

Cloning of Three Cytokinin Oxidase/Dehydrogenase Genes in Bambusa oldhamii


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Reagents
2.3. Total RNA Extraction, cDNA Synthesis, and DIG Labeled Probe Preparation
2.4. Genomic Library Screening, Phage DNA Preparation, and DNA Sequencing by Chromosome Walking
2.5. Bioinformatics and Promoter Analysis
3. Results
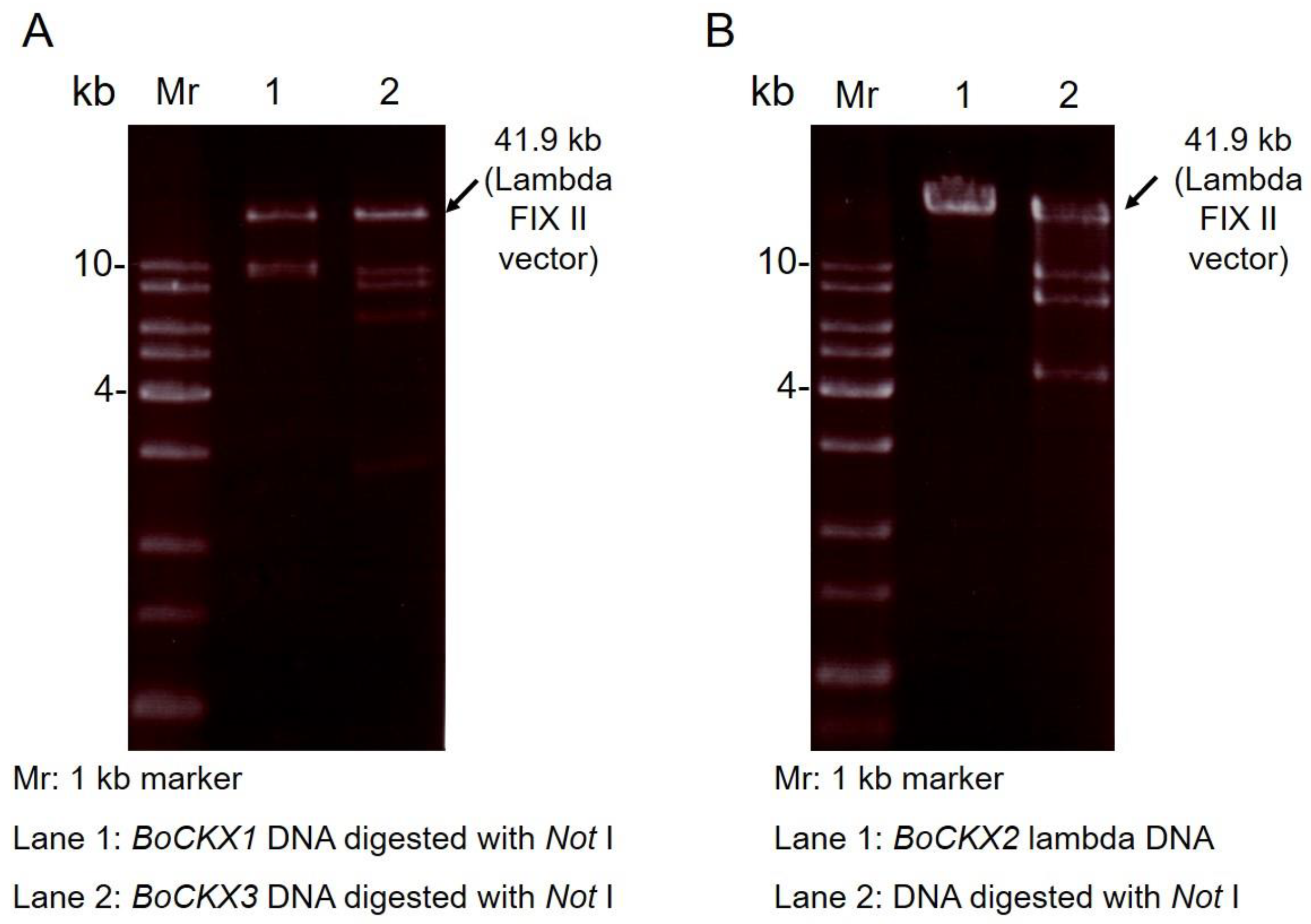
3.1. Cloning of Three BoCKXs Genes by Screening a Bamboo Genomic DNA Library
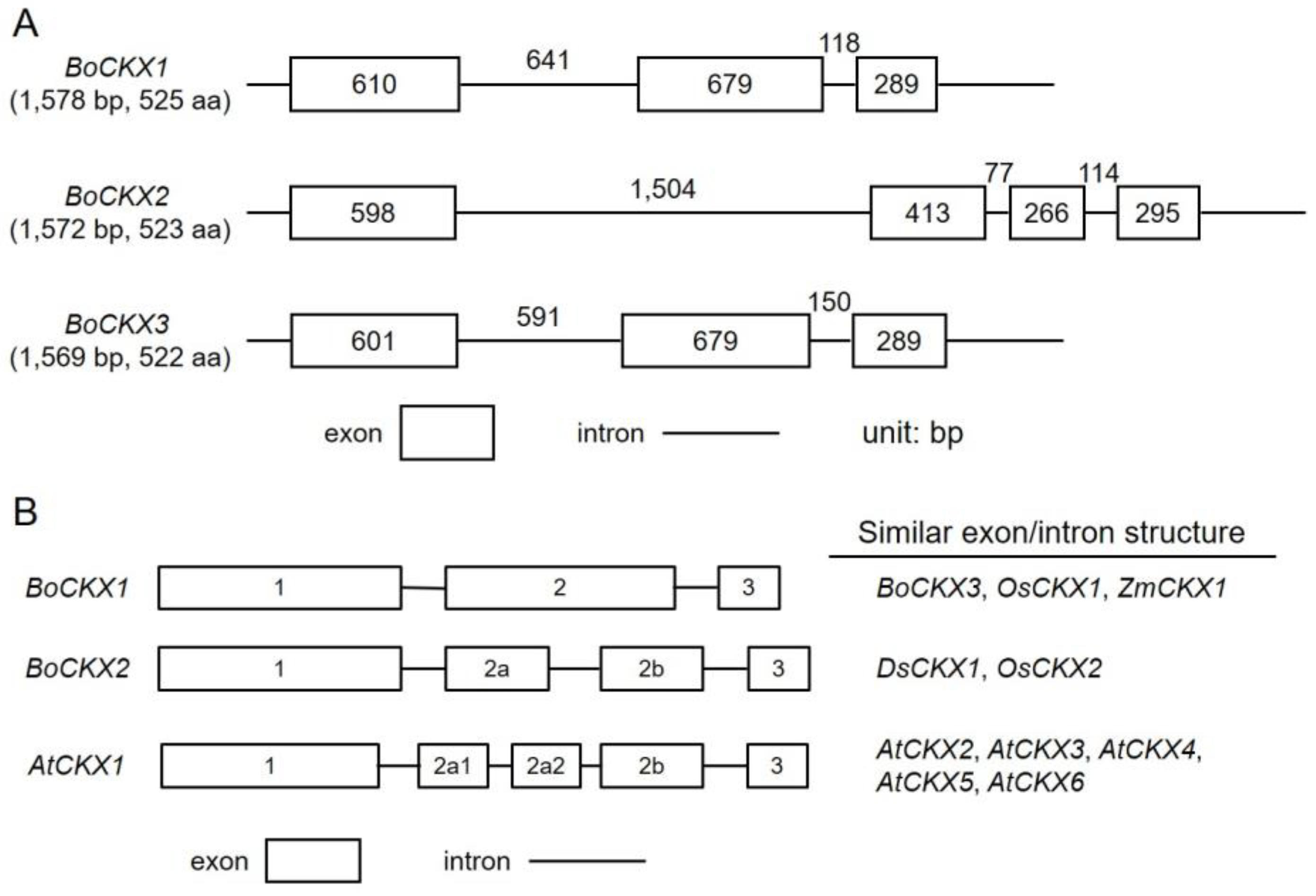
3.2. Genomic Organization of the Three BoCKXs Genes
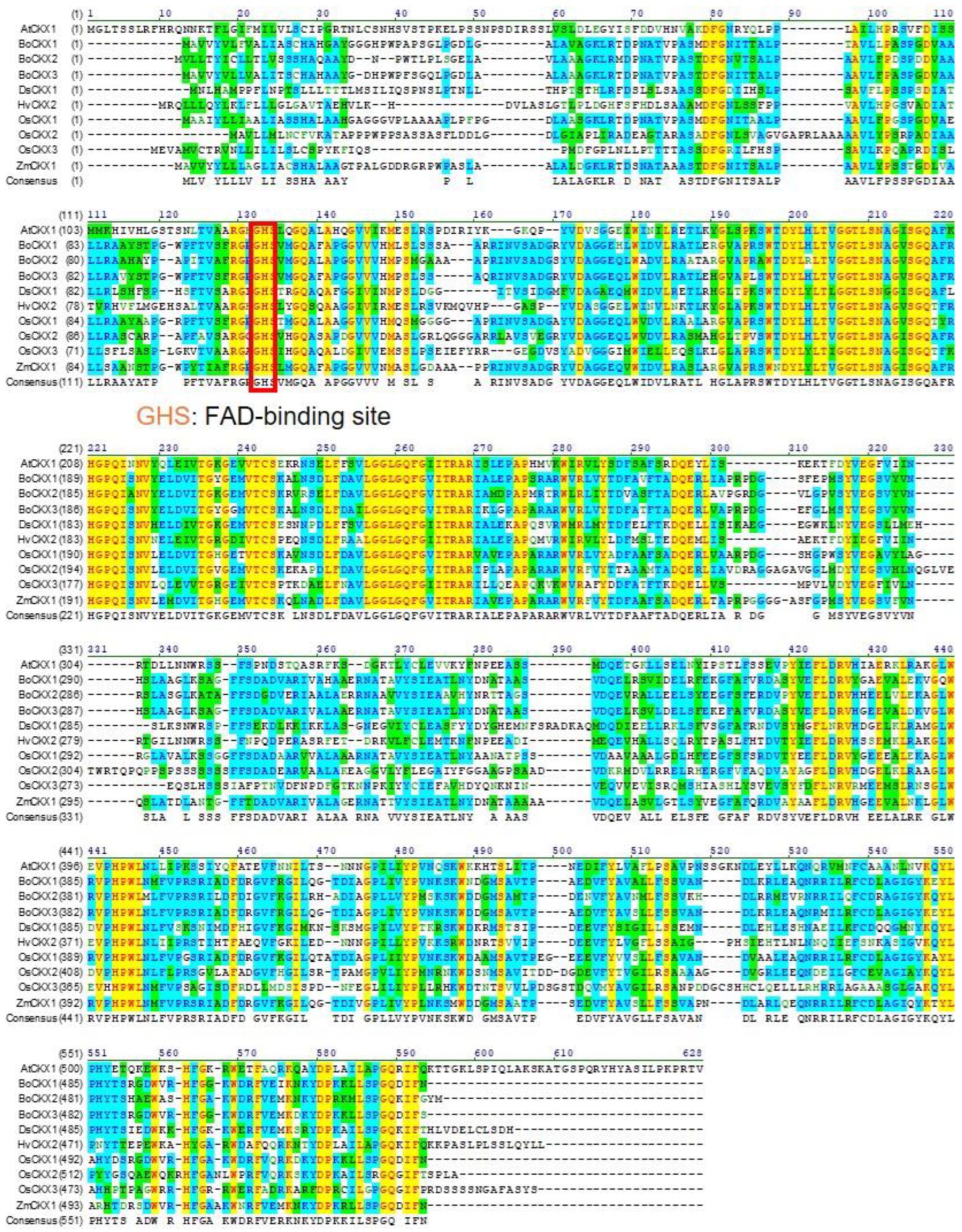
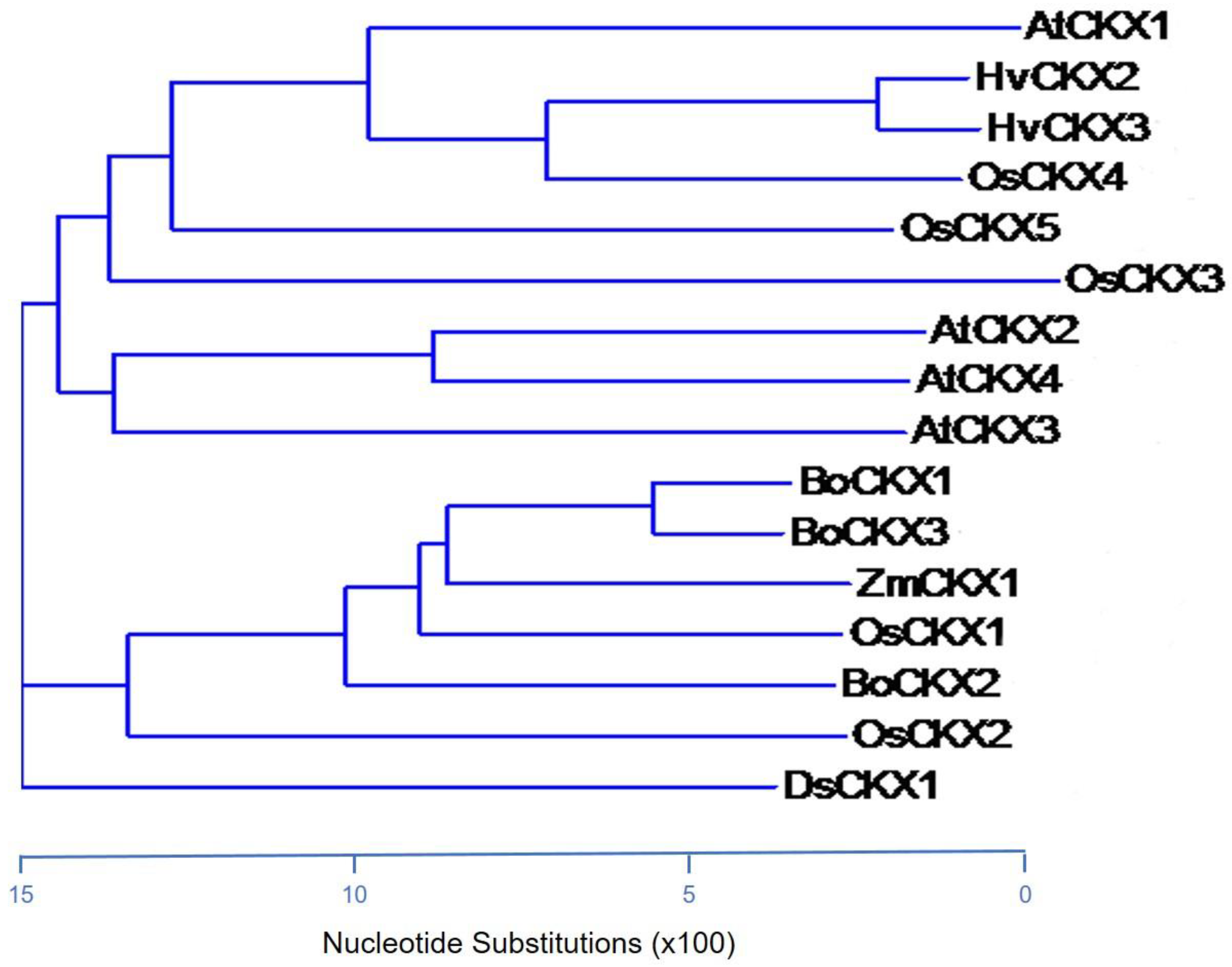
3.3. Protein Similarity of CKXs Proteins
3.4. Analysis of the cis-Acting Elements in BoCKX1
3.5. Analysis of the cis-Acting Elements in BoCKX2
3.6. Analysis of the cis-Acting Elements in BoCKX3
3.7. Comparison of Features in Plant CKX Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gan, S.; Amasino, R.M. Inhibition of leaf senescence by autoregulated production of cytokinin. Science 1995, 270, 1986–1988. [Google Scholar] [CrossRef]
- Mok, D.W.S.; Mok, M.C. Cytokinin metabolism and action. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 89–118. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Schmülling, T. Cytokinin action in plant development. Curr. Opin. Plant Biol. 2009, 12, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.C.; Weir, N.R.; Hill, K.; Zhang, W.; Kim, H.J.; Shiu, S.H.; Schaller, G.E.; Kieber, J.J. Characterization of genes involved in cytokinin signaling and metabolism from rice. Plant Physiol. 2012, 158, 1666–1684. [Google Scholar] [CrossRef] [Green Version]
