essential to salinity tolerance in two halophyte species, Atriplex lentiformis and Chenopodium quinoa. Ann. Bot. 2015, 115, 481–494. [Google Scholar] [CrossRef]
- Zhao, J.; Cheng, N.; Motes, C.M.; Blancaflor, E.B.; Moore, M.; Gonzales, N.; Padmanaban, S.; Sze, H.; Ward, J.M.; Hirschi, K.D. AtCHX13 Is a Plasma Membrane K+ Transporter. Plant Physiol. 2008, 148, 796–807. [Google Scholar] [CrossRef] [PubMed]
- Chanroj, S.; Lu, Y.; Padmanaban, S.; Nanatani, K.; Uozumi, N.; Rao, R.; Sze, H. Plant-specific cation/H+ exchanger 17 and its homologs are endomembrane K+ transporters with roles in protein sorting. J. Biol. Chem. 2011, 286, 33931–33941. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.; Evans, A.R.; Newbury, H.J.; Pritchard, J. Functional analysis of CHX, 21: A putative sodium transporter in Arabidopsis. J. Exp. Bot. 2006, 57, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Guan, R.; Yu, L.; Berkowitz, O.; David, R.; Whelan, J.; Ford, M.; Wege, S.; Qiu, L.; Gilliham, M. Enhanced reactive oxygen detoxification occurs in salt-stressed soybean roots expressing GmSALT3. Physiol. Plant 2022, 174, e13709. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Li, M.W.; Zheng, C.; Xu, Y.; Sun, S.; Li, Z.; Wong, F.L.; Song, J.; Lin, W.; Li, Q.; et al. The soybean plasma membrane-localized cation/H+ exchanger GmCHX20a plays a negative role under salt stress. Physiol. Plant 2021, 171, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Sun, M.; DuanMu, H.; Ding, X.; Liu, B.; Zhu, Y.; Sun, X. GsCHX19.3, a member of cation/H+ exchanger superfamily from wild soybean contributes to high salinity and carbonate alkaline tolerance. Sci. Rep. 2017, 7, 9423. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, J.; Miller, A.J.; Luo, B.; Wang, M.; Zhu, Z.; Ouwerkerk, P.B.F. OsCHX14 is Involved in the K+ Homeostasis in Rice (Oryza sativa) Flowers. Plant. Cell Physiol. 2016, 57, 1530–1543. [Google Scholar] [CrossRef]
- Hamada, A.; Shono, M.; Xia, T.; Ohta, M.; Hayashi, Y.; Tanaka, A.; Hayakawa, T. Isolation and characterization of a Na+/H+ antiporter gene from the halophyte Atriplex gmelini. Plant Mol. Biol. 2001, 46, 35–42. [Google Scholar] [CrossRef]
- Shi, H.; Lee, B.H.; Wu, S.J.; Zhu, J. Overexpression of a plasma membrane Na+/H+ antiporter gene improves salt tolerance in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 81–85. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GSP1-5′-2 | 5′-CCCAAAACTTCAGGACCAGG-3′ | GSP1-3′-2 | 5′-CCTGGTCCTGAAGTTTTGGG-3′ |
| GSP1-5′-3 | 5′-GCTTCGTGATTGTCGTGTCC-3′ | GSP1-3′-3 | 5′-CAATCTTTTCGTGGTGGGCC-3′ |
| GSP2-5′-1 | 5′-GGGGAGAAGAGTTTGTTCCTG-3′ | GSP3-5′-1 | 5′-CAACTATCTCGCCGATGACC-3′ |
| GSP2-5′-2 | 5′-TACGGGCTAACACAGGGAAG-3′ | GSP3-5′-2 | 5′-GCGAGGATACGAGTGAGTGC-3′ |
| GSP2-5′-3 | 5′-AAGGTGTCCAGCACCGTTAG-3′ | GSP3-5′-3 | 5′-GTACCGCTATCACCGACGAA-3′ |
| RV-M | 5′-GAGCGGATAACAATTTCACACAGG-3′ | M13-47 | 5′-CGCCAGGGTTTTCCCAGTCACGAC-3′ |
| PCR Reaction | Cycle No. | Thermal Condition |
|---|---|---|
| The first round of PCR(GSP-5’-1/AP) | 1 | 94 °C 1min, 98 °C 1min; |
| 5 | 94 °C (30 s), 60 °C (1 min), 72 °C (3 min) | |
| 1 | 94 °C (30 s), 25 °C (3 min), 72 °C (3 min) | |
| 15 | 94 °C (30 s), 60 °C (1 min), 72 °C (3 min); 94 °C (30 s), 60 °C (1 min), 72 °C (3 min); 94 °C(30 s), 44 °C (1 min), 72 °C (3 min) | |
| 1 | 72 °C (10 min) | |
| The second round of PCR(GSP-5′-2/AP) | 15 | 94 °C (30 s), 60 °C (1 min), 72 °C (3 min); 94 °C (30 s), 60 °C (1 min), 72 °C (3 min); 94 °C (30 s), 44 °C (1 min), 72 °C (3 min) |
| 1 | 72 °C (10min) | |
| The third round of PCR(GSP-5′-3/AP) | 15 | 94 °C (30 s), 60 °C (1 min), 72 °C (3 min); 94 °C (30 s), 60 °C (1 min), 72 °C (3 min); 94 °C (30 s), 44 °C (1 min),72 °C (3 min) |
| 1 | 72 °C, 10 min |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Zhu, C.; Tian, Z. Overexpression of KvCHX Enhances Salt Tolerance in Arabidopsis thaliana Seedlings. Curr. Issues Mol. Biol. 2023, 45, 9692-9708. https://doi.org/10.3390/cimb45120605
Guo Y, Zhu C, Tian Z. Overexpression of KvCHX Enhances Salt Tolerance in Arabidopsis thaliana Seedlings. Current Issues in Molecular Biology. 2023; 45(12):9692-9708. https://doi.org/10.3390/cimb45120605
Chicago/Turabian StyleGuo, Yuqi, Chengrong Zhu, and Zengyuan Tian. 2023. "Overexpression of KvCHX Enhances Salt Tolerance in Arabidopsis thaliana Seedlings" Current Issues in Molecular Biology 45, no. 12: 9692-9708. https://doi.org/10.3390/cimb45120605
APA StyleGuo, Y., Zhu, C., & Tian, Z. (2023). Overexpression of KvCHX Enhances Salt Tolerance in Arabidopsis thaliana Seedlings. Current Issues in Molecular Biology, 45(12), 9692-9708. https://doi.org/10.3390/cimb45120605