
Disarm The Bacteria: What Temperate Phages Can Do

Abstract
:1. Introduction
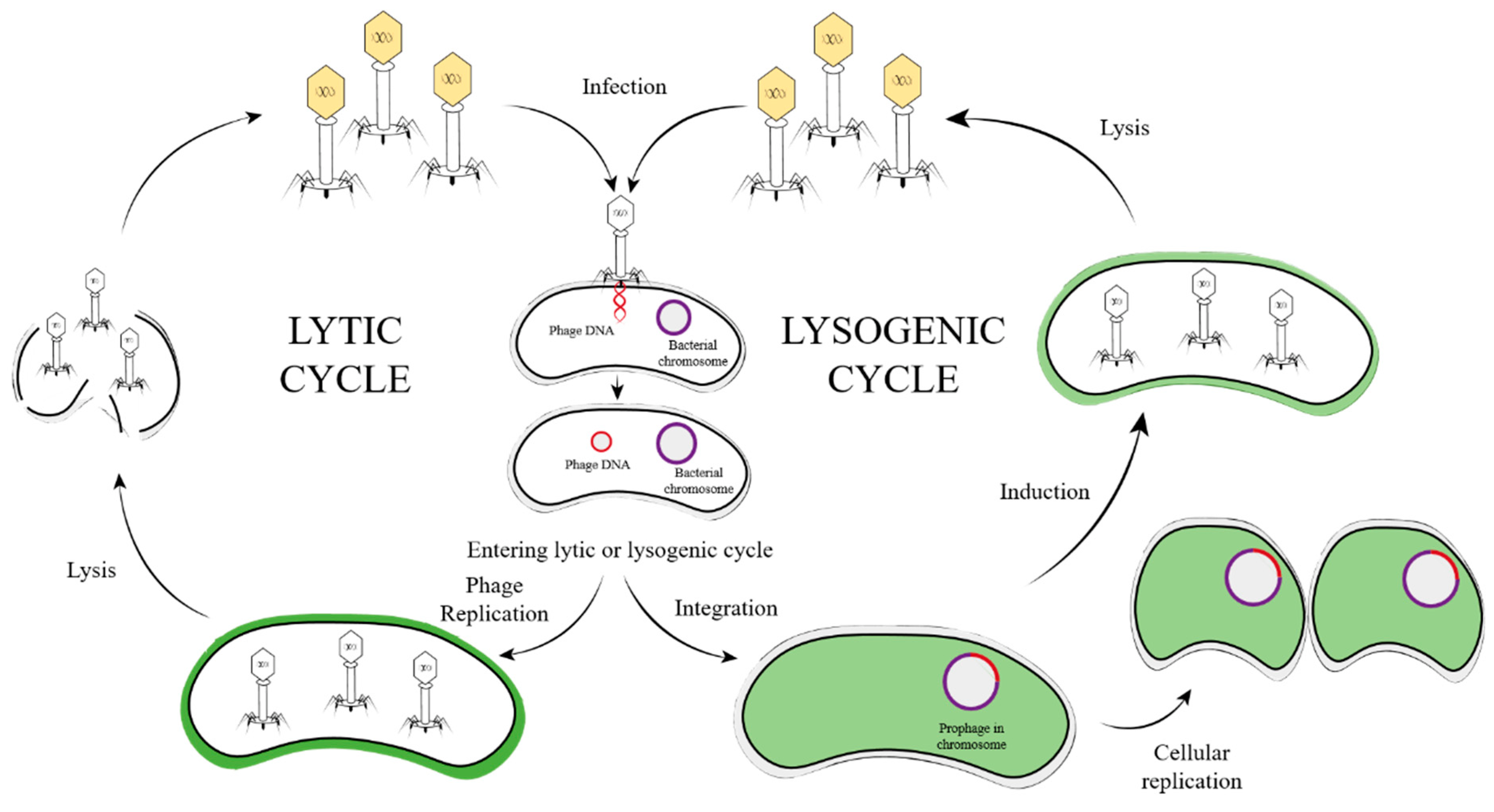
2. Temperate Phage
3. Temperate Phage Life Cycle
3.1. Step One—Diffusion
3.2. Step Two—Absorption and Injection
3.3. Step Three—Integration and Replication
3.4. Step Four—Induction and Packaging
3.5. Step Five—Lysis of Host Bacteria
4. Temperate Phages for Therapeutic Purposes
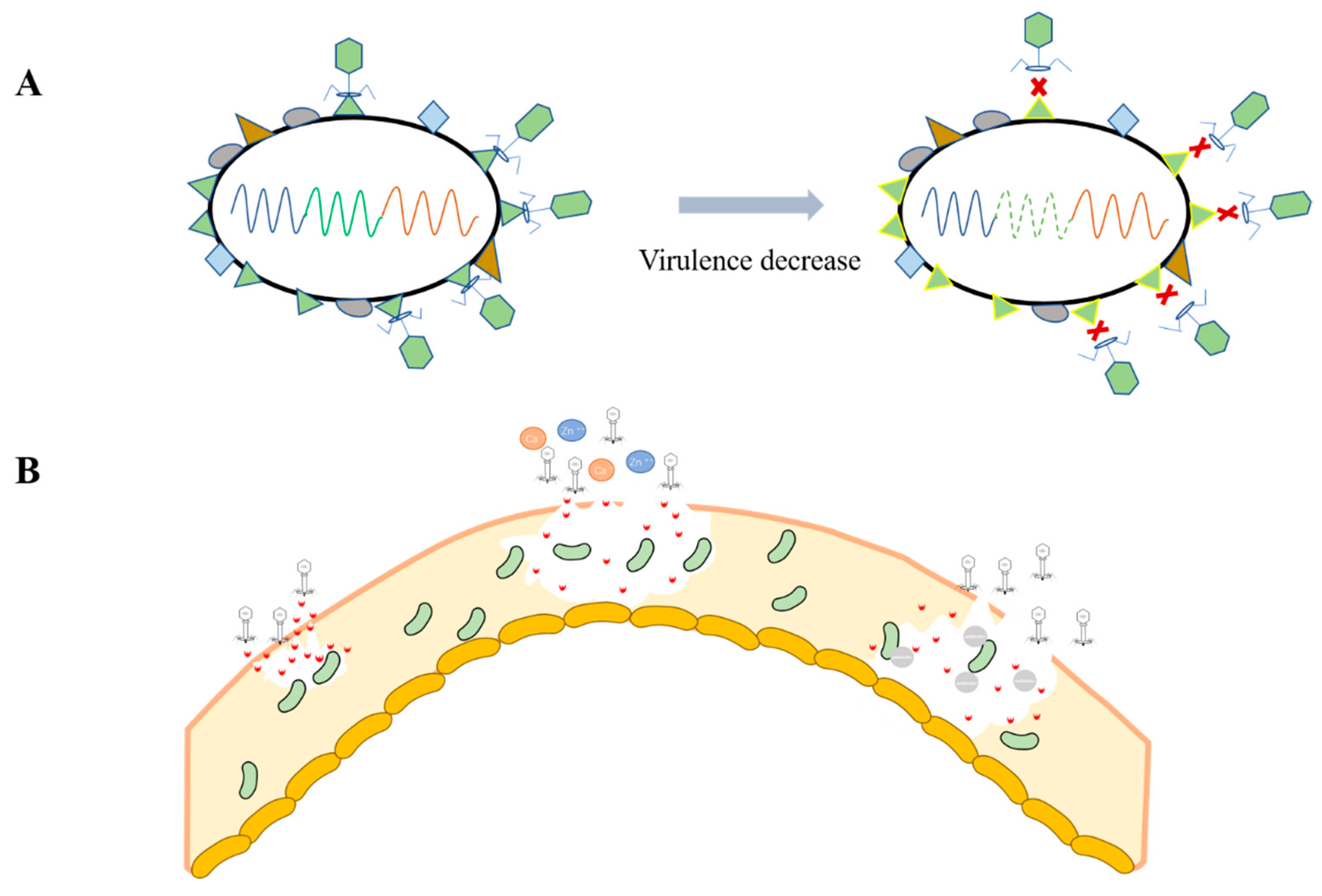
4.1. Host Virulence Reduction
4.2. Biofilm Degradation
5. Temperate Phage Gene Engineering and Display
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rohwer, F.; Edwards, R. The Phage Proteomic Tree: A Genome-Based Taxonomy for Phage. J. Bacteriol. 2002, 184, 4529–4535. [Google Scholar] [CrossRef] [PubMed]
- Watts, G. Phage therapy: Revival of the bygone antimicrobial. Lancet (London, England) 2017, 390, 2539–2540. [Google Scholar] [CrossRef]
- Fruciano, D.E.; Bourne, S. Phage as an Antimicrobial Agent: D’Herelle’s Heretical Theories and Their Role in the Decline of Phage Prophylaxis in the West. Can. J. Infect. Dis. Med. Microbiol. 2007, 18, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Gulumbe, B.H.; Haruna, U.A.; Almazan, J.; Ibrahim, I.H.; Faggo, A.A.; Bazata, A.Y. Combating the menace of antimicrobial resistance in Africa: A review on stewardship, surveillance and diagnostic strategies. Biol. Proced. Online 2022, 24, 1–13. [Google Scholar] [CrossRef]
- Gelman, D.; Eisenkraft, A.; Chanishvili, N.; Nachman, D.; Glazer, S.C.; Hazan, R. The history and promising future of phage therapy in the military service. J. Trauma: Inj. Infect. Crit. Care 2018, 85 (Suppl. S2), S18–S26. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, L.; Sun, E.; Song, J.; Wu, B. Characterisation of a newly detected bacteriophage infecting Bordetella bronchiseptica in swine. Arch. Virol. 2019, 164, 33–40. [Google Scholar] [CrossRef]
- Chen, Y.; Li, X.; Wang, S.; Guan, L.; Li, X.; Hu, D.; Gao, D.; Song, J.; Chen, H.; Qian, P. A Novel Tail-Associated O91-Specific Polysaccharide Depolymerase from a Podophage Reveals Lytic Efficacy of Shiga Toxin-Producing Escherichia coli. Appl. Environ. Microbiol. 2020, 86, e00145-20. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, E.; Song, J.; Yang, L.; Wu, B. Complete Genome Sequence of a Novel T7-Like Bacteriophage from a Pasteurella multocida Capsular Type A Isolate. Curr. Microbiol. 2018, 75, 574–579. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, S.; Wang, Y.; Zhangxiang, L.; Chen, H.; Li, X.; Qian, P. Complete genome sequence of the novel phage vB_EcoS_PHB17, which infects Shiga-toxin-producing Escherichia coli. Arch. Virol. 2019, 164, 3111–3113. [Google Scholar] [CrossRef]
- Maiques, E.; Úbeda, C.; Campoy, S.; Salvador, N.; Lasa, I.; Novick, R.P.; Barbé, J.; Penadés, J.R. β-Lactam Antibiotics Induce the SOS Response and Horizontal Transfer of Virulence Factors in Staphylococcus aureus. J. Bacteriol. 2006, 188, 2726–2729. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sun, E.; Yang, L.; Song, J.; Wu, B. Therapeutic Application of Bacteriophage PHB02 and Its Putative Depolymerase Against Pasteurella multocida Capsular Type A in Mice. Front. Microbiol. 2018, 9, 1678. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guo, G.; Sun, E.; Song, J.; Yang, L.; Zhu, L.; Liang, W.; Hua, L.; Peng, Z.; Tang, X.; et al. Isolation of a T7-Like Lytic Pasteurella Bacteriophage vB_PmuP_PHB01 and Its Potential Use in Therapy against Pasteurella multocida Infections. Viruses 2019, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sun, E.; Song, J.; Tong, Y.; Wu, B. Three Salmonella enterica serovar Enteritidis bacteriophages from the Siphoviridae family are promising candidates for phage therapy. Can. J. Microbiol. 2018, 64, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.R.J.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Olszak, T.; Latka, A.; Roszniowski, B.; Valvano, M.A.; Drulis-Kawa, Z. Phage Life Cycles Behind Bacterial Biodiversity. Curr. Med. Chem. 2017, 24, 3987–4001. [Google Scholar] [CrossRef]
- Albright, M.B.N.; Gallegos-Graves, L.V.; Feeser, K.L.; Montoya, K.; Emerson, J.B.; Shakya, M.; Dunbar, J. Experimental evidence for the impact of soil viruses on carbon cycling during surface plant litter decomposition. ISME Commun. 2022, 2, 24. [Google Scholar] [CrossRef]
- Weinbauer, M.G. Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef]
- Shkoporov, A.N.; Turkington, C.J.; Hill, C. Mutualistic interplay between bacteriophages and bacteria in the human gut. Nat. Rev. Genet. 2022, 20, 737–749. [Google Scholar] [CrossRef]
- Chen, Y.; Li, X.; Song, J.; Yang, D.; Liu, W.; Chen, H.; Wu, B.; Qian, P. Isolation and characterization of a novel temperate bacteriophage from gut-associated Escherichia within black soldier fly larvae (Hermetia illucens L. [Diptera: Stratiomyidae]). Arch. Virol. 2019, 164, 2277–2284. [Google Scholar] [CrossRef]
- Turner, D.; Kropinski, A.; Adriaenssens, E. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Genet. 2020, 18, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Hampton, H.G.; Watson, B.N.J.; Fineran, P.C. The arms race between bacteria and their phage foes. Nature 2020, 577, 327–336. [Google Scholar] [CrossRef]
