
Chemopreventive Effects of Onosma mutabilis against Azoxymethane-Induced Colon Cancer in Rats via Amendment of Bax/Bcl-2 and NF-κB Signaling Pathways

 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Extraction
2.2. Cancer Cell
2.3. MTT Essay
2.4. Animal Study and Ethical Approval
2.5. Acute Toxicity Test
2.6. Experimental Design for the Chemoprotective Test of OME
2.7. Scoring of Aberrant Crypts
2.8. Histological Examination of ACFs
2.9. Immunohistochemistry
2.10. Antioxidants of Colon Homogenates
2.11. Statistical Analysis
3. Results
3.1. In Vitro Anticancer Effects
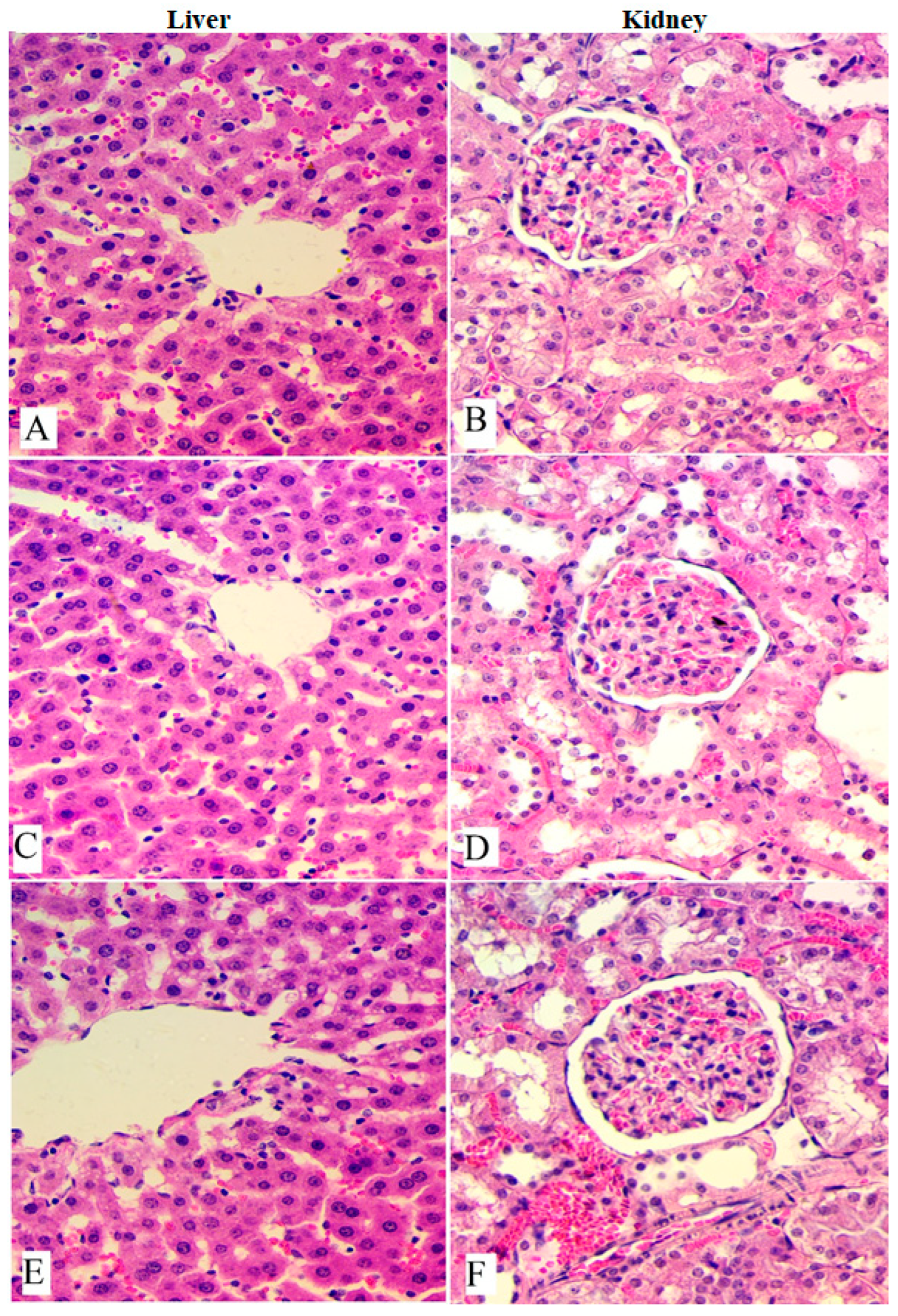
3.2. Acute Toxicity
3.3. In Vivo Anticancer Study
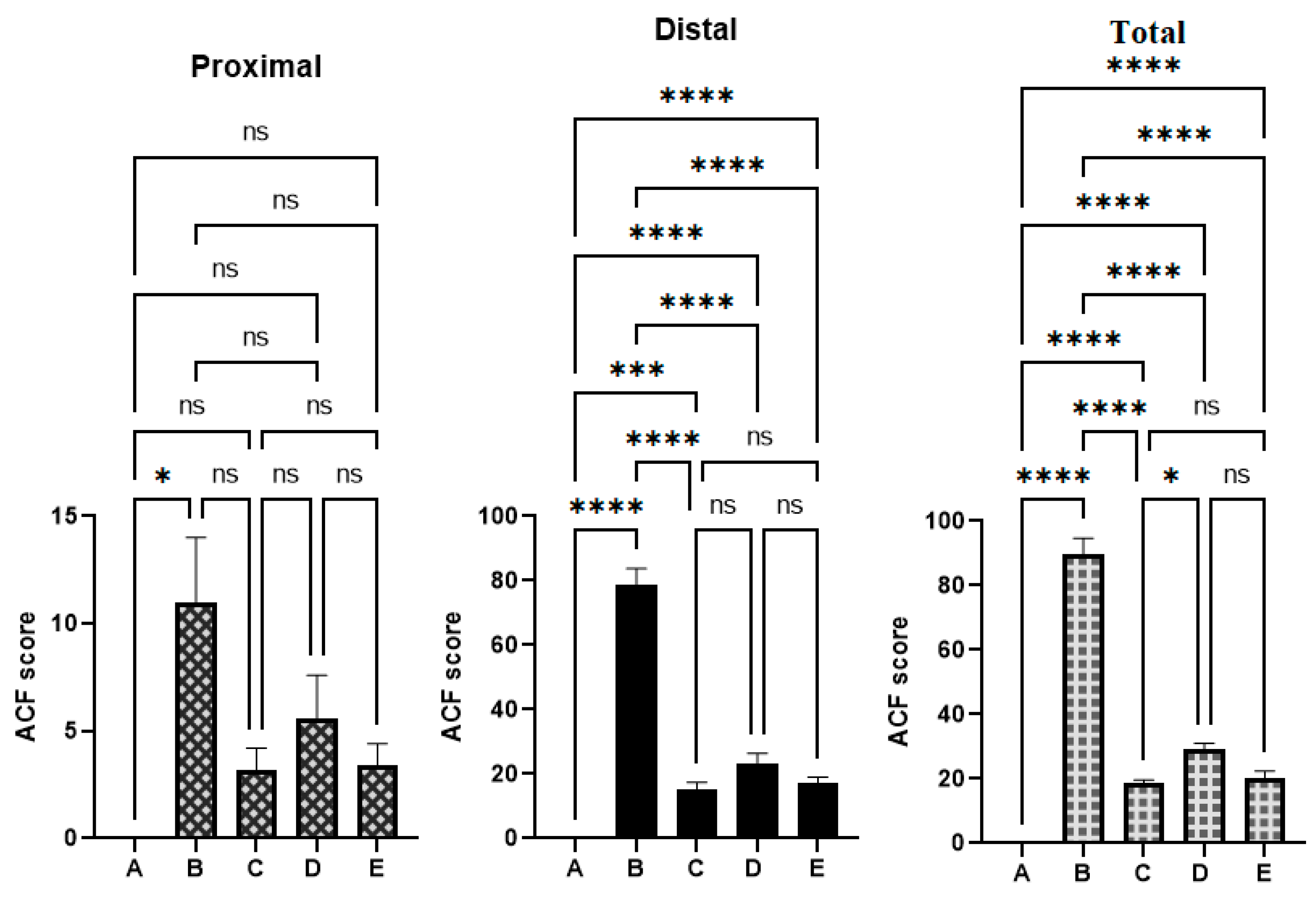
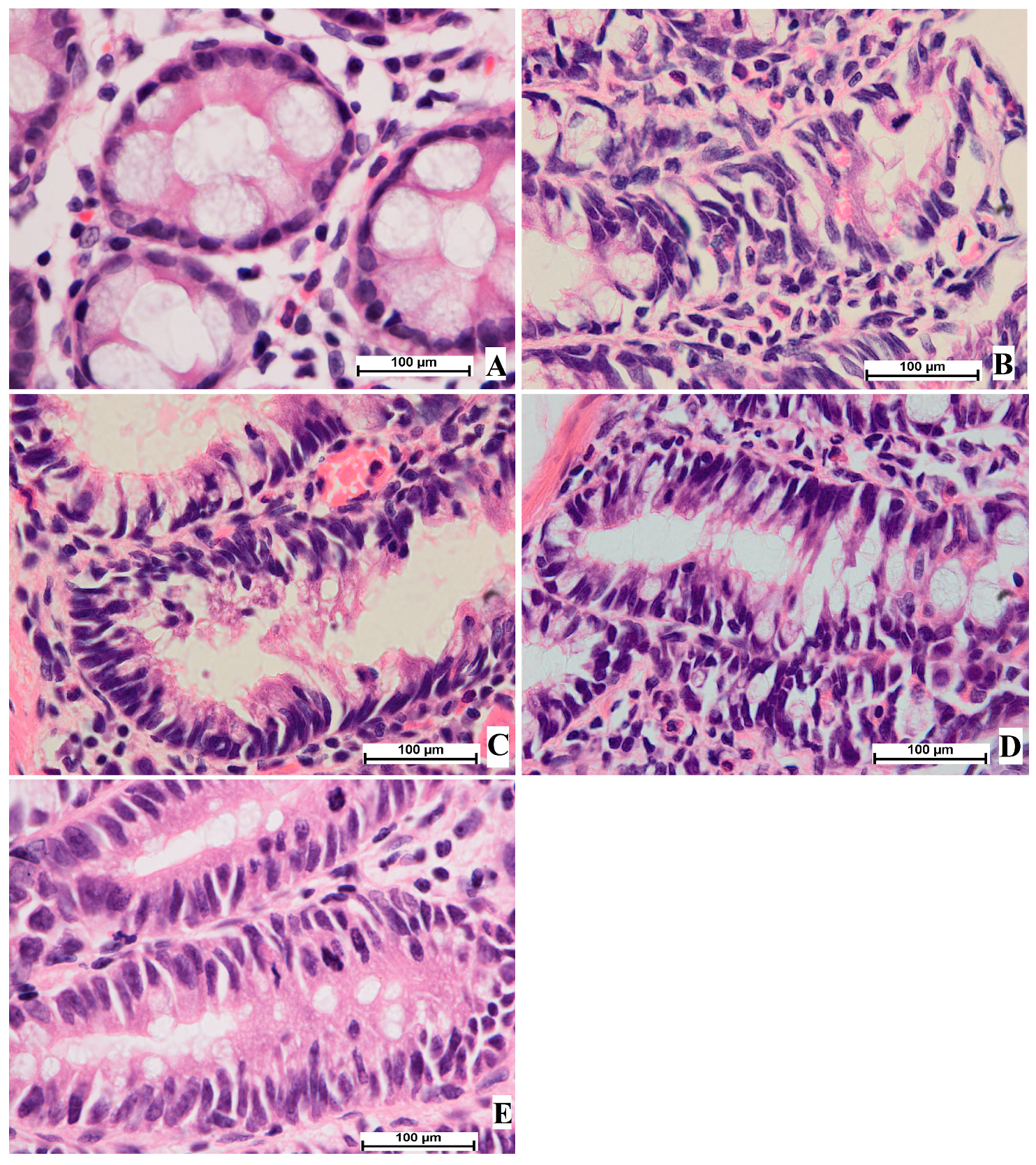
3.4. Histological Analysis of Aberrant Crypt Foci
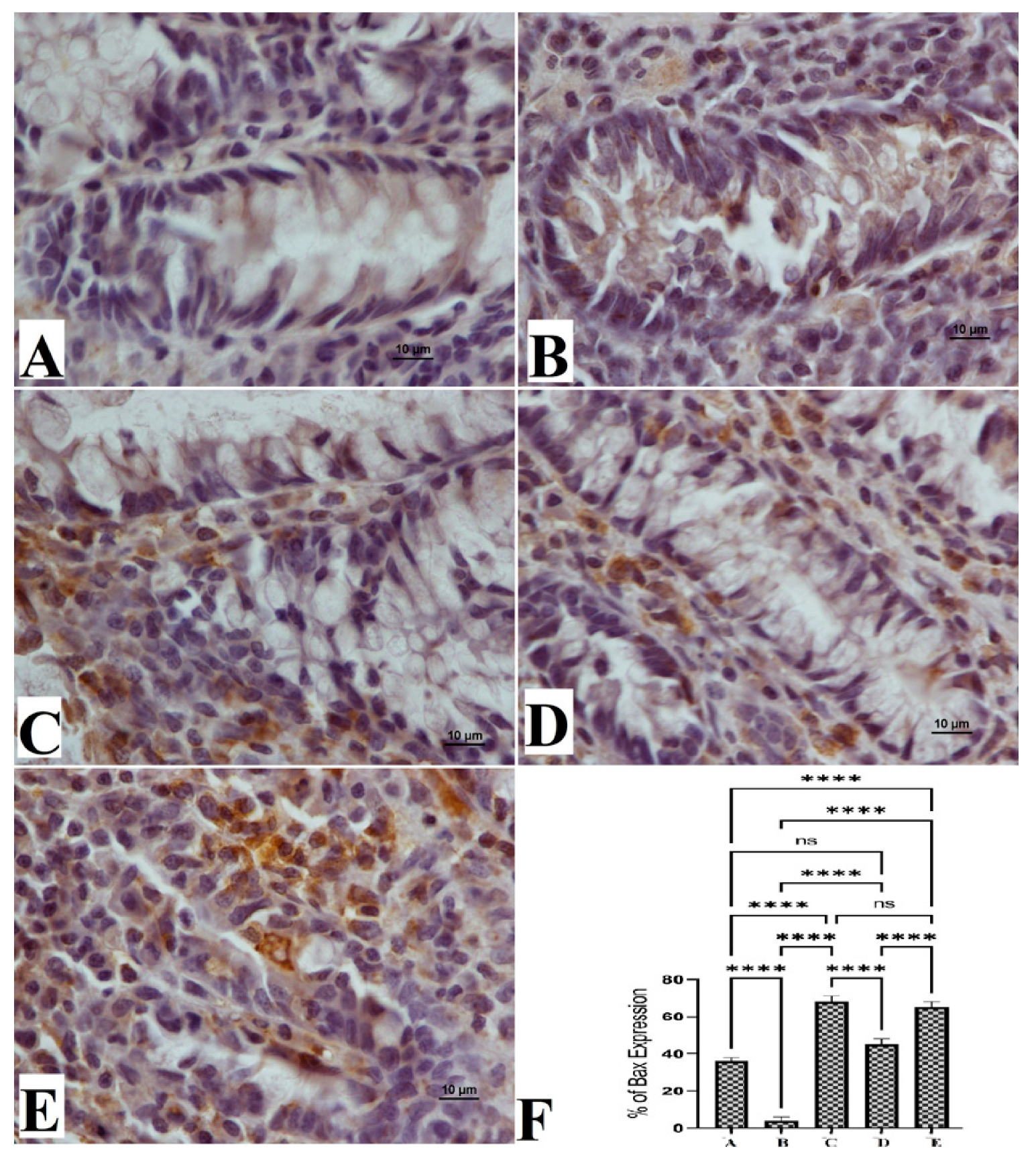
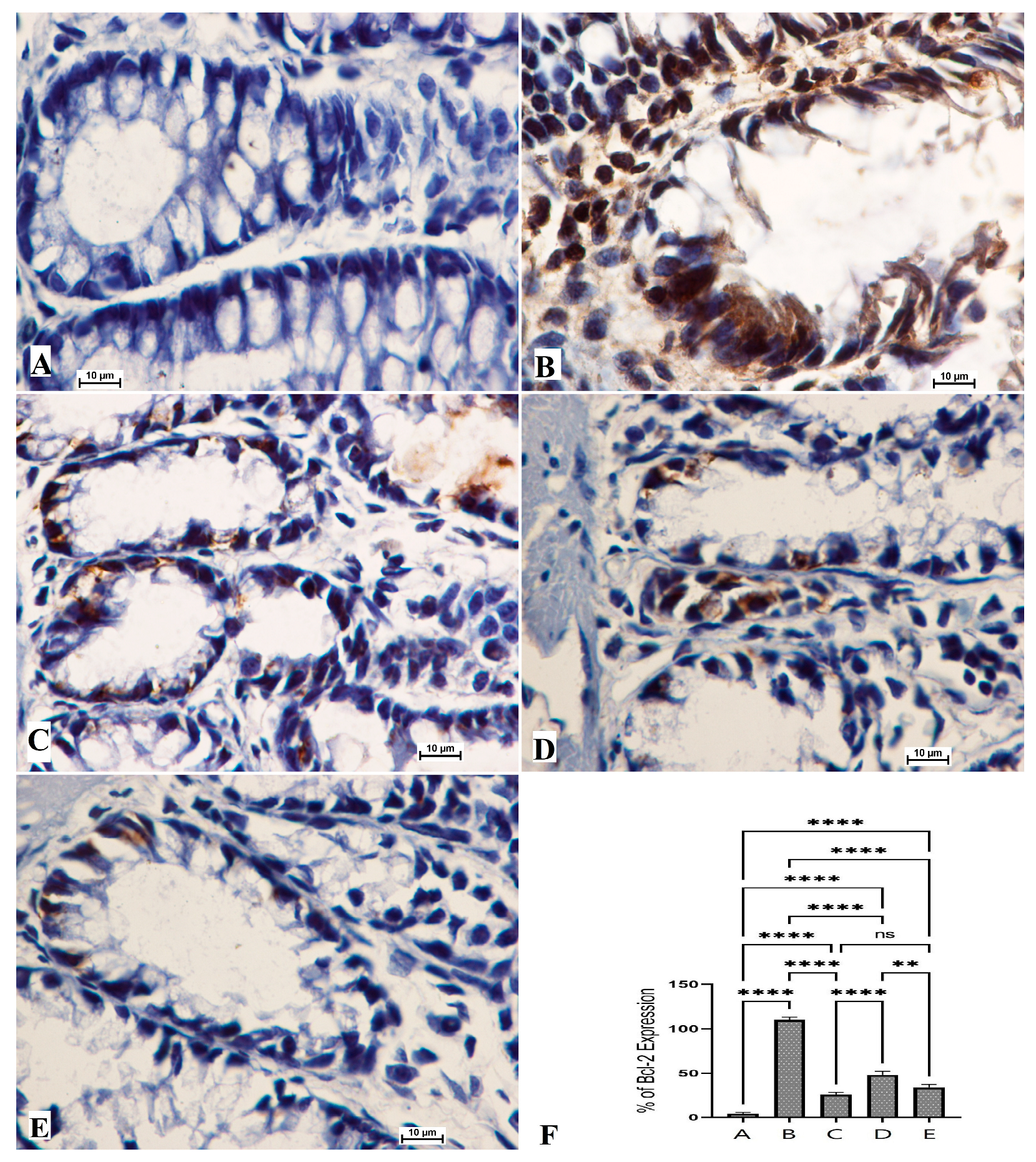
3.5. The Bax and Bcl-2 Protein Expressions
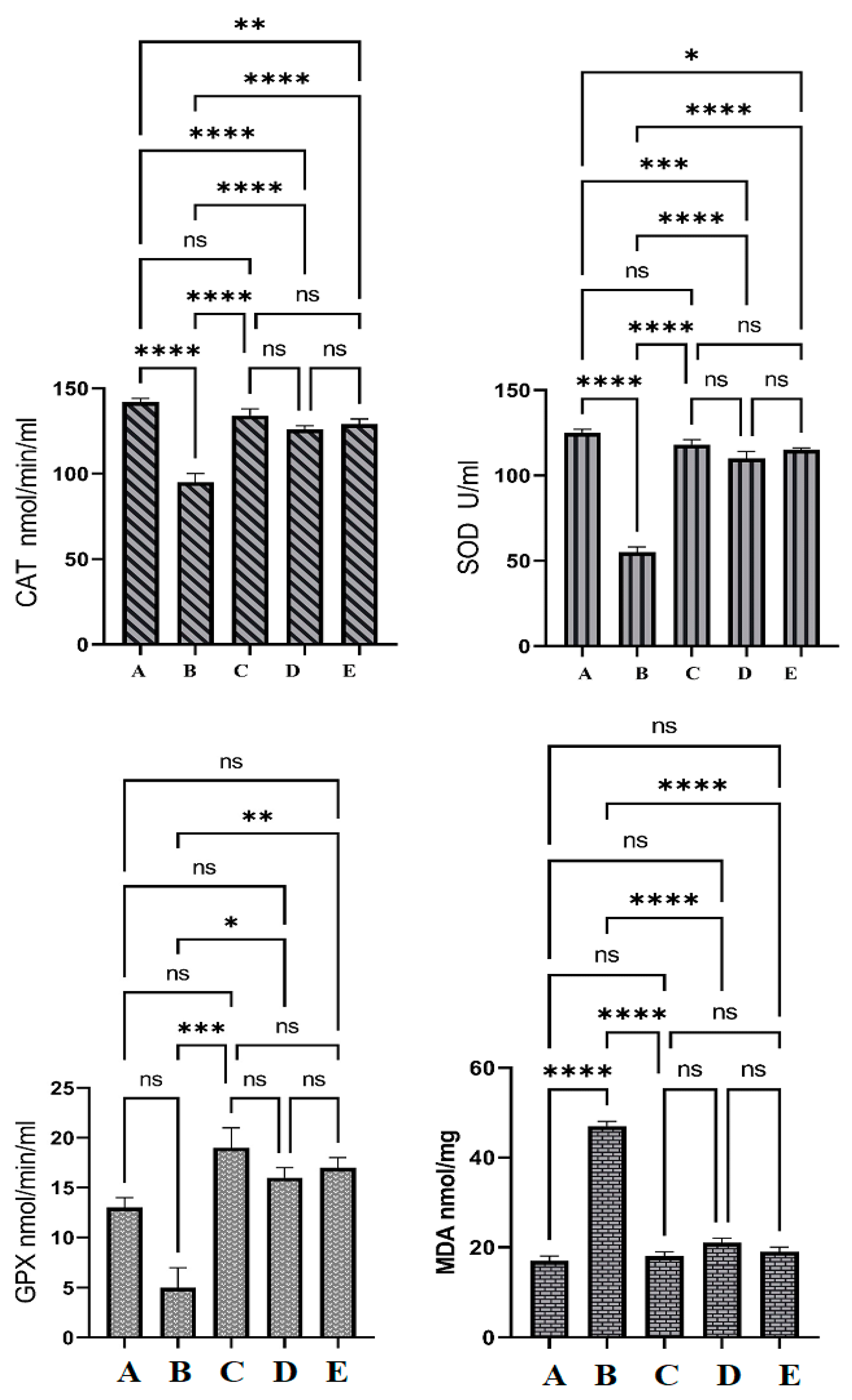
3.6. Antioxidative Enzyme Actions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, K.D.; Nogueira, L.; Devasia, T.; Mariotto, A.B.; Yabroff, K.R.; Jemal, A.; Kramer, J.; Siegel, R.L. Cancer treatment and survivorship statistics, 2022. CA Cancer J. Clin. 2022, 72, 409–436. [Google Scholar] [CrossRef]
- Luo, Q.; Lew, J.B.; Steinberg, J.; Worthington, J.; Yu, X.Q.; Caruana, M.; Soerjomataram, I.; Bray, F.; Lawrance, S.; Arcorace, M.; et al. Trends in colon and rectal cancer mortality in Australia from 1972 to 2015 and associated projections to 2040. Sci. Rep. 2022, 12, 3994. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Seker-Polat, F.; Pinarbasi Degirmenci, N.; Solaroglu, I.; Bagci-Onder, T. Tumor Cell Infiltration into the Brain in Glioblastoma: From Mechanisms to Clinical Perspectives. Cancers 2022, 14, 443. [Google Scholar] [CrossRef]
- Ozel-Tasci, C.; Gulec, S. Effects of Golden Thistle (Scolymus hispanicus L.) on Cytotoxic Activity: Cell Cycle Arrest and Apoptotic Properties on the CaCo-2 Cell Line. J. Med. Food 2022, 25, 523–528. [Google Scholar] [CrossRef]
