
Peroxiredoxins and Hypoxia-Inducible Factor-1α in Duodenal Tissue: Emerging Factors in the Pathophysiology of Pediatric Celiac Disease Patients

Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Sample Preparation
2.2. RNA Isolation, cDNA Preparation, and Real-Time PCR Analysis
2.3. Protein Extraction and Western Blotting
2.4. Statistical Analysis
3. Results
3.1. tTG and Inflammatory Proteins Expressions in Duodenal Biopsy Samples of CD Patients
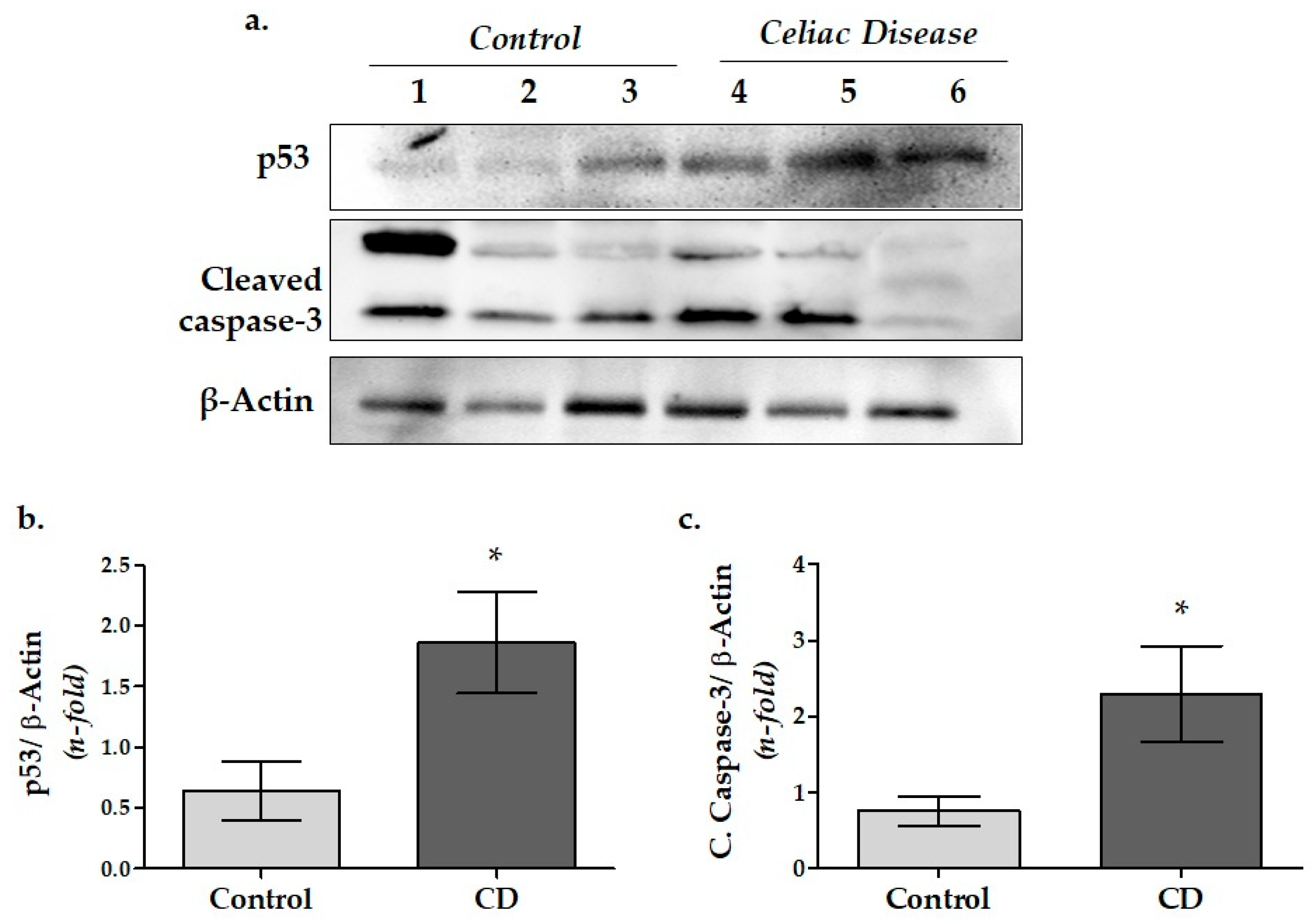
3.2. Apoptosis-Related Protein Expressions in Duodenal Biopsy Samples of CD Patients
3.3. HIF-1α Protein and mRNA Expressions in Duodenal Biopsy Samples of CD Patients
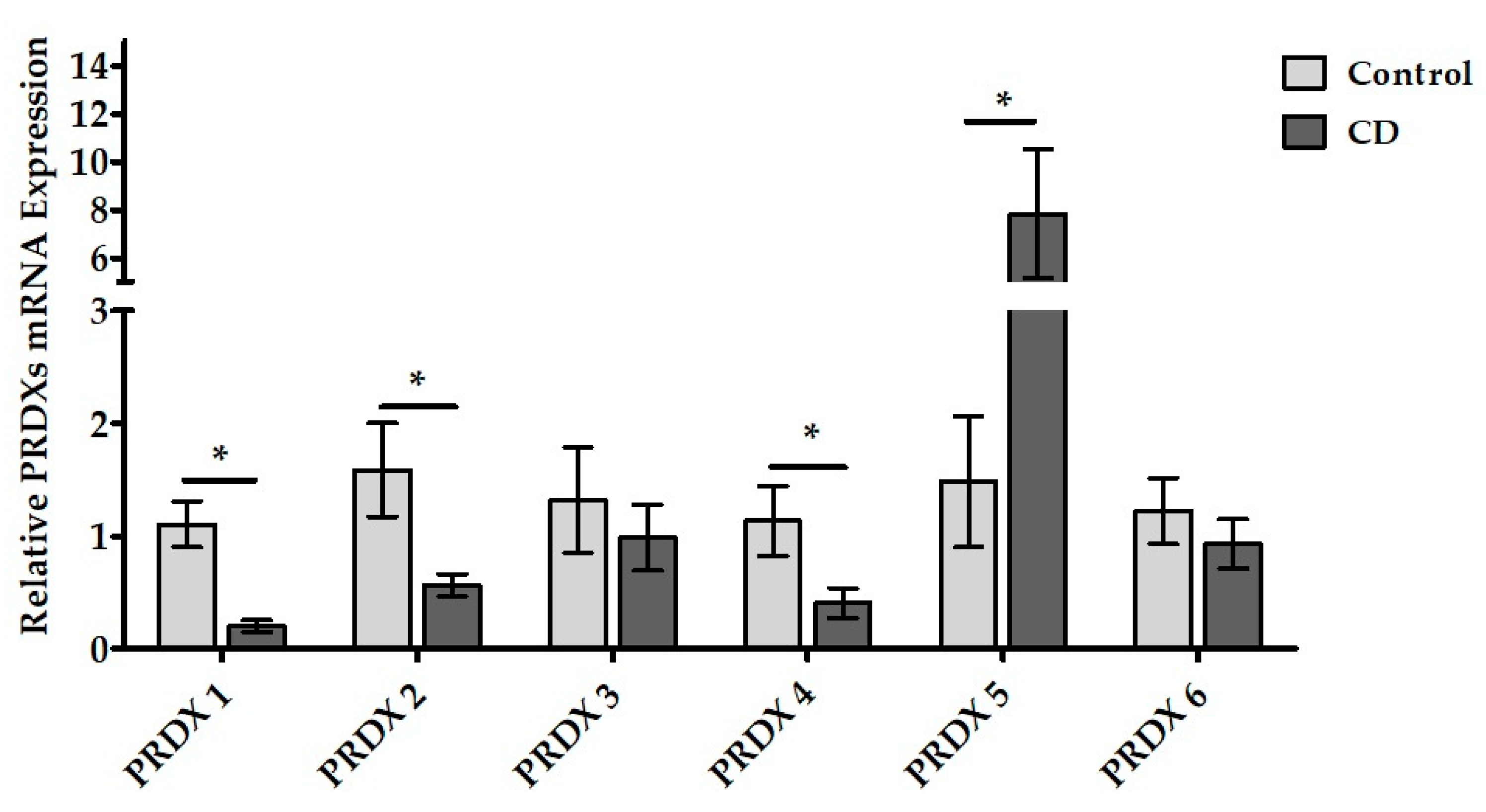
3.4. PRDXs mRNA Expressions in Duodenal Samples of CD Patients
3.5. PRDXs Protein Expressions in Duodenal Samples of CD Patients
4. Discussion
4.1. tTG and Inflammatory Proteins Expression in Duodenal Tissue of Pediatric CD Patients
4.2. Apoptosis-Related Caspase-3 and p53 Protein Expressions in Duodenal Tissue of Pediatric CD Patients
4.3. Hypoxia-Inducible Factor (HIF-1α) mRNA and Protein Expressions in Duodenal Tissue of Pediatric CD Patients
4.4. Peroxiredoxins mRNA and Protein Expressions in Duodenal Tissue of Pediatric CD Patients
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sahin, Y. Celiac Disease in Children: A Review of the Literature. World J. Clin. Pediatr. 2021, 10, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Vaquero, L.; Bernardo, D.; León, F.; Rodríguez-Martín, L.; Alvarez-Cuenllas, B.; Vivas, S. Challenges to Drug Discovery for Celiac Disease and Approaches to Overcome Them. Expert Opin. Drug Discov. 2019, 14, 957–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindfors, K.; Ciacci, C.; Kurppa, K.; Lundin, K.E.A.; Makharia, G.K.; Mearin, M.L.; Murray, J.A.; Verdu, E.F.; Kaukinen, K. Coeliac Disease. Nat. Rev. Dis. Primers 2019, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Arora, A.; Strand, T.A.; Leffler, D.A.; Catassi, C.; Green, P.H.; Kelly, C.P.; Ahuja, V.; Makharia, G.K. Global Prevalence of Celiac Disease: Systematic Review and Meta-Analysis. Clin. Gastroenterol. Hepatol. 2018, 16, 823–836.e2. [Google Scholar] [CrossRef] [Green Version]
- Kupfer, S.S.; Jabri, B. Pathophysiology of Celiac Disease. Gastrointest Endosc. Clin. N. Am. 2012, 22, 639–660. [Google Scholar] [CrossRef] [Green Version]
- Denham, J.M.; Hill, I.D. Celiac Disease and Autoimmunity: Review and Controversies. Curr. Allergy Asthma Rep. 2013, 13, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Ilus, T.; Kaukinen, K.; Virta, L.J.; Pukkala, E.; Collin, P. Incidence of Malignancies in Diagnosed Celiac Patients: A Population-Based Estimate. Am. J. Gastroenterol. 2014, 109, 1471–1477. [Google Scholar] [CrossRef]