- Jameson, P.E.; Song, J. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2016, 67, 593–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhao, J.; Song, J.; Jameson, P.E. Cytokinin glucosyl transferases, key regulators of cytokinin homeostasis, have potential value for wheat improvement. Plant Biotechnol. J. 2021, 19, 878–896. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.S.; Hsieh, L.S.; Yang, C.C.; Lee, P.D. Molecular characterization of isopentenyltransferase (BoAIPT1) from Bambusa oldhamii expressed in Escherichia coli. Bot. Stud. 2011, 52, 249–256. [Google Scholar]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashikari, M.; Sakakibara, H.; Lin, S.; Yamamoto, T.; Takashi, T.; Nishimura, A.; Angeles, E.R.; Qian, Q.; Kitano, H.; Matsuoka, M. Cytokinin oxidase regulates rice grain production. Science 2005, 309, 741–745. [Google Scholar] [CrossRef]
- Werner, T.; Köllmer, I.; Bartrina, I.; Holst, K.; Schmülling, T. New insights into the biology of cytokinin degradation. Plant Biol. 2006, 8, 371–381. [Google Scholar] [CrossRef]
- Pačes, V.; Werstiuk, E.; Hall, R.H. Conversion of N6-(D2-isopentenyl) adenosine to adenosine by enzyme activity in tobacco tissue. Plant Physiol. 1971, 48, 775–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitty, C.D.; Hall, R.H. A cytokinin oxidase in Zea mays. Can. J. Biochem. 