- Esmeeta, A.; Adhikary, S.; Dharshnaa, V.; Swarnamughi, P.; Ummul Maqsummiya, Z.; Banerjee, A.; Pathak, S.; Duttaroy, A.K. Plant-derived bioactive compounds in colon cancer treatment: An updated review. Biomed. Pharmacother. 2022, 153, 113384. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Lau, H.C.-H.; Zhang, X.; Yu, J. Mouse Models for Application in Colorectal Cancer: Understanding the Pathogenesis and Relevance to the Human Condition. Biomedicines 2022, 10, 1710. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Li, P.; Liu, S.; Liu, Q.; Li, Y.; Zhang, Z.; Yang, C.; Hu, M.; Sun, Y.; He, C. Correction: The chemopreventive effects of Huangqin-tea against AOM-induced preneoplastic colonic aberrant crypt foci in rats and omics analysis. Food Funct. 2021, 12, 2336–2337. [Google Scholar] [CrossRef] [PubMed]
- Arango-Varela, S.S.; Luzardo-Ocampo, I.; Maldonado-Celis, M.E. Andean berry (Vaccinium meridionale Swartz) juice, in combination with Aspirin, displayed antiproliferative and pro-apoptotic mechanisms in vitro while exhibiting protective effects against AOM-induced colorectal cancer in vivo. Food Res. Int. 2022, 157, 111244. [Google Scholar] [CrossRef] [PubMed]
- Rayan, A.; Raiyn, J.; Falah, M. Nature is the best source of anticancer drugs: Indexing natural products for their anticancer bioactivity. PLoS ONE 2017, 12, e0187925. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, M.M.; Abo-Youssef, A.M.; Malak, M.N.; Hamzawy, M.A. Novel therapeutic modalities target cell signaling of Renin-Angiotensin system/NF-κB-induced cell cycle arrest and apoptosis in urethane-induced lung cancer in mice: An in vivo study. J. Biochem. Mol. Toxicol. 2022, 36, e23162. [Google Scholar] [CrossRef] [PubMed]
- Mašković, P.Z.; Diamanto, L.D.; Vujic, J.M.; Cvetanović, A.D.; Radojković, M.M.; Gadžurić, S.B.; Zengin, G. Onosma aucheriana: A source of biologically active molecules for novel food ingredients and pharmaceuticals. J. Funct. Foods 2015, 19, 479–486. [Google Scholar] [CrossRef]
- Jabbar, A.A.; Abdullah, F.O.; Hassan, A.O.; Galali, Y.; Hassan, R.R.; Rashid, E.Q.; Salih, M.I.; Aziz, K.F. Ethnobotanical, Phytochemistry, and Pharmacological Activity of Onosma (Boraginaceae): An Updated Review. Molecules 2022, 27, 8687. [Google Scholar] [CrossRef]
- Erenler, R.; Yildiz, I.; Aydin, A.; Genc, N. Antiproliferative and cytotoxic effects of bioactive compounds isolated from Onosma bourgaei. Med. Oncol. 2022, 39, 116. [Google Scholar] [CrossRef]