- Simula, M.P.; Cannizzaro, R.; Canzonieri, V.; Pavan, A.; Maiero, S.; Toffoli, G.; de Re, V. PPAR Signaling Pathway and Cancer-Related Proteins Are Involved in Celiac Disease-Associated Tissue Damage. Mol. Med. 2010, 16, 199–209. [Google Scholar] [CrossRef]
- Sziksz, E.; Vörös, P.; Veres, G.; Fekete, A.; Vannay, Á. Coeliac Disease: From Triggering Factors to Treatment. Int. J. Celiac Dis. 2016, 1, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, G.; Bacchetti, T.; Masciangelo, S.; Saturni, L. Celiac Disease, Inflammation and Oxidative Damage: A Nutrigenetic Approach. Nutrients 2012, 4, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Barone, M.V.; Troncone, R.; Auricchio, S. Gliadin Peptides as Triggers of the Proliferative and Stress/Innate Immune Response of the Celiac Small Intestinal Mucosa. Int. J. Mol. Sci. 2014, 15, 20518–20537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abadie, V.; Jabri, B. IL-15: A Central Regulator of Celiac Disease Immunopathology. Immunol. Rev. 2014, 260, 221–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Re, V.; Caggiari, L.; Tabuso, M.; Cannizzaro, R. The Versatile Role of Gliadin Peptides in Celiac Disease. Clin. Biochem. 2013, 46, 552–560. [Google Scholar] [CrossRef]
- Palová-Jelínková, L.; Dáňová, K.; Drašarová, H.; Dvořák, M.; Funda, D.P.; Fundová, P.; Kotrbová-Kozak, A.; Černá, M.; Kamanová, J.; Martin, S.F.; et al. Pepsin Digest of Wheat Gliadin Fraction Increases Production of IL-1β via TLR4/MyD88/TRIF/MAPK/NF-ΚB Signaling Pathway and an NLRP3 Inflammasome Activation. PLoS ONE 2013, 8, e62426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, M.; Luo, Z.; Zhang, W.; Zhu, D.; Lu, Y.; Wu, J.; Yao, X. Protective Effect of Curcumin against Irinotecan-Induced Intestinal Mucosal Injury via Attenuation of NF-ΚB Activation, Oxidative Stress and Endoplasmic Reticulum Stress. Int. J. Oncol. 2019, 54, 1376–1386. [Google Scholar] [CrossRef]
- Holmgren Peterson, K.K. Fälth-Magn Children with Celiac Disease Express Inducible Nitric Oxide Synthase in the Small Intestine during Gluten Challenge. Scand. J. Gastroenterol. 1998, 33, 939–943. [Google Scholar] [CrossRef]
- Korbecki, J.; Simińska, D.; Gąssowska-Dobrowolska, M.; Listos, J.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Chronic and Cycling Hypoxia: Drivers of Cancer Chronic Inflammation through Hif-1 and Nf-Κb Activation. A Review of the Molecular Mechanisms. Int. J. Mol. Sci. 2021, 22, 10701. [Google Scholar] [CrossRef]
- Perez, F.; Ruera, C.N.; Miculan, E.; Carasi, P.; Chirdo, F.G. Programmed Cell Death in the Small Intestine: Implications for the Pathogenesis of Celiac Disease. Int. J. Mol. Sci. 2021, 22, 7426. [Google Scholar] [CrossRef]
- Piatek-Guziewicz, A.; Ptak-Belowska, A.; Przybylska-Felus, M.; Pasko, P.; Zagrodzki, P.; Brzozowski, T.; Mach, T.; Zwolinska-Wcislo, M. Intestinal Parameters of Oxidative Imbalance in Celiac Adults with Extraintestinal Manifestations. World J. Gastroenterol. 2017, 23, 7849–7862. [Google Scholar] [CrossRef]