- Rohwer, F.; Segall, A.M. A century of phage lessons. Nature 2015, 528, 46–47. [Google Scholar] [CrossRef]
- Taylor, V.L.; Fitzpatrick, A.D.; Islam, Z.; Maxwell, K.L. The Diverse Impacts of Phage Morons on Bacterial Fitness and Virulence. Adv. Virus Res. 2019, 103, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Panis, G.; Franche, N.; Méjean, V.; Ansaldi, M. Insights into the Functions of a Prophage Recombination Directionality Factor. Viruses 2012, 4, 2417–2431. [Google Scholar] [CrossRef] [PubMed]
- Touchon, M.; Bernheim, A.; Rocha, E.P. Genetic and life-history traits associated with the distribution of prophages in bacteria. ISME J. 2016, 10, 2744–2754. [Google Scholar] [CrossRef]
- Paul, J.H. Prophages in marine bacteria: Dangerous molecular time bombs or the key to survival in the seas? ISME J. 2008, 2, 579–589. [Google Scholar] [CrossRef]
- Wendling, C.C.; Goehlich, H.; Roth, O. The structure of temperate phage–bacteria infection networks changes with the phylogenetic distance of the host bacteria. Biol. Lett. 2018, 14, 20180320. [Google Scholar] [CrossRef]
- Castillo, D.; Espejo, R.; Middelboe, M. Genomic structure of bacteriophage 6H and its distribution as prophage in Flavobacterium psychrophilum strains. FEMS Microbiol. Lett. 2014, 351, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Cortes, M.G.; Lin, Y.; Zeng, L.; Balázsi, G. From Bench to Keyboard and Back Again: A Brief History of Lambda Phage Modeling. Annu. Rev. Biophys. 2021, 50, 117–134. [Google Scholar] [CrossRef]
- Casjens, S.R.; Hendrix, R.W. Bacteriophage lambda: Early pioneer and still relevant. Virology 2015, 479–480, 310–330. [Google Scholar] [CrossRef] [PubMed]
- Tsao, Y.-F.; Taylor, V.L.; Kala, S.; Bondy-Denomy, J.; Khan, A.N.; Bona, D.; Cattoir, V.; Lory, S.; Davidson, A.R.; Maxwell, K.L. Phage Morons Play an Important Role in Pseudomonas aeruginosa Phenotypes. J. Bacteriol. 2018, 200, e00189-18. [Google Scholar] [CrossRef] [PubMed]
- Little, J.W.; Michalowski, C.B. Stability and Instability in the Lysogenic State of Phage Lambda. J. Bacteriol. 2010, 192, 6064–6076. [Google Scholar] [CrossRef]
- Węgrzyn, G.; Węgrzyn, A. Genetic Switches During Bacteriophage λ Development. Prog. Nucleic Acid Res. Mol. Biol. 2005, 79, 1–48. [Google Scholar] [PubMed]
- Dragoš, A.; Priyadarshini, B.; Hasan, Z.; Strube, M.L.; Kempen, P.J.; Maróti, G.; Kaspar, C.; Bose, B.; Burton, B.M.; Bischofs, I.B.; et al. Pervasive prophage recombination occurs during evolution of spore-forming Bacilli. ISME J. 2021, 15, 1344–1358. [Google Scholar] [CrossRef]
- Wong, S.; Alattas, H.; Slavcev, R.A. A snapshot of the λ T4rII exclusion (Rex) phenotype in Escherichia coli. Curr. Genet. 2021, 67, 739–745. [Google Scholar] [CrossRef]
- Obeng, N.; Pratama, A.A.; van Elsas, J.D. The Significance of Mutualistic Phages for Bacterial Ecology and Evolution. Trends Microbiol. 2016, 24, 440–449. [Google Scholar] [CrossRef]
- Bloch, S.; Lewandowska, N.; Węgrzyn, G.; Nejman-Faleńczyk, B. Bacteriophages as sources of small non-coding RNA molecules. Plasmid 2021, 113, 102527. [Google Scholar] [CrossRef]
- Fortier, L.-C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef]
- Hsu, B.B.; Way, J.C.; Silver, P.A. Stable Neutralization of a Virulence Factor in Bacteria Using Temperate Phage in the Mammalian Gut. Msystems 2020, 5, e00013-20. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, L.; Yang, D.; Song, J.; Wang, C.; Sun, E.; Gu, C.; Chen, H.; Tong, Y.; Tao, P.; et al. Specific Integration of Temperate Phage Decreases the Pathogenicity of Host Bacteria. Front. Cell. Infect. Microbiol. 2020, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shahin, K.; Soleimani-Delfan, A.; Ding, H.; Wang, H.; Sun, L.; Wang, R. Phage JS02, a putative temperate phage, a novel biofilm-degrading agent for Staphylococcus aureus. Lett. Appl. Microbiol. 2022, 75, 643–654. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, T.; Yu, M.; Chen, Y.-L.; Jin, M. The Life Cycle Transitions of Temperate Phages: Regulating Factors and Potential Ecological Implications. Viruses 2022, 14, 1904. [Google Scholar] [CrossRef] [PubMed]
- Gandon, S. Why Be Temperate: Lessons from Bacteriophage λ. Trends Microbiol. 2016, 24, 356–365. [Google Scholar] [CrossRef]
- Mäntynen, S.; Laanto, E.; Oksanen, H.M.; Poranen, M.M.; Díaz-Muñoz, S.L. Black box of phage–bacterium interactions: Exploring alternative phage infection strategies. Open Biol. 2021, 11, 210188. [Google Scholar] [CrossRef]
- Abedon, S.T. Phage evolution and ecology. Adv. Appl. Microbiol. 2009, 67, 1–45. [Google Scholar] [CrossRef]
- Pires, D.P.; Melo, L.D.; Azeredo, J. Understanding the Complex Phage-Host Interactions in Biofilm Communities. Annu. Rev. Virol. 2021, 8, 73–94. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Márquez, D.L.; Díaz-Orejas, R.; Portillo, F.G.-D. Toxin-antitoxins and bacterial virulence. FEMS Microbiol. Rev. 2016, 40, 592–609. [Google Scholar] [CrossRef] [Green Version]