- Kumar, A.; Kaur, S.; Dhiman, S.; Singh, P.P.; Bhatia, G.; Thakur, S.; Tuli, H.S.; Sharma, U.; Kumar, S.; Almutary, A.G. Targeting Akt/NF-κB/p53 Pathway and Apoptosis Inducing Potential of 1, 2-Benzenedicarboxylic Acid, Bis (2-Methyl Propyl) Ester Isolated from Onosma bracteata Wall. against Human Osteosarcoma (MG-63) Cells. Molecules 2022, 27, 3478. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, A.A. Onosma mutabilis: Phytochemical composition, antioxidant, cytotoxicity, and acute oral toxicity. Food Sci. Nutr. 2021, 9, 5755–5764. [Google Scholar] [CrossRef]
- Maskovic, P.Z.; Mira, A.C.; Pavlovic, M.; Vujosevic, M.R.; Blagojevic, J.V.; Djuric, M.; Moracanin, S.V.; Djukic, D.A. A study on the ethanolic extract of Onosma aucheriana biological and toxicological evaluation. Rev. Chim. 2016, 67, 2511–2518. [Google Scholar]
- Jabbar, A.A.; Abdullah, F.O.; Abdulrahman, K.K.; Galali, Y.; Sardar, A.S. GC-MS Analysis of Bioactive Compounds in Methanolic Extracts of Papaver decaisnei and Determination of Its Antioxidants and Anticancer Activities. J. Food Qual. 2022, 2022, 1405157. [Google Scholar] [CrossRef]
- Garber, J.C.; Barbee, R.W.; Bielitzki, J.T.; Clayton, L.A.; Donovan, J.C.; Hendriksen, C.F.; Kohn, D.F.; Lipman, N.S.; Locke, P.A.; Melcher, J.; et al. Guide for the Care and Use of Laboratory Animals; The National Academic Press: Washington, DC, USA, 2011; ISBN 978-0-309-15400-0. [Google Scholar]
- Schlede, E.; Genschow, E.; Spielmann, H.; Stropp, G.; Kayser, D. Oral acute toxic class method: A successful alternative to the oral LD50 test. Regul. Toxicol. Pharmacol. 2005, 42, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Al-Medhtiy, M.H.; Jabbar, A.A.; Shareef, S.H.; Ibrahim, I.A.A.; Alzahrani, A.R.; Abdulla, M.A. Histopathological Evaluation of Annona muricata in TAA-Induced Liver Injury in Rats. Processes 2022, 10, 1613. [Google Scholar] [CrossRef]
- Flores-Romero, H.; Hohorst, L.; John, M.; Albert, M.; King, L.E.; Beckmann, L.; Szabo, T.; Hertlein, V.; Luo, X.; Villunger, A. BCL-2-family protein tBID can act as a BAX-like effector of apoptosis. EMBO J. 2022, 41, e108690. [Google Scholar] [CrossRef]
- Kaur, J.; Mahey, S.; Ahluwalia, P.; Joshi, R.; Kumar, R. Role of Plant Secondary Metabolites as Anticancer and Chemopreventive Agents. In Plant Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2022; pp. 97–119. [Google Scholar]