- Lee, M.; Betman, S.; Iuga, A.; Yang, H.M.; Fleming, J.; Green, P.H.R.; Lebwohl, B.; Lagana, S.M. An Association between Crypt Apoptotic Bodies and Mucosal Flattening in Celiac Disease Patients Exposed to Dietary Gluten. Diagn. Pathol. 2019, 14, 98. [Google Scholar] [CrossRef] [Green Version]
- Szaflarska-Poplawska, A.; Siomek, A.; Czerwionka-Szaflarska, M.; Gackowski, D.; Rózalski, R.; Guz, J.; Szpila, A.; Zarakowska, E.; Olinski, R. Oxidatively Damaged DNA/Oxidative Stress in Children with Celiac Disease. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1960–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, L.B.; Hall, A.; Nelson, K.J. Overview of Peroxiredoxins in Oxidant Defense and Redox Regulation. In Current Protocols in Toxicology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 1–20. [Google Scholar]
- Luciani, A.; Villella, V.R.; Vasaturo, A.; Giardino, I.; Pettoello-Mantovani, M.; Guido, S.; Cexus, O.N.; Peake, N.; Londei, M.; Quaratino, S.; et al. Lysosomal Accumulation of Gliadin P31-43 Peptide Induces Oxidative Stress and Tissue Transglutaminase-Mediated PPAR Downregulation in Intestinal Epithelial Cells and Coeliac Mucosa. Gut 2010, 59, 311–319. [Google Scholar] [CrossRef]
- Forshaw, T.E.; Reisz, J.A.; Nelson, K.J.; Gumpena, R.; Lawson, J.R.; Jönsson, T.J.; Wu, H.; Clodfelter, J.E.; Johnson, L.C.; Furdui, C.M.; et al. Specificity of Human Sulfiredoxin for Reductant and Peroxiredoxin Oligomeric State. Antioxidants 2021, 10, 946. [Google Scholar] [CrossRef]
- Rhee, S.G.; Chae, H.Z.; Kim, K. Peroxiredoxins: A Historical Overview and Speculative Preview of Novel Mechanisms and Emerging Concepts in Cell Signaling. Free Radic Biol. Med. 2005, 38, 1543–1552. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Woo, H.A.; Kil, I.S.; Bae, S.H. Peroxiredoxin Functions as a Peroxidase and a Regulator and Sensor of Local Peroxides. J. Biol. Chem. 2012, 287, 4403–4410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.G.; Kang, S.W.; Jeong, W.; Chang, T.-S.; Yang, K.-S.; Woo, H.A. Intracellular Messenger Function of Hydrogen Peroxide and Its Regulation by Peroxiredoxins. Curr. Opin. Cell Biol. 2005, 17, 183–189. [Google Scholar] [CrossRef]
- Ishii, T.; Itoh, K.; Akasaka, J.; Yanagawa, T.; Takahashi, S.; Yoshida, H.; Bannai, S.; Yamamoto, M. Induction of Murine Intestinal and Hepatic Peroxiredoxin MSP23 by Dietary Butylated Hydroxyanisole. Carcinogenesis 2000, 21, 1013–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, T.; Itoh, K.; Takahashi, S.; Sato, H.; Yanagawa, T.; Katoh, Y.; Bannai, S.; Yamamoto, M. Transcription Factor Nrf2 Coordinately Regulates a Group of Oxidative Stress-Inducible Genes in Macrophages. J. Biol. Chem. 2000, 275, 16023–16029. [Google Scholar] [CrossRef] [Green Version]