- Biancalani, T.; Gore, J. Disentangling bacterial invasiveness from lethality in an experimental host-pathogen system. Mol. Syst. Biol. 2019, 15, e8707. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.J., Jr.; Mende, K.; Beckius, M.L.; Akers, K.S.; Romano, D.R.; Wenke, J.C.; Murray, C.K. Biofilm formation by clinical isolates and the implications in chronic infections. BMC Infect. Dis. 2013, 13, 47. [Google Scholar] [CrossRef]
- Gödeke, J.; Paul, K.; Lassak, J.; Thormann, K.M. Phage-induced lysis enhances biofilm formation in Shewanella oneidensis MR-1. ISME J. 2011, 5, 613–626. [Google Scholar] [CrossRef]
- Secor, P.R.; Sweere, J.M.; Michaels, L.A.; Malkovskiy, A.V.; Lazzareschi, D.; Katznelson, E.; Rajadas, J.; Birnbaum, M.E.; Arrigoni, A.; Braun, K.R.; et al. Filamentous Bacteriophage Promote Biofilm Assembly and Function. Cell Host Microbe 2015, 18, 549–559. [Google Scholar] [CrossRef]
- Chhibber, S.; Nag, D.; Bansal, S. Inhibiting biofilm formation by Klebsiella pneumoniae B5055 using an iron antagonizing molecule and a bacteriophage. BMC Microbiol. 2013, 13, 174. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wilksch, J.J.; Strugnell, R.A.; Gee, M.L. Role of Capsular Polysaccharides in Biofilm Formation: An AFM Nanomechanics Study. ACS Appl. Mater. Interfaces 2015, 7, 13007–13013. [Google Scholar] [CrossRef] [PubMed]
- Van Houdt, R.; Michiels, C. Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl. Microbiol. 2010, 109, 1117–1131. [Google Scholar] [CrossRef] [PubMed]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363, fnw002. [Google Scholar] [CrossRef]
- Sun, J.; Geng, P.; Wan, X.; Yuan, Z.; Xiong, H.; Hu, X. [Advances of phage receptor binding proteins]. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2021, 37, 2614–2622. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Gao, D.; Ji, H.; Wang, L.; Li, X.; Hu, D.; Zhao, J.; Wang, S.; Tao, P.; Li, X.; Qian, P. Fitness Trade-Offs in Phage Cocktail-Resistant Salmonella enterica Serovar Enteritidis Results in Increased Antibiotic Susceptibility and Reduced Virulence. Microbiol. Spectr. 2022, 10, e0291422. [Google Scholar] [CrossRef] [PubMed]
- de Jonge, P.A.; Nobrega, F.L.; Brouns, S.J.; Dutilh, B.E. Molecular and Evolutionary Determinants of Bacteriophage Host Range. Trends Microbiol. 2019, 27, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Dunne, M.; Prokhorov, N.S.; Loessner, M.J.; Leiman, P.G. Reprogramming bacteriophage host range: Design principles and strategies for engineering receptor binding proteins. Curr. Opin. Biotechnol. 2021, 68, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liu, Z.; Zhang, Q.; Liu, Y.; Chen, Y. Phage Engineering for Targeted Multidrug-Resistant Escherichia coli. Int. J. Mol. Sci. 2023, 24, 2459. [Google Scholar] [CrossRef]
- Li, X.; Chen, Y.; Wang, S.; Duan, X.; Zhang, F.; Guo, A.; Tao, P.; Chen, H.; Li, X.; Qian, P. Exploring the Benefits of Metal Ions in Phage Cocktail for the Treatment of Methicillin-Resistant Staphylococcus aureus (MRSA) Infection. Infect. Drug Resist. 2022, 15, 2689–2702. [Google Scholar] [CrossRef]
- Lood, C.; Haas, P.-J.; van Noort, V.; Lavigne, R. Shopping for phages? Unpacking design rules for therapeutic phage cocktails. Curr. Opin. Virol. 2022, 52, 236–243. [Google Scholar] [CrossRef]
- Mapes, A.C.; Trautner, B.W.; Liao, K.S.; Ramig, R.F. Development of expanded host range phage active on biofilms of multi-drug resistant Pseudomonas aeruginosa. Bacteriophage 2016, 6, e1096995. [Google Scholar] [CrossRef]
- Fischer, S.; Kittler, S.; Klein, G.; Glünder, G. Impact of a Single Phage and a Phage Cocktail Application in Broilers on Reduction of Campylobacter jejuni and Development of Resistance. PLoS ONE 2013, 8, e78543. [Google Scholar] [CrossRef]
- Khaleel, T.; Younger, E.; McEwan, A.R.; Varghese, A.S.; Smith, M.C.M. A phage protein that binds φC31 integrase to switch its directionality. Mol. Microbiol. 2011, 80, 1450–1463. [Google Scholar] [CrossRef]
- Ravin, N. Replication and Maintenance of Linear Phage-Plasmid N15. Microbiol. Spectr. 2015, 3, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Pfeifer, E.; Sousa, J.A.M.D.; Touchon, M.; Rocha, E.P.C. Bacteria have numerous distinctive groups of phage–plasmids with conserved phage and variable plasmid gene repertoires. Nucleic Acids Res. 2021, 49, 2655–2673. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, K.; Yuan, P.; Perry, K.; Sharp, R.; Van Duyne, G.D. Attachment site recognition and regulation of directionality by the serine integrases. Nucleic Acids Res. 2013, 41, 8341–8356. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich, L.; Sigal, N.; Borovok, I.; Nir-Paz, R.; Herskovits, A.A. Prophage Excision Activates Listeria Competence Genes that Promote Phagosomal Escape and Virulence. Cell 2012, 150, 792–802. [Google Scholar] [CrossRef]
- Fang, Y.; Mercer, R.G.; McMullen, L.M.; Gänzle, M.G. Induction of Shiga Toxin-Encoding Prophage by Abiotic Environmental Stress in Food. Appl. Environ. Microbiol. 2017, 83, e01378-17. [Google Scholar] [CrossRef]
- In Proceedings of theChang, Y.; Bai, J.; Lee, J.-H.; Ryu, S. Mutation of a Staphylococcus aureus temperate bacteriophage to a virulent one and evaluation of its application. Food Microbiol. 2019, 82, 523–532. [Google Scholar] [CrossRef]