- Ben Kridis, W.; Mnif, A.; Khmiri, S.; Toumi, N.; Khanfir, A. Self-medication with herbal medicine and breast cancer survival: A prospective monocentric study. J. Cancer Res. Clin. Oncol. 2021, 147, 3401–3407. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, H.; Chen, X.; Gao, J.; Duan, Y.; Wei, D.; Zhang, J.; Ge, K.; Liang, X.-J.; Huang, Y. Nano-herb medicine and PDT induced synergistic immunotherapy for colon cancer treatment. Biomaterials 2021, 269, 120654. [Google Scholar] [CrossRef]
- Manjunath, C.; Mahurkar, N. In vitro cytotoxicity of cardamom oil, lemon oil, and jasmine oil on human skin, gastric, and brain cancer cell line. J. Cancer Res. Ther. 2021, 17, 62. [Google Scholar] [CrossRef]
- Kostić, A.Ž.; Mačukanović-Jocić, M.P.; Milinčić, D.D.; Petrović, J.D.; Gašić, U.M.; Gligorijević, N.N.; Jarić, S.V.; Soković, M.D.; Tešić, Ž.L.; Pešić, M.B.; et al. Hieracium waldsteinii (Asteraceae) and Onosma stellulata (Boraginaceae) as a Source of Antioxidant and Antimicrobial Agents. Chem. Biodivers. 2022, 19, e202200069. [Google Scholar] [CrossRef] [PubMed]
- Dilshara, M.G.; Karunarathne, W.A.H.M.; Molagoda, I.M.N.; Kang, C.-H.; Jeong, J.-W.; Choi, Y.H.; Kim, G.-Y. β-Hydroxyisovalerylshikonin promotes reactive oxygen species production in HCT116 colon cancer cells, leading to caspase-mediated apoptosis. Rev. Bras. Farmacogn. 2018, 28, 344–351. [Google Scholar] [CrossRef]
- Wu, R.; Shen, Q.; Li, P.; Shang, N. Sturgeon Chondroitin Sulfate Restores the Balance of Gut Microbiota in Colorectal Cancer Bearing Mice. Int. J. Mol. Sci. 2022, 23, 3723. [Google Scholar] [CrossRef]
- Shilnikova, K.; Piao, M.J.; Kang, K.A.; Fernando, P.D.S.M.; Herath, H.M.U.L.; Cho, S.J.; Hyun, J.W. Natural compound Shikonin induces apoptosis and attenuates epithelial to mesenchymal transition in radiation-resistant human colon cancer cells. Biomol. Ther. 2022, 30, 137. [Google Scholar] [CrossRef]
- Jabbar, A.A.; Abdullah, F.O.; Abdoulrahman, K.; Galali, Y.; Ibrahim, I.A.; Alzahrani, A.R.; Hassan, R.R. Gastroprotective, Biochemical, and Acute Toxicity Effects of Papaver decaisnei against Ethanol-Induced Gastric Ulcers in Rats. Processes 2022, 10, 1985. [Google Scholar] [CrossRef]