- Perkins, A.; Poole, L.B.; Karplus, P.A. Tuning of Peroxiredoxin Catalysis for Various Physiological Roles. Biochemistry 2014, 53, 7693–7705. [Google Scholar] [CrossRef] [Green Version]
- Knoops, B.; Argyropoulou, V.; Becker, S.; Ferté, L.; Kuznetsova, O. Multiple Roles of Peroxiredoxins in Inflammation. Mol. Cells 2016, 39, 60–64. [Google Scholar]
- Wu, M.; Deng, C.; Lo, T.H.; Chan, K.Y.; Li, X.; Wong, C.M. Peroxiredoxin, Senescence, and Cancer. Cells 2022, 11, 1772. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Du, X.; Peng, Z.; Zhang, Z.; Cui, L.; Li, D.; Wang, R.; Ma, M. Silencing of Peroxiredoxin 1 Expression Ameliorates Ulcerative Colitis in a Rat Model. J. Int. Med. Res. 2021, 49, 300060520986313. [Google Scholar] [CrossRef]
- Iizuka, M.; Nakagomi, O.; Nanjo, H.; Chiba, M.; Fukushima, T.; Sugita, A.; Sagara, S.; Horie, Y.; Watanabe, S. Molecular Cloning Reveals Nearly Half of Patients with Crohn’s Disease Have an Antibody to Peroxiredoxin 6-like Protein. J. Gastroenterol. Hepatol. 2012, 27, 1388–1394. [Google Scholar] [CrossRef]
- Dehne, N.; Brüne, B. HIF-1 in the Inflammatory Microenvironment. Exp. Cell Res. 2009, 315, 1791–1797. [Google Scholar] [CrossRef]
- Vörös, P.; Sziksz, E.; Himer, L.; Ónody, A.; Pap, D.; Frivolt, K.; Szebeni, B.; Lippai, R.; Győrffy, H.; Fekete, A.; et al. Expression of PARK7 Is Increased in Celiac Disease. Virchows Archiv. 2013, 463, 401–408. [Google Scholar] [CrossRef]
- Hirota, S.; Beck, P.; MacDonald, J. Targeting Hypoxia-Inducible Factor-1 (HIF-1) Signaling in Therapeutics: Implications for the Treatment of Inflammatory Bowel Disease. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karhausen, J.; Furuta, G.T.; Tomaszewski, J.E.; Johnson, R.S.; Colgan, S.P.; Haase, V.H. Epithelial Hypoxia-Inducible Factor-1 Is Protective in Murine Experimental Colitis. J. Clin. Investig. 2004, 114, 1098–1106. [Google Scholar] [CrossRef]
- Vannay, A.; Sziksz, E.; Prókai, A.; Veres, G.; Molnár, K.; Szakál, D.N.; Onódy, A.; Korponay-Szabó, I.R.; Szabó, A.; Tulassay, T.; et al. Increased Expression of Hypoxia-Inducible Factor 1alpha in Coeliac Disease. Pediatr. Res. 2010, 68, 118–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddell, J.R.; Maier, P.; Sass, S.N.; Moser, M.T.; Foster, B.A.; Gollnick, S.O. Peroxiredoxin 1 Stimulates Endothelial Cell Expression of VEGF via TLR4 Dependent Activation of HIF-1α. PLoS ONE 2012, 7, e50394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabharwal, S.S.; Waypa, G.B.; Marks, J.D.; Schumacker, P.T. Peroxiredoxin-5 Targeted to the Mitochondrial Intermembrane Space Attenuates Hypoxia-Induced Reactive Oxygen Species Signalling. Biochem. J. 2013, 456, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Gordeeva, A.E.; Temnov, A.A.; Charnagalov, A.A.; Sharapov, M.G.; Fesenko, E.E.; Novoselov, V.I. Protective Effect of Peroxiredoxin 6 in Ischemia/Reperfusion-Induced Damage of Small Intestine. Dig Dis. Sci. 2015, 60, 3610–3619. [Google Scholar] [CrossRef] [PubMed]