- Nepal, R.; Houtak, G.; Shaghayegh, G.; Bouras, G.; Shearwin, K.; Psaltis, A.J.; Wormald, P.-J.; Vreugde, S. Prophages encoding human immune evasion cluster genes are enriched in Staphylococcus aureus isolated from chronic rhinosinusitis patients with nasal polyps. Microb. Genom. 2021, 7, 000726. [Google Scholar] [CrossRef] [PubMed]
- Bondy-Denomy, J.; Qian, J.; Westra, E.R.; Buckling, A.; Guttman, D.S.; Davidson, A.R.; Maxwell, K.L. Prophages mediate defense against phage infection through diverse mechanisms. ISME J. 2016, 10, 2854–2866. [Google Scholar] [CrossRef]
- Fillol-Salom, A.; Rostøl, J.T.; Ojiogu, A.D.; Chen, J.; Douce, G.; Humphrey, S.; Penadés, J.R. Bacteriophages benefit from mobilizing pathogenicity islands encoding immune systems against competitors. Cell 2022, 185, 3248–3262.e20. [Google Scholar] [CrossRef]
- Azulay, G.; Pasechnek, A.; Stadnyuk, O.; Ran-Sapir, S.; Fleisacher, A.M.; Borovok, I.; Sigal, N.; Herskovits, A.A. A dual-function phage regulator controls the response of cohabiting phage elements via regulation of the bacterial SOS response. Cell Rep. 2022, 39, 110723. [Google Scholar] [CrossRef]
- Hu, J.; Ye, H.; Wang, S.; Wang, J.; Han, D. Prophage Activation in the Intestine: Insights Into Functions and Possible Applications. Front. Microbiol. 2021, 12, 785634. [Google Scholar] [CrossRef]
- Jones, E.C.; Uphoff, S. Single-molecule imaging of LexA degradation in Escherichia coli elucidates regulatory mechanisms and heterogeneity of the SOS response. Nat. Microbiol. 2021, 6, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Jancheva, M.; Böttcher, T. A Metabolite of Pseudomonas Triggers Prophage-Selective Lysogenic to Lytic Conversion in Staphylococcus aureus. J. Am. Chem. Soc. 2021, 143, 8344–8351. [Google Scholar] [CrossRef] [PubMed]
- Campoy, S.; Hervàs, A.; Busquets, N.; Erill, I.; Teixidó, L.; Barbé, J. Induction of the SOS response by bacteriophage lytic development in Salmonella enterica. Virology 2006, 351, 360–367. [Google Scholar] [CrossRef]
- Ghosh, D.; Roy, K.; Williamson, K.E.; Srinivasiah, S.; Wommack, K.E.; Radosevich, M. Acyl-Homoserine Lactones Can Induce Virus Production in Lysogenic Bacteria: An Alternative Paradigm for Prophage Induction. Appl. Environ. Microbiol. 2009, 75, 7142–7152. [Google Scholar] [CrossRef] [PubMed]
- Fillol-Salom, A.; Alsaadi, A.; de Sousa, J.A.M.; Zhong, L.; Foster, K.R.; Rocha, E.P.C.; Penadés, J.R.; Ingmer, H.; Haaber, J. Bacteriophages benefit from generalized transduction. PLOS Pathog. 2019, 15, e1007888. [Google Scholar] [CrossRef]
- Colavecchio, A.; Cadieux, B.; Lo, A.; Goodridge, L.D. Bacteriophages Contribute to the Spread of Antibiotic Resistance Genes among Foodborne Pathogens of the Enterobacteriaceae Family—A Review. Front. Microbiol. 2017, 8, 1108. [Google Scholar] [CrossRef]
- Sternberg, N. The production of generalized transducing phage by bacteriophage lambda. Gene 1986, 50, 69–85. [Google Scholar] [CrossRef]
- Kwoh, D.Y.; Kemper, J. Bacteriophage P22-mediated specialized transduction in Salmonella typhimurium: Identification of different types of specialized transducing particles. J. Virol. 1978, 27, 535–550. [Google Scholar] [CrossRef]
- Humphrey, S.; Fillol-Salom, A.; Quiles-Puchalt, N.; Ibarra-Chávez, R.; Haag, A.F.; Chen, J.; Penadés, J.R. Bacterial chromosomal mobility via lateral transduction exceeds that of classical mobile genetic elements. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Catalao, M.J.; Gil, F.; Moniz-Pereira, J.; São-José, C.; Pimentel, M. Diversity in bacterial lysis systems: Bacteriophages show the way. FEMS Microbiol. Rev. 2013, 37, 554–571. [Google Scholar] [CrossRef] [Green Version]
- Secor, P.R.; Burgener, E.B.; Kinnersley, M.; Jennings, L.K.; Roman-Cruz, V.; Popescu, M.; Van Belleghem, J.D.; Haddock, N.; Copeland, C.; Michaels, L.A.; et al. Pf Bacteriophage and Their Impact on Pseudomonas Virulence, Mammalian Immunity, and Chronic Infections. Front. Immunol. 2020, 11, 244. [Google Scholar] [CrossRef]
- Rice, S.A.; Tan, C.H.; Mikkelsen, P.J.; Kung, V.; Woo, J.; Tay, M.; Hauser, A.; McDougald, D.; Webb, J.; Kjelleberg, S. The biofilm life cycle and virulence of Pseudomonas aeruginosa are dependent on a filamentous prophage. ISME J. 2009, 3, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, S.; Sun, E.; Chen, Y.; Hua, L.; Wang, X.; Zhou, R.; Chen, H.; Peng, Z.; Wu, B. A temperate Siphoviridae bacteriophage isolate from Siberian tiger enhances the virulence of methicillin-resistant Staphylococcus aureus through distinct mechanisms. Virulence 2022, 13, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Clooney, A.G.; Sutton, T.D.; Shkoporov, A.N.; Holohan, R.K.; Daly, K.M.; O’Regan, O.; Ryan, F.J.; Draper, L.A.; Plevy, S.E.; Ross, R.P.; et al. Whole-Virome Analysis Sheds Light on Viral Dark Matter in Inflammatory Bowel Disease. Cell Host Microbe 2019, 26, 764–778.e5. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.-W.; Adams, J.B.; Gregory, A.C.; Borody, T.; Chittick, L.; Fasano, A.; Khoruts, A.; Geis, E.; Maldonado, J.; McDonough-Means, S.; et al. Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: An open-label study. Microbiome 2017, 5, 10. [Google Scholar] [CrossRef]