- Gajendran, B.; Durai, P.; Varier, K.M.; Chinnasamy, A. A novel phytosterol isolated from Datura inoxia, RinoxiaB is a potential cure colon cancer agent by targeting BAX/Bcl2 pathway. Bioorg. Med. Chem. 2020, 28, 115242. [Google Scholar] [CrossRef] [PubMed]
- Shareef, S.H.; Al-Medhtiy, M.H.; Ibrahim, I.A.; Alzahrani, A.R.; Jabbar, A.A.; Galali, Y.; Agha, N.F.; Aziz, P.Y.; Thabit, M.A.; Agha, D.N.F.; et al. Gastroprophylactic Effects of p-Cymene in Ethanol-Induced Gastric Ulcer in Rats. Process 2022, 10, 1314. [Google Scholar] [CrossRef]
- Badr, A.M.; EL-Orabi, N.F.; Ali, R.A. The implication of the crosstalk of Nrf2 with NOXs, and HMGB1 in ethanol-induced gastric ulcer: Potential protective effect is afforded by Raspberry Ketone. PLoS ONE 2019, 14, e0220548. [Google Scholar] [CrossRef] [Green Version]
- Tomar, V.; Kukreti, S.; Prakash, S.; Madan, J.; Chandra, R. Noscapine and its Analogs as Chemotherapeutic Agent: Current updates. Curr. Top. Med. Chem. 2017, 17, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.-B.; Tian, Z.-S.; Gui, S.-L.; Cui, W.-G.; Li, W. Anti-prostate cancer effect of roemerine: An experimental study. Zhonghua Nan Ke Xue 2017, 23, 27–33. [Google Scholar] [PubMed]
- Capriello, T.; Monteiro, S.M.; Félix, L.M.; Donizetti, A.; Aliperti, V.; Ferrandino, I. Apoptosis, oxidative stress and genotoxicity in developing zebrafish after aluminium exposure. Aquat. Toxicol. 2021, 236, 105872. [Google Scholar] [CrossRef]
- Li, C.; Wang, L.; Zhao, J.; Wei, Y.; Zhai, S.; Tan, M.; Guan, K.; Huang, Z.; Chen, C. Lonicera rupicola Hook.f.et Thoms flavonoids ameliorated dysregulated inflammatory responses, intestinal barrier, and gut microbiome in ulcerative colitis via PI3K/AKT pathway. Phytomedicine 2022, 104, 154284. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Mao, X.; Chen, Y.; Yan, L.; Ye, L.; Li, S. Reactive Oxygen Species Induce Fatty Liver and Ischemia-Reperfusion Injury by Promoting Inflammation and Cell Death. Front. Immunol. 2022, 13, 2029. [Google Scholar] [CrossRef]
- Byrne, F.L.; Olzomer, E.M.; Marriott, G.R.; Quek, L.-E.; Katen, A.; Su, J.; Nelson, M.E.; Hart-Smith, G.; Larance, M.; Sebesfi, V.F. Phenotypic screen for oxygen consumption rate identifies an anti-cancer naphthoquinone that induces mitochondrial oxidative stress. Redox Biol. 2020, 28, 101374. [Google Scholar] [CrossRef]