- Xi, H.; Gao, Y.-H.; Han, D.-Y.; Li, Q.-Y.; Feng, L.-J.; Zhang, W.; Ji, G.; Xiao, J.-C.; Zhang, H.-Z.; Wei, Q. Hypoxia Inducible Factor-1α Suppresses Peroxiredoxin 3 Expression to Promote Proliferation of CCRCC Cells. FEBS Lett. 2014, 588, 3390–3394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revised Criteria for Diagnosis of Coeliac Disease. Report of Working Group of European Society of Paediatric Gastroenterology and Nutrition. Arch. Dis. Child 1990, 65, 909–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Armagan, G.; Sevgili, E.; Gürkan, F.T.; Köse, F.A.; Bilgiç, T.; Dagci, T.; Saso, L. Regulation of the Nrf2 Pathway by Glycogen Synthase Kinase-3β in MPP+-Induced Cell Damage. Molecules 2019, 24, 1377. [Google Scholar] [CrossRef] [Green Version]
- Caputo, I.; Secondo, A.; Lepretti, M.; Paolella, G.; Auricchio, S.; Barone, M.V.; Esposito, C. Gliadin Peptides Induce Tissue Transglutaminase Activation and ER-Stress through Ca2+ Mobilization in Caco-2 Cells. PLoS ONE 2012, 7, e45209. [Google Scholar] [CrossRef]
- Antonella Nadalutti, C.; Korponay-Szabo, I.R.; Kaukinen, K.; Wang, Z.; Griffin, M.; Mäki, M.; Lindfors, K. Thioredoxin Is Involved in Endothelial Cell Extracellular Transglutaminase 2 Activation Mediated by Celiac Disease Patient IgA. PLoS ONE 2013, 8, e77277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, I.; Cavill, D.; Murray, I.A.; Long, R.G. Elevated Expression of INOS MRNA and Protein in Coeliac Disease. Clin. Chim. Acta 2005, 356, 134–142. [Google Scholar] [CrossRef]
- Yu, T.; Hu, S.; Min, F.; Li, J.; Shen, Y.; Yuan, J.; Gao, J.; Wu, Y.; Chen, H. Wheat Amylase Trypsin Inhibitors Aggravate Intestinal Inflammation Associated with Celiac Disease Mediated by Gliadin in BALB/c Mice. Foods 2022, 11, 1559. [Google Scholar] [CrossRef]
- Shalimar, D.M.; Das, P.; Sreenivas, V.; Gupta, S.D.; Panda, S.K.; Makharia, G.K. Mechanism of Villous Atrophy in Celiac Disease: Role of Apoptosis and Epithelial Regeneration. Arch. Pathol. Lab. Med. 2013, 137, 1262–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannini, C.; Sanchez, M.; Straface, E.; Scazzocchio, B.; Silano, M.; de Vincenzi, M. Induction of Apoptosis in Caco-2 Cells by Wheat Gliadin Peptides. Toxicology 2000, 145, 63–71. [Google Scholar] [CrossRef] [PubMed]
- McGettrick, A.F.; O’Neill, L.A.J. The Role of HIF in Immunity and Inflammation. Cell Metab. 2020, 32, 524–536. [Google Scholar] [CrossRef] [PubMed]