1974, 52, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Houba-Hérin, N.; Pethe, C.; d’Alayer, J.; Laloue, M. Cytokinin oxidase from Zea mays: Purification, cDNA cloning and expression in moss protoplasts. Plant J. 1999, 17, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.O.; Bilyeu, K.D.; Laskey, J.G.; Cheikh, N.N. Isolation of a gene encoding a glycosylated cytokinin oxidase from maize. Biochem. Biophys. Res. Commun. 1999, 255, 328–333. [Google Scholar] [CrossRef]
- Vision, T.J.; Brown, D.G.; Tanksley, S.D. The origins of genomic duplications in Arabidopsis. Science 2000, 290, 2114–2117. [Google Scholar] [CrossRef] [Green Version]
- Niemann, M.C.E.; Weber, H.; Hluska, T.; Leonte, G.; Anderson, S.M.; Novák, O.; Senes, A.; Werner, T. The cytokinin oxidase/dehydrogenase CKX1 is a membrane-bound protein requiring homooligomerization in the endoplasmic reticulum for its cellular activity. Plant Physiol. 2018, 176, 2024–2039. [Google Scholar] [CrossRef]
- Galuszka, P.; Frébortová, J.; Werner, T.; Yamada, M.; Strnad, M.; Schmülling, T.; Frébort, I. Cytokinin oxidase/dehydrogenase genes in barley and wheat: Cloning and heterologous expression. Eur. J. Biochem. 2004, 271, 3990–4002. [Google Scholar] [CrossRef]
- Gasparis, S.; Przyborowski, M.; Kała, M.; Nadolska-Orczyk, A. Knockout of the HvCKX1 or HvCKX3 gene in barley (Hordeum vulgare L.) by RNA-guided cas9 nuclease affects the regulation of cytokinin metabolism and root morphology. Cells 2019, 8, 782. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Zhang, L.; Fu, Q.; Xu, Z.F. Identification and expression analysis of cytokinin metabolic genes IPTs, CYP735A and CKXs in the biofuel plant Jatropha curcas. PeerJ 2018, 6, e4812. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Fang, J.; Xu, F.; Wang, W.; Sun, X.; Chu, J.; Cai, B.; Feng, Y.; Chu, C. Cytokinin oxidase/dehydrogenase 4 integrates cytokinin and auxin signaling to control rice crown root formation. Plant Physiol. 2014, 165, 1035–1046. [Google Scholar] [CrossRef] [Green Version]
- Yeh, S.Y.; Chen, H.W.; Ng, C.Y.; Lin, C.Y.; Tseng, T.H.; Li, W.H.; Ku, M. Down-regulation of cytokinin oxidase 2 expression increases tiller number and improves rice yield. Rice 2015, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massonneau, A.; Houba-Hérin, N.; Pethe, C.; Madzak, C.; Falque, M.; Mercy, M.; Kopecny, D.; Majira, A.; Rogowsky, P.; Laloue, M. Maize cytokinin oxidase genes: Differential expression and cloning of two new cDNAs. J. Exp. Bot. 2004, 55, 2549–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smehilová, M.; Galuszka, P.; Bilyeu, K.D.; Jaworek, P.; Kowalska, M.; Sebela, M.; Sedlárová, M.; English, J.T.; Frébort, I. Subcellular localization and biochemical comparison of cytosolic and secreted cytokinin dehydrogenase enzymes from maize. J. Exp. Bot. 2009, 60, 2701–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Zhang, C.; Ma, J.Q.; Zhang, L.Y.; Yang, B.; Tang, X.Y.; Huang, L.; Zhou, X.T.; Lu, K.; Li, J.N. Genome-wide identification and expression profiling of cytokinin oxidase/dehydrogenase (CKX) genes reveal likely roles in pod development and stress responses in oilseed rape (Brassica napus L.). Genes 2018, 9, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugière, N.; Jiao, S.; Hantke, S.; Zinselmeier, C.; Roessler, J.A.; Niu, X.; Jones, R.J.; Habben, J.E. Cytokinin oxidase gene expression in maize is localized to the vasculature, and is induced by cytokinins, abscisic acid, and abiotic stress. Plant Physiol. 2003, 132, 1228–1240. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Jiang, L.; Jameson, P.E. Co-ordinate regulation of cytokinin gene family members during flag leaf and reproductive development in wheat. BMC Plant Biol. 2012, 12, 78. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Jiang, L.; Jameson, P.E. Expression patterns of Brassica napus genes implicate IPT, CKX, sucrose transporter, cell wall invertase and amino acid permease gene family members in leaf, flower, silique and seed development. J. Exp. Bot. 2015, 66, 5067–5082. [Google Scholar] [CrossRef] [Green Version]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H.; Schmülling, T. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2003, 15, 2532–2550. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, R.; Watanabe, Y.; Fujita, Y.; Le, D.T.; Kojima, M.; Werner, T.; Vankova, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Kakimoto, T.; et al. Analysis of cytokinin mutants and regulation of cytokinin metabolic genes reveals important regulatory roles of cytokinins in drought, salt and abscisic acid responses, and abscisic acid biosynthesis. Plant Cell 2011, 23, 2169–2183. [Google Scholar] [CrossRef] [Green Version]