- Wu, Q.; Er-bu, A.; Liang, X.; He, C.; Yin, L.; Xu, F.; Zou, Y.; Yin, Z.; Yue, G.; Li, L.; et al. Isolation, structure identification, and immunostimulatory effects in vitro and in vivo of polysaccharides from Onosma hookeri Clarke var. longiforum Duthie. J. Sci. Food Agric. 2023, 103, 328–338. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Sahinler, S.S.; Ceylan, O.; Tepe, B. Onosma pulchra: Phytochemical composition, antioxidant, skin-whitening and anti-diabetic activity. Ind. Crops Prod. 2020, 154, 112632. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Sahinler, S.S.; Ceylan, O.; Tepe, B. Onosma ambigens: Phytochemical composition, antioxidant and enzyme inhibitory activity. Ind. Crops Prod. 2020, 154, 112651. [Google Scholar] [CrossRef]
- Shoaib, A.; Siddiqui, H.H.; Badruddeen; Dixit, R.K. Evaluation of Noxious Consequence of Bark Extract of Onosma echioides Linn Root: Hematology, Biochemistry, and Histopathological Findings. J. Diet. Suppl. 2020, 17, 110–119. [Google Scholar] [CrossRef]
- Jabbar, A.A.; Abdulrahman, K.K.; Abdulsamad, P.; Mojarrad, S.; Mehmetçik, G. Phytochemical profile, Antioxidant, Enzyme inhibitory and acute toxicity activity of Astragalus bruguieri. Baghdad Sci. J. 2022, 20, 157–165. [Google Scholar]
- Abudoulrahman, K.K.; Mustafa, M.A.; Abduljabbar, A.A. The Effect of Heat Stress on Oxidative Stress and Antioxidant Status in Local Quail Hens Supplemented with Onion and Garlic Oils. Tikrit J. Agric. Sci. 2019, 19, 103–110. [Google Scholar] [CrossRef]
- Hasanvand, D.; Amiri, I.; Soleimani Asl, S.; Saidijam, M.; Shabab, N.; Artimani, T. Effects of CeO(2) nanoparticles on the HO-1, NQO1, and GCLC expression in the testes of diabetic rats. Can. J. Physiol. Pharmacol. 2018, 96, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Liu, Y.; Li, C.; Zhuo, F.; Xu, Z.; Hong, H.; Sun, H.; Huang, X.; Yu, X. Oxidative Stress Responses and Gene Transcription of Mice under Chronic-Exposure to 2, 6-Dichlorobenzoquinone. Int. J. Environ. Res. Public Health 2022, 19, 13801. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-S.; Lin, S.-C.; Chu, C.-H.; Chang, Y.-K.; Zhang, X.; Lin, C.-C.; Tung, Y.-T. The Gastroprotective Effect of Naringenin against Ethanol-Induced Gastric Ulcers in Mice through Inhibiting Oxidative and Inflammatory Responses. Int. J. Mol. Sci. 2021, 22, 11985. [Google Scholar] [CrossRef]