- Olson, N.; Hristova, M.; Heintz, N.H.; Lounsbury, K.M.; van der Vliet, A. Activation of Hypoxia-Inducible Factor-1 Protects Airway Epithelium against Oxidant-Induced Barrier Dysfunction. Am. J. Physiol. Lung Cell Mol. Physiol. 2011, 301, L993–L1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummins, E.P.; Taylor, C.T. Hypoxia and Inflammation. Biochem. (Lond.) 2017, 39, 34–36. [Google Scholar] [CrossRef]
- Yin, J.; Ren, Y.; Yang, K.; Wang, W.; Wang, T.; Xiao, W.; Yang, H. The Role of Hypoxia-Inducible Factor 1-Alpha in Inflammatory Bowel Disease. Cell Biol. Int. 2022, 46, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.-J.; Isaacs, J.S.; Lee, S.; Trepel, J.; Neckers, L. IL-1β Mediated Up-regulation of HIF-lα via an NFkB/COX-2 Pathway Identifies HIF-1 as a Critical Link between Inflammation and Oncogenesis. FASEB J. 2003, 17, 2115–2117. [Google Scholar] [CrossRef] [Green Version]
- Cramer, T.; Yamanishi, Y.; Clausen, B.E.; Förster, I.; Pawlinski, R.; Mackman, N.; Haase, V.H.; Jaenisch, R.; Corr, M.; Nizet, V.; et al. HIF-1α Is Essential for Myeloid Cell-Mediated Inflammation. Cell 2003, 112, 645–657. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, G.; Bacchetti, T.; Saturni, L.; Manzella, N.; Candelaresi, C.; Benedetti, A.; di Sario, A. Lipid Peroxidation and Paraoxonase-1 Activity in Celiac Disease. J. Lipids 2012, 2012, 587479. [Google Scholar] [CrossRef]
- Stojiljković, V.; Todorović, A.; Pejić, S.; Kasapović, J.; Saičić, Z.S.; Radlović, N.; Pajović, S.B. Antioxidant Status and Lipid Peroxidation in Small Intestinal Mucosa of Children with Celiac Disease. Clin. Biochem. 2009, 42, 1431–1437. [Google Scholar] [CrossRef]
- Murray, I.A.; Daniels, I.; Coupland, K.; Smith, J.A.; Long, R.G. Increased Activity and Expression of INOS in Human Duodenal Enterocytes from Patients with Celiac Disease. Am. J. Physiol. -Gastrointest. Liver Physiol. 2002, 283, G319–G326. [Google Scholar] [CrossRef] [Green Version]
- Salzano, S.; Checconi, P.; Hanschmann, E.-M.; Lillig, C.H.; Bowler, L.D.; Chan, P.; Vaudry, D.; Mengozzi, M.; Coppo, L.; Sacre, S.; et al. Linkage of Inflammation and Oxidative Stress via Release of Glutathionylated Peroxiredoxin-2, Which Acts as a Danger Signal. Proc. Natl. Acad. Sci. USA 2014, 111, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Shichita, T.; Sakaguchi, R.; Ito, M.; Kondo, T.; Yoshimura, A. Peroxiredoxin Triggers Cerebral Post-Ischemic Inflammation. Inflamm. Regen 2013, 33, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Thapa, P.; Ding, N.; Hao, Y.; Alshahrani, A.; Jiang, H.; Wei, Q. Essential Roles of Peroxiredoxin IV in Inflammation and Cancer. Molecules 2022, 27, 6513. [Google Scholar] [CrossRef]
- Kunze, A.; Zierath, D.; Tanzi, P.; Cain, K.; Becker, K. Peroxiredoxin 5 (PRX5) Is Correlated Inversely to Systemic Markers of Inflammation in Acute Stroke. Stroke 2014, 45, 608–610. [Google Scholar] [CrossRef] [Green Version]