- Cai, M.; Pu, B.; Wang, Y.; Lv, L.; Jiang, C.; Fu, X.; Zhang, Y.; Zhao, W.; Dong, K.; Yang, Y.; et al. A Plasmid With Conserved Phage Genes Helps Klebsiella pneumoniae Defend Against the Invasion of Transferable DNA Elements at the Cost of Reduced Virulence. Front. Microbiol. 2022, 13, 827545. [Google Scholar] [CrossRef]
- Sumrall, E.T.; Shen, Y.; Keller, A.P.; Rismondo, J.; Pavlou, M.; Eugster, M.R.; Boulos, S.; Disson, O.; Thouvenot, P.; Kilcher, S.; et al. Phage resistance at the cost of virulence: Listeria monocytogenes serovar 4b requires galactosylated teichoic acids for InlB-mediated invasion. PLOS Pathog. 2019, 15, e1008032. [Google Scholar] [CrossRef]
- Roszniowski, B.; Latka, A.; Maciejewska, B.; Vandenheuvel, D.; Olszak, T.; Briers, Y.; Holt, G.S.; Valvano, M.A.; Lavigne, R.; Smith, D.L.; et al. The temperate Burkholderia phage AP3 of the Peduovirinae shows efficient antimicrobial activity against B. cenocepacia of the IIIA lineage. Appl. Microbiol. Biotechnol. 2017, 101, 1203–1216. [Google Scholar] [CrossRef]
- Bhattarai, S.R.; Yoo, S.Y.; Lee, S.-W.; Dean, D. Engineered phage-based therapeutic materials inhibit Chlamydia trachomatis intracellular infection. Biomaterials 2012, 33, 5166–5174. [Google Scholar] [CrossRef]
- Jin, M.; Chen, J.; Zhao, X.; Hu, G.; Wang, H.; Liu, Z.; Chen, W.-H. An Engineered λ Phage Enables Enhanced and Strain-Specific Killing of Enterohemorrhagic Escherichia coli. Microbiol. Spectr. 2022, 10, e0127122. [Google Scholar] [CrossRef]
- Al-Anany, A.M.; Fatima, R.; Hynes, A.P. Temperate phage-antibiotic synergy eradicates bacteria through depletion of lysogens. Cell Rep. 2021, 35, 109172. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Friedman, N.; Molshanski-Mor, S.; Qimron, U. Reversing Bacterial Resistance to Antibiotics by Phage-Mediated Delivery of Dominant Sensitive Genes. Appl. Environ. Microbiol. 2012, 78, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Jepson, C.D.; March, J.B. Bacteriophage lambda is a highly stable DNA vaccine delivery vehicle. Vaccine 2004, 22, 2413–2419. [Google Scholar] [CrossRef] [PubMed]
- Gamage, L.N.; Ellis, J.; Hayes, S. Immunogenicity of bacteriophage lambda particles displaying porcine Circovirus 2 (PCV2) capsid protein epitopes. Vaccine 2009, 27, 6595–6604. [Google Scholar] [CrossRef]
- Fosså, A.; Alsøe, L.; Crameri, R.; Funderud, S.; Gaudernack, G.; Smeland, E.B. Serological cloning of cancer/testis antigens expressed in prostate cancer using cDNA phage surface display. Cancer Immunol. Immunother. 2004, 53, 431–438. [Google Scholar] [CrossRef]
- Hashemi, H.; Bamdad, T.; Jamali, A.; Pouyanfard, S.; Mohammadi, M.G. Evaluation of humoral and cellular immune responses against HSV-1 using genetic immunization by filamentous phage particles: A comparative approach to conventional DNA vaccine. J. Virol. Methods 2010, 163, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Catala, A.; Dzieciatkowska, M.; Wang, G.; Gutierrez-Hartmann, A.; Simberg, D.; Hansen, K.C.; D’Alessandro, A.; Catalano, C.E. Targeted Intracellular Delivery of Trastuzumab Using Designer Phage Lambda Nanoparticles Alters Cellular Programs in Human Breast Cancer Cells. ACS Nano 2021, 15, 11789–11805. [Google Scholar] [CrossRef]
- Shah, M.; Taylor, V.L.; Bona, D.; Tsao, Y.; Stanley, S.Y.; Pimentel-Elardo, S.M.; McCallum, M.; Bondy-Denomy, J.; Howell, P.L.; Nodwell, J.R.; et al. A phage-encoded anti-activator inhibits quorum sensing in Pseudomonas aeruginosa. Mol. Cell 2021, 81, 571–583.e6. [Google Scholar] [CrossRef]
- Olszak, T.; Shneider, M.M.; Latka, A.; Maciejewska, B.; Browning, C.; Sycheva, L.V.; Cornelissen, A.; Danis-Wlodarczyk, K.; Senchenkova, S.N.; Shashkov, A.S.; et al. The O-specific polysaccharide lyase from the phage LKA1 tailspike reduces Pseudomonas virulence. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Zhang, H.; Fouts, D.E.; DePew, J.; Stevens, R.H. Genetic modifications to temperate Enterococcus faecalis phage ϕEf11 that abolish the establishment of lysogeny and sensitivity to repressor, and increase host range and productivity of lytic infection. Microbiology 2013, 159 Pt 6, 1023–1035. [Google Scholar] [CrossRef] [Green Version]
- Arroyave, E.; Hyseni, I.; Burkhardt, N.; Kuo, Y.-F.; Wang, T.; Munderloh, U.; Fang, R. Rickettsia parkeri with a Genetically Disrupted Phage Integrase Gene Exhibits Attenuated Virulence and Induces Protective Immunity against Fatal Rickettsioses in Mice. Pathogens 2021, 10, 819. [Google Scholar] [CrossRef]
- Landlinger, C.; Tisakova, L.; Oberbauer, V.; Schwebs, T.; Muhammad, A.; Latka, A.; Van Simaey, L.; Vaneechoutte, M.; Guschin, A.; Resch, G.; et al. Engineered Phage Endolysin Eliminates Gardnerella Biofilm Without Damaging Beneficial Bacteria in Bacterial Vaginosis Ex Vivo. Pathogens 2021, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Payaslian, F.; Gradaschi, V.; Piuri, M. Genetic manipulation of phages for therapy using BRED. Curr. Opin. Biotechnol. 2021, 68, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Nunez, C.; Kostoulias, X.; Peleg, A.; Short, F.; Qu, Y. A comprehensive comparison of biofilm formation and capsule production for bacterial survival on hospital surfaces. Biofilm 2017, 5, 100105. [Google Scholar] [CrossRef] [PubMed]