- Prescott, J.A.; Mitchell, J.P.; Cook, S.J. Inhibitory feedback control of NF-κB signalling in health and disease. Biochem. J. 2021, 478, 2619–2664. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.A.; Shaukat, A.; Wu, K.; Rajput, I.R.; Baloch, D.M.; Akhtar, R.W.; Raza, M.A.; Najda, A.; Rafał, P.; Albrakati, A. Luteolin alleviates aflatoxinB1-induced apoptosis and oxidative stress in the liver of mice through activation of Nrf2 signaling pathway. Antioxidants 2021, 10, 1268. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Youn, K.; Jun, M. Protective effect of sargahydroquinoic acid against Aβ25–35-evoked damage via PI3K/Akt mediated Nrf2 antioxidant defense system. Biomed. Pharmacother. 2021, 144, 112271. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, N.; Kandwal, P.; Sharma, G.; Gambhir, L. Redox ticklers and beyond: Naphthoquinone repository in the spotlight against inflammation and associated maladies. Pharmacol. Res. 2021, 174, 105968. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Deng, Z.-Y.; Chen, X.; Zhang, B.; Fan, Y.; Li, H. Synergistic antioxidant effects of phenolic acids and carotenes on H2O2-induced H9c2 cells: Role of cell membrane transporters. Food Chem. 2021, 341, 128000. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell | IC50 Values (μg/mL) 1 | |
|---|---|---|
| O. mutabilis MeOH Extract | DOX 2 | |
| Caco-2 | 22.28 ± 0.45 a | 0.83 ± 0.28 |
| HT-29 | 36.55 ± 1.20 b | 0.62 ± 0.32 |
| NHDF | 173.26 ± 1.89 c | 1.22 ± 0.15 |
| Biochemical Parameters | Control | LD | HD |
|---|---|---|---|
| LFT | |||
| Albumin g/L | 32.2± 1.8 a | 33.1 ± 2.0 a | 33.1 ± 2.0 a |
| Total bilirubin mmol/L | <2 a | <2 a | <2 a |
| Alkaline phosphatase U/L | 166.2 ± 24.4 a | 162.71 ± 23.2 a | 164.35 ± 36.8 a |
| Alanine aminotransferase U/L | 78.32 ± 11.8 a | 80.12 ± 14.1 a | 81.34 ± 12.2 a |
| G-glutamyl transferase U/L | 0.07 ± 0.5 a | 0.08 ± 0.4 a | 0.09 ± 0.3 a |
| RFT | |||
| Sodium mmol/L | 139.2 ± 5.2 a | 142.5 ± 4.2 a | 143.4 ± 2.2 a |
| Potassium mmol/L | 4.9 ± 1.6 a | 5.1 ± 1.8 a | 5.7 ± 2.5 a |