- Mo, Y.; Feinstein, S.I.; Manevich, Y.; Zhang, Q.; Lu, L.; Ho, Y.-S.; Fisher, A.B. 1-Cys Peroxiredoxin Knock-out Mice Express MRNA but Not Protein for a Highly Related Intronless Gene. FEBS Lett. 2003, 555, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Naito, Y.; Takagi, T.; Okada, H.; Omatsu, T.; Mizushima, K.; Handa, O.; Kokura, S.; Ichikawa, H.; Fujiwake, H.; Yoshikawa, T. Identification of Inflammation-Related Proteins in a Murine Colitis Model by 2D Fluorescence Difference Gel Electrophoresis and Mass Spectrometry. J. Gastroenterol. Hepatol. 2010, 25, S144–S148. [Google Scholar] [CrossRef]
- Jiang, P.; Smith, B.; Qvist, N.; Nielsen, C.; Wan, J.M.-F.; Sit, W.-H.; Jensen, T.K.; Wang, H.; Sangild, P.T. Intestinal Proteome Changes during Infant Necrotizing Enterocolitis. Pediatr. Res. 2013, 73, 268–276. [Google Scholar] [CrossRef]
- Lu, W.; Fu, Z.; Wang, H.; Feng, J.; Wei, J.; Guo, J. Peroxiredoxin 2 Knockdown by RNA Interference Inhibits the Growth of Colorectal Cancer Cells by Downregulating Wnt/β-Catenin Signaling. Cancer Lett. 2014, 343, 190–199. [Google Scholar] [CrossRef]
- Zhang, S.; Fu, Z.; Wei, J.; Guo, J.; Liu, M.; Du, K. Peroxiredoxin 2 Is Involved in Vasculogenic Mimicry Formation by Targeting VEGFR2 Activation in Colorectal Cancer. Med. Oncol. 2015, 32, 414. [Google Scholar] [CrossRef]
- Lu, W.; Fu, Z.; Wang, H.; Feng, J.; Wei, J.; Guo, J. Peroxiredoxin 2 Is Upregulated in Colorectal Cancer and Contributes to Colorectal Cancer Cells’ Survival by Protecting Cells from Oxidative Stress. Mol. Cell Biochem. 2014, 387, 261–270. [Google Scholar] [CrossRef]
- Lim, J.Y. Thioredoxin and Thioredoxin-Interacting Protein as Prognostic Markers for Gastric Cancer Recurrence. World J. Gastroenterol. 2012, 18, 5581. [Google Scholar] [CrossRef]
- Nicolussi, A.; D’Inzeo, S.; Capalbo, C.; Giannini, G.; Coppa, A. The Role of Peroxiredoxins in Cancer. Mol. Clin. Oncol. 2017, 6, 139–153. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient No. | MARSH Classifications | Serum EMA-IgA 1 | Serum tTG 2 Levels |
|---|---|---|---|
| 1 | Type 3a | + | >200 IU |
| 2 | Type 3b | +++ | >200 IU |
| 3 | Type 3a | +++ | >200 IU |
| 4 | Type 2a | +++ | >200 IU |
| 5 | Type 3c | +++ | >200 IU |
| 6 | Type 3c | + | >200 IU |
| 7 | Type 3c | +++ | >200 IU |
| Gene | Forward Primer (5′ → 3′) | Reverse Primer (5′ → 3′) | Base Pair |
|---|---|---|---|
| PRDX-1 (NM_181697.3) | 5′tttggtatcagacccgaagc3′ | 5′agtgaactggaaggcctgaa3′ | 185 bp |
| PRDX-2 (NM_005809.6) | 5′agatcatcgcgttcagcaac 3′ | 5′atcctcagacaagcgtctgg3′ | 182 bp |
| PRDX-3 (NM_001302272.2) | 5′gtcgcagtctcagtggattc3′ | 5′aacagcacaccgtagtctcg3′ | 140 bp |
| PRDX-4 (NM_006406.2) | 5′aacagctgtgatcgatggag3′ | 5′tcaagtctgtcgccaaaagc3′ | 138 bp |
| PRDX-5 (NM_012094.5) | 5′caagaagggtgtgctgtttg3′ | 5′taacactcagacaggccacc | 134 bp |