- Howard-Varona, C.; Hargreaves, K.R.; Abedon, S.T.; Sullivan, M.B. Lysogeny in nature: Mechanisms, impact and ecology of temperate phages. ISME J. 2017, 11, 1511–1520. [Google Scholar] [CrossRef]
- Łobocka, M.; Dąbrowska, K.; Górski, A. Engineered Bacteriophage Therapeutics: Rationale, Challenges and Future. Biodrugs 2021, 35, 255–280. [Google Scholar] [CrossRef]
- Diard, M.; Hardt, W.-D. Evolution of bacterial virulence. FEMS Microbiol. Rev. 2017, 41, 679. [Google Scholar] [CrossRef]
- Schubert, W.-D.; Urbanke, C.; Ziehm, T.; Beier, V.; Machner, M.P.; Domann, E.; Wehland, J.; Chakraborty, T.; Heinz, D.W. Structure of Internalin, a Major Invasion Protein of Listeria monocytogenes, in Complex with Its Human Receptor E-Cadherin. Cell 2002, 111, 825–836. [Google Scholar] [CrossRef]
- Markwitz, P.; Olszak, T.; Gula, G.; Kowalska, M.; Arabski, M.; Drulis-Kawa, Z. Emerging Phage Resistance in Pseudomonas aeruginosa PAO1 Is Accompanied by an Enhanced Heterogeneity and Reduced Virulence. Viruses 2021, 13, 1332. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, H.; Zhou, Y.; Zhu, S.; Pang, M.; Wang, Y.; Wang, J.; Olaniran, A.; Xiao, Y.; Schmidt, S.; et al. Dysbiosis and intestinal inflammation caused by Salmonella Typhimurium in mice can be alleviated by preadministration of a lytic phage. Microbiol. Res. 2022, 260, 127020. [Google Scholar] [CrossRef]
- Hathroubi, S.; Mekni, M.A.; Domenico, P.; Nguyen, D.; Jacques, M. Biofilms: Microbial Shelters Against Antibiotics. Microb. Drug Resist. 2017, 23, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Tagliaferri, T.L.; Jansen, M.; Horz, H.-P. Fighting Pathogenic Bacteria on Two Fronts: Phages and Antibiotics as Combined Strategy. Front. Cell. Infect. Microbiol. 2019, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, Y.; Wang, Z.; Wei, J.; Hu, T.; Si, J.; Tao, G.; Zhang, L.; Xie, L.; Abdalla, A.E.; et al. A combination therapy of Phages and Antibiotics: Two is better than one. Int. J. Biol. Sci. 2021, 17, 3573–3582. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Green, S.I.; Min, L.; Clark, J.R.; Salazar, K.C.; Terwilliger, A.L.; Kaplan, H.B.; Trautner, B.W.; Ramig, R.F.; Maresso, A.W. Phage-Antibiotic Synergy Is Driven by a Unique Combination of Antibacterial Mechanism of Action and Stoichiometry. mBio 2020, 11, e01462-20. [Google Scholar] [CrossRef]
- Rhodes, K.A.; Schweizer, H.P. Antibiotic resistance in Burkholderia species. Drug Resist. Updat. 2016, 28, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Endres, T.M.; Konstan, M.W. What Is Cystic Fibrosis? Jama 2022, 327, 191. [Google Scholar] [CrossRef]
- Elborn, J.S. Cystic fibrosis. Lancet (London, England) 2016, 388, 2519–2531. [Google Scholar] [CrossRef]
- Shteinberg, M.; Haq, I.J.; Polineni, D.; Davies, J.C. Cystic fibrosis. Lancet (London, England) 2021, 397, 2195–2211. [Google Scholar] [CrossRef]
- Tariq, M.A.; Everest, F.L.C.; Cowley, L.A.; Wright, R.; Holt, G.S.; Ingram, H.; Duignan, L.A.M.; Nelson, A.; Lanyon, C.V.; Perry, A.; et al. Temperate Bacteriophages from Chronic Pseudomonas aeruginosa Lung Infections Show Disease-Specific Changes in Host Range and Modulate Antimicrobial Susceptibility. Msystems 2019, 4, e00191-18. [Google Scholar] [CrossRef]
- Łusiak-Szelachowska, M.; Międzybrodzki, R.; Drulis-Kawa, Z.; Cater, K.; Knežević, P.; Winogradow, C.; Amaro, K.; Jończyk-Matysiak, E.; Weber-Dąbrowska, B.; Rękas, J.; et al. Bacteriophages and antibiotic interactions in clinical practice: What we have learned so far. J. Biomed. Sci. 2022, 29, 1–17. [Google Scholar] [CrossRef]
- Peng, H.; Chen, I.A. Phage engineering and the evolutionary arms race. Curr. Opin. Biotechnol. 2021, 68, 23–29. [Google Scholar] [CrossRef] [PubMed]
- de Vries, C.R.; Chen, Q.; Demirdjian, S.; Kaber, G.; Khosravi, A.; Liu, D.; Van Belleghem, J.D.; Bollyky, P.L. Phages in vaccine design and immunity; mechanisms and mysteries. Curr. Opin. Biotechnol. 2021, 68, 160–165. [Google Scholar] [CrossRef]
- Iwagami, Y.; Casulli, S.; Nagaoka, K.; Kim, M.; Carlson, R.I.; Ogawa, K.; Lebowitz, M.S.; Fuller, S.; Biswas, B.; Stewart, S.; et al. Lambda phage-based vaccine induces antitumor immunity in hepatocellular carcinoma. Heliyon 2017, 3, e00407. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.U.; Wang, W.; Sun, Q.; Shah, J.A.; Li, C.; Sun, Y.; Li, Y.; Zhang, B.; Chen, W.; Wang, S. Endolysin, a Promising Solution against Antimicrobial Resistance. Antibiotics 2021, 10, 1277. [Google Scholar] [CrossRef] [PubMed]
- Warner, C.M.; Barker, N.; Lee, S.-W.; Perkins, E.J. M13 bacteriophage production for large-scale applications. Bioprocess Biosyst. Eng. 2014, 37, 2067–2072. [Google Scholar] [CrossRef]
- Chavda, V.P.; Pandya, A.; Kypreos, E.; Patravale, V.; Apostolopoulos, V. Chlamydia trachomatis: Quest for an eye-opening vaccine breakthrough. Expert Rev. Vaccines 2022, 21, 771–781. [Google Scholar] [CrossRef]
- Carter, M.Q.; Laniohan, N.; Pham, A.; Quiñones, B. Comparative genomic and phenotypic analyses of the virulence potential in Shiga toxin-producing Escherichia coli O121:H7 and O121:H10. Front. Cell. Infect. Microbiol. 2022, 12, 1043726. [Google Scholar] [CrossRef]