| Chloride mmol/L | 102.5 ± 3.6 a | 104.08 ± 3.2 a | 105.12 ± 6.2 a |
| Carbon Dioxide mmol/L | 34.30 ± 1.9 a | 35.22 ± 1.3 a | 36.70 ± 2.3 |
| Anion gap mmol/L | 14.4 ± 1.4 a | 12.80 ± 1.7 b | 11.60 ± 3.5 b |
| Urea mmol/L | 5.80 ± 1.4 a | 6.10 ± 1.8 a | 6.8 ± 5.8 a |
| Creatinine umol/L | 31.20 ± 3.2 a | 29.10 ± 3.0 a | 28.52 ± 5.0 b |
| Number of Foci Containing | ||||||
|---|---|---|---|---|---|---|
| Groups | 1 Crypt | 2 Crypt | 3 Crypt | ≥4 | Total | Inhibition % |
| Crypt | ACF | |||||
| A | 0 | 0 | 0 | 0 | 0 | N/A |
| B | 11.0 ± 3 c | 28.4 ± 4 c | 21.3 ± 1 c | 29.0 ± 1 c | 89.7 ± 5 c | N/A |
| C | 3.2 ± 1 a | 6.7 ± 2 a | 3.3 ± 2 a | 5.4 ± 1 a | 18.6 ± 1 a | 79.2 a |
| D | 5.6 ± 2 a | 8.3 ± 2 a | 7.0 ± 2 b | 8.1 ± 1 a | 29.0 ± 2 a | 67.5 b |
| E | 3.4 ± 1 a | 7.3 ± 1 a | 3.5 ± 2 a | 6.2 ± 2 a | 20.4 ± 2 a | 77.25 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabbar, A.A.; Ibrahim, I.A.A.; Abdullah, F.O.; Aziz, K.F.; Alzahrani, A.R.; Abdulla, M.A. Chemopreventive Effects of Onosma mutabilis against Azoxymethane-Induced Colon Cancer in Rats via Amendment of Bax/Bcl-2 and NF-κB Signaling Pathways. Curr. Issues Mol. Biol. 2023, 45, 885-902. https://doi.org/10.3390/cimb45020057
Jabbar AA, Ibrahim IAA, Abdullah FO, Aziz KF, Alzahrani AR, Abdulla MA. Chemopreventive Effects of Onosma mutabilis against Azoxymethane-Induced Colon Cancer in Rats via Amendment of Bax/Bcl-2 and NF-κB Signaling Pathways. Current Issues in Molecular Biology. 2023; 45(2):885-902. https://doi.org/10.3390/cimb45020057
Chicago/Turabian StyleJabbar, Ahmed Aj., Ibrahim Abdel Aziz Ibrahim, Fuad O. Abdullah, Kareem Fattah Aziz, Abdullah R. Alzahrani, and Mahmood Ameen Abdulla. 2023. "Chemopreventive Effects of Onosma mutabilis against Azoxymethane-Induced Colon Cancer in Rats via Amendment of Bax/Bcl-2 and NF-κB Signaling Pathways" Current Issues in Molecular Biology 45, no. 2: 885-902. https://doi.org/10.3390/cimb45020057
APA StyleJabbar, A. A., Ibrahim, I. A. A., Abdullah, F. O., Aziz, K. F., Alzahrani, A. R., & Abdulla, M. A. (2023). Chemopreventive Effects of Onosma mutabilis against Azoxymethane-Induced Colon Cancer in Rats via Amendment of Bax/Bcl-2 and NF-κB Signaling Pathways. Current Issues in Molecular Biology, 45(2), 885-902. https://doi.org/10.3390/cimb45020057