| PRDX-6 (NM_004905.3) | 5′atgcctgtgacagctcgtgtg 3′ | 5′tcttcttcagggatggttgg3′ | 212 bp |
| HIF-1α (NM_001530.4) | 5′tttccttctcttctccgcgtg 3′ | 5′ggctgcatctcgagactttt3′ | 175 bp |
| GAPDH (NM_002046.7) | 5′agccacatcgctcagacac 3′ | 5′gcccaatacgaccaaatcc3′ | 65 bp |
| Antibody | Brand | Source | Dilution | Antibody | Brand | Source | Dilution |
|---|---|---|---|---|---|---|---|
| anti-PRDX 1 | Pierce (PA5-29830) | Rabbit | 1:3000 | anti-iNOS | Pierce (MA3-030) | Mouse | 1:250 |
| anti-PRDX 2 | Proteintech (60202-1-Ig) | Mouse | 1:5000 | anti-TNFα | Proteintech (60291-1-Ig) | Mouse | 1:100 |
| anti-PRDX 3 | Proteintech (10664-1-AP) | Rabbit | 1:2000 | anti-pNF-κB-p65 | Pierce (MA5-15160) | Rabbit | 1:500 |
| anti-PRDX 4 | Pierce (PA5-34853) | Mouse | 1:1000 | anti-HIF-1α | Pierce (MA1-16511) | Mouse | 1:500 |
| anti-PRDX 5 | Proteintech (17724-1-AP) | Rabbit | 1:1000 | anti-tTG | Proteintech (15100-1-AP) | Rabbit | 1:2000 |
| anti-PRDX 6 | Pierce (H00009588-M01) | Rabbit | 1:1000 | Anti-p53 | Pierce (MA5-12557) | Mouse | 1:100 |
| anti-HIF-1α | Novus Biological (MA1-16511) | Mouse | 1:500 | Anti-cleaved caspase 3 | Cell Signalling (9661) | Rabbit | 1:1000 |
| anti-β-Actin | Invitrogen (15G5A11/E2) | Mouse | 1:10,000 |
| Gene | Control | CD |
|---|---|---|
| PRDX-1 | 1.105 ± 0.21 | 0.203 ± 0.05 * |
| PRDX-2 | 1.587 ± 0.41 | 0.566 ± 0.1 * |
| PRDX-3 | 1.32 ± 0.47 | 0.987 ± 0.29 |
| PRDX-4 | 1.135 ± 0.31 | 0.407 ± 0.13 * |
| PRDX-5 | 1.484 ± 0.58 | 7.832 ± 2.68 * |
| PRDX-6 | 1.226 ± 0.29 | 0.93 ± 0.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Köse, F.A.; Pabuccuoglu, A.; Karakoyun, M.; Aydogdu, S. Peroxiredoxins and Hypoxia-Inducible Factor-1α in Duodenal Tissue: Emerging Factors in the Pathophysiology of Pediatric Celiac Disease Patients. Curr. Issues Mol. Biol. 2023, 45, 1779-1793. https://doi.org/10.3390/cimb45020114
Köse FA, Pabuccuoglu A, Karakoyun M, Aydogdu S. Peroxiredoxins and Hypoxia-Inducible Factor-1α in Duodenal Tissue: Emerging Factors in the Pathophysiology of Pediatric Celiac Disease Patients. Current Issues in Molecular Biology. 2023; 45(2):1779-1793. https://doi.org/10.3390/cimb45020114
Chicago/Turabian StyleKöse, Fadime Aydın, Aysun Pabuccuoglu, Miray Karakoyun, and Sema Aydogdu. 2023. "Peroxiredoxins and Hypoxia-Inducible Factor-1α in Duodenal Tissue: Emerging Factors in the Pathophysiology of Pediatric Celiac Disease Patients" Current Issues in Molecular Biology 45, no. 2: 1779-1793. https://doi.org/10.3390/cimb45020114
APA StyleKöse, F. A., Pabuccuoglu, A., Karakoyun, M., & Aydogdu, S. (2023). Peroxiredoxins and Hypoxia-Inducible Factor-1α in Duodenal Tissue: Emerging Factors in the Pathophysiology of Pediatric Celiac Disease Patients. Current Issues in Molecular Biology, 45(2), 1779-1793. https://doi.org/10.3390/cimb45020114