- Chibani-Chennoufi, S.; Sidoti, J.; Bruttin, A.; Kutter, E.; Sarker, S.; Brüssow, H. In Vitro and In Vivo Bacteriolytic Activities of Escherichia coli Phages: Implications for Phage Therapy. Antimicrob. Agents Chemother. 2004, 48, 2558–2569. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR–Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef]
- Czarnek, M.; Bereta, J. The CRISPR-Cas system—from bacterial immunity to genome engineering. Postep. Hig. I Med. Dosw. (Online) 2016, 70, 901–916. [Google Scholar] [CrossRef]
- Rollie, C.; Chevallereau, A.; Watson, B.N.J.; Chyou, T.-Y.; Fradet, O.; McLeod, I.; Fineran, P.C.; Brown, C.M.; Gandon, S.; Westra, E.R. Targeting of temperate phages drives loss of type I CRISPR–Cas systems. Nature 2020, 578, 149–153. [Google Scholar] [CrossRef]
- Vink, J.N.; Martens, K.J.; Vlot, M.; McKenzie, R.E.; Almendros, C.; Bonilla, B.E.; Brocken, D.J.; Hohlbein, J.; Brouns, S.J. Direct Visualization of Native CRISPR Target Search in Live Bacteria Reveals Cascade DNA Surveillance Mechanism. Mol. Cell 2020, 77, 39–50.e10. [Google Scholar] [CrossRef]
- Bruger, E.L.; Snyder, D.J.; Cooper, V.S.; Waters, C.M. Quorum sensing provides a molecular mechanism for evolution to tune and maintain investment in cooperation. ISME J. 2021, 15, 1236–1247. [Google Scholar] [CrossRef]
- Jugder, B.-E.; Batista, J.H.; Gibson, J.A.; Cunningham, P.M.; Asara, J.M.; Watnick, P.I. Vibrio cholerae high cell density quorum sensing activates the host intestinal innate immune response. Cell Rep. 2022, 40, 111368. [Google Scholar] [CrossRef]
- Silpe, J.E.; Bassler, B.L. A Host-Produced Quorum-Sensing Autoinducer Controls a Phage Lysis-Lysogeny Decision. Cell 2019, 176, 268–280.e13. [Google Scholar] [CrossRef]
- Amankwah, S.; Abdusemed, K.; Kassa, T. Bacterial Biofilm Destruction: A Focused Review On The Recent Use of Phage-Based Strategies With Other Antibiofilm Agents. Nanotechnol. Sci. Appl. 2021, 14, 161–177. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Phage | Host | Function | Description | Reference |
|---|---|---|---|---|
| PHB09 | B. bronchiseptica | Virulence shrinks under massive phage predation | PHB09 inserted and thus disrupted pilin protein gene, but the vaccine made of lysogenic B. bronchiseptica strain Bb01+ also showed effective protection of mice challenged with virulent B. bronchiseptica. | [42] |
| p2 | K. pneumoniae | Host prevents invasion by reducing virulence | The presence of a plasmid form of prophage can provide host bacterium with resistance to other foreign DNA at the cost of the host virulence. | [96] |
| ΔLCRA500 | L.monocytogenes | Host prevents invasion by reducing virulence | The temperate phage ΔLCRA500, which has been knocked out of the gp32, gp33 and integrase genes, has marked lytic ability and a specific Listeria serotype 4b host range. | [97] |
| PHB22a, PHB25a, PHB38a, and PHB40a | Methicillin-Resistant S. aureus S-18 | Temperate phage cocktails enhanced with ions | The antibacterial effect of this recipe is determined by the biofilm removal efficiency, where added ions proved higher bacterial CFU reduction ability. Moreover, using G. mellonella larvae as animal model against MRSA S-18 infection, the survival rate resulting from ions–phages therapy is 10% higher. | [65] |
| SA13m | S.aureus | Converted into stable lytic phage | A virulent mutant SA13m obtained through random deletion of temperate phage SA13 exhibits active lytic activity and no sign of lysogenicity. Application of SA13m in sterilized milk showed that S. aureus was reduced to non-detectable levels, suggesting that SA13m can efficiently control the growth of S. aureus in food. | [75] |
| AP3 | B.cenocepacia | Combined with antibiotics | Temperate Burkholderia phage AP3 combined with antibiotics demonstrates increased bactericidal effects in in vivo experiments with moth larvae. | [98] |
| M13 | C. trachomatis | Temperate phage display | Compared to C. trachomatis infection alone, engineered phages stably express RGD motifs and C. trachomatis peptides and significantly reduce C. trachomatis infection in HeLa and primary cervical cells. | [99] |
| 933W | E. coli | Modification of phage genes to inhibit toxin production | The phage demonstrated superior toxin inhibition in both in vivo and in vitro infections. In the foodborne pathogen EHEC, the λ prophage 933W both produces Stx2 and inhibits phage overlap infection of other λ phages. | [41] |
| Eλ | E. coli | Gene-modified phage with CRISPR-Cas3 system | A genetically engineered λ phage exhibits enhanced killing ability and host specificity when incorporated with CRISPR-Cas3 system and knockdown of the lytic gene cro. This engineered phage specifically and effectively eliminates enterohemorrhagic E. coli infection and validated the superior performance over wild-type phages through in vitro and in vivo experiments. In addition, there is no evidence in this study showing that EHEC developed resistance to engineered lambda phage. | [100] |
| HK97 | E. coli | Combined with antibiotics | In vitro bacterial eradication is observed after coadministration of E. coli temperate phage HK97 and antibiotic ciprofloxacin. This synergy works in line with the depletion of lysogens which ciprofloxacin specially targets. | [101] |
| λ | E. coli | Combined with antibiotics | The restoration of antibiotic sensitivity to two antibiotics, streptomycin and nalidixic acid, can be realized by the introduction of specific genes rpsL and gyrA, respectively, in the process of temperate phage lysogenization. | [102] |
| λ | E. coli | Phage vaccine | Recombined with targeted DNA, phage λ can carry the particulate DNA into human system and become protected from degradation, making sure the antigen presenting cells can recognize and capture them. | [103] |
| λ | E. coli | Phage vaccine | A vaccine made from temperate phage λ using phage display technique showed significant efficiency in eliciting anti-PCV2 immune response after the first vaccination without adjuvant. | [104] |
| M13 | E. coli | Phage vaccine | Using temperate phage M13 surface display, the diverse clone of tumor-associated antigens in prostate cancer is achieved and makes it a desirable candidate for vaccine development in prostate cancer. | [105] |
| Filamentous phage | E. coli | Phage vaccine | The filamentous phage inoculation induced both humoral and cellular immune response against HSV-1 in BALB/c mice. | [106] |
| λ | E. coli | Lambda PLP | Phage-like particles (PLPs) are derived from phage lambda, and robust internalization of Trz PLPs resulted in increased intracellular Trz concentrations, prolonged cell growth inhibition and regulation of cellular programs associated with HER2 signaling, proliferation, metabolism and protein synthesis compared to Trz treatment. | [107] |
| λ | E. coli | Reverse antibiotic sensitivity | Using lysogenic conversion, a sensitivity cassette is brought into the bacteria genome and unwanted recombination is managed to be avoided. | [102] |
| DMS3 | P.aeruginosa | Encode proteins that block QS system | P. aeruginosa phage DMS3 can protect bacteria from attack by other phages by inhibiting bacterial quorum sensing. DMS3 encodes a QS anti-activator protein aqs1 that is expressed immediately after phage infection. aqs1 inhibits the activity of LasR, a major regulator of quorum sensing, and restrains twitching motility and superinfection. | [108] |
| LKA1 | P.aeruginosa | Lyase production to eradicate biofilm | A temperate phage of Pseudomonas has been proved to be able to produce a lyase, LKA1gp49, to degrade LPS. LKA1gp49 lyase efficiently reduces P. aeruginosa virulence in the in vivo G. mellonella infection model, and sensitizes bacterial cells to the lytic activity of serum complement. | [109] |
| Ef11 | E. faecalis | Converted into stable lytic phage | By deletion of putative lysogeny gene module and replacement of putative cro promoter from the recombinant phage genome with a 50 nisin-inducible promoter, the temperate phage is rendered virulent and with expanded host range. | [110] |
| 3A2 | R. parkeri | Gene insertion led to attenuated phenotype | The R. parkeri mutant strain is genetically modified by inserting a transposon into the gene encoding the phage integrase in the bacterial genome. Such a mutant exhibited significantly reduced virulence, significantly smaller phage plaques and improved histopathological alterations in intravenously infected mice compared to the parental wild type. | [111] |
| Gardnerella phage | G. vaginalis | Engineered endolysins | A genetically modified endolysin PM-477 produced by Gardnerella phage exhibits the ability to completely disrupt bacterial biofilms of G. ardnerella vaginalis and has no effect on beneficial Lactobacillus or other species of vaginal bacteria. | [112] |
| ZoeJ and BPs | M. abscessus | Converted into stable lytic phage | Two temperate phages are transformed into lytic phages and made into a three-phage cocktail along with one lytic phage. The cocktail is administered to a cystic fibrosis patient and recovering signs are observed after six months’ treatment. | [113] |
| BP96115 | Salmonella | Virulence shrinks under massive phage invasion | As opposed to the streptomycin treatment, pre-treatment of mice with temperate phage safeguarded a stable and more diverse gut ecosystem and protected the intestinal system of mice against the pathogen challenge. | [114] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Liu, Z.; Song, J.; Chen, Y. Disarm The Bacteria: What Temperate Phages Can Do. Curr. Issues Mol. Biol. 2023, 45, 1149-1167. https://doi.org/10.3390/cimb45020076
Zhou S, Liu Z, Song J, Chen Y. Disarm The Bacteria: What Temperate Phages Can Do. Current Issues in Molecular Biology. 2023; 45(2):1149-1167. https://doi.org/10.3390/cimb45020076
Chicago/Turabian StyleZhou, Shiyue, Zhengjie Liu, Jiaoyang Song, and Yibao Chen. 2023. "Disarm The Bacteria: What Temperate Phages Can Do" Current Issues in Molecular Biology 45, no. 2: 1149-1167. https://doi.org/10.3390/cimb45020076
APA StyleZhou, S., Liu, Z., Song, J., & Chen, Y. (2023). Disarm The Bacteria: What Temperate Phages Can Do. Current Issues in Molecular Biology, 45(2), 1149-1167. https://doi.org/10.3390/cimb45020076