- Macková, H.; Hronková, M.; Dobrá, J.; Turečková, V.; Novák, O.; Lubovská, Z.; Motyka, V.; Haisel, D.; Hájek, T.; Prášil, I.T.; et al. Enhanced drought and heat stress tolerance of tobacco plants with ectopically enhanced cytokinin oxidase/dehydrogenase gene expression. J. Exp. Bot. 2013, 64, 2805–2815. [Google Scholar] [CrossRef]
- Lubovská, Z.; Dobrá, J.; Storchová, H.; Wilhelmová, N.; Vanková, R. Cytokinin oxidase/dehydrogenase overexpression modifies antioxidant defense against heat, drought and their combination in Nicotiana tabacum plants. J. Plant Physiol. 2014, 171, 1625–1633. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.; Sahoo, K.K.; Tripathi, A.K.; Kumar, R.; Gupta, B.K.; Pareek, A.; Singla-Pareek, S.L. Knockdown of an inflorescence meristem-specific cytokinin oxidase—OsCKX2 in rice reduces yield penalty under salinity stress condition. Plant Cell Environ. 2018, 41, 936–946. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Comandini, O. Cytokinin oxidase strikes again. Trends Plant Sci. 1999, 4, 300. [Google Scholar] [CrossRef] [PubMed]
- Bilyeu, K.D.; Cole, J.L.; Laskey, J.G.; Riekhof, W.R.; Esparza, T.J.; Kramer, M.D.; Morris, R.O. Molecular and biochemical characterization of a cytokinin oxidase from maize. Plant Physiol. 2001, 125, 378–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmülling, T.; Werner, T.; Riefler, M.; Krupková, E.; Bartrina y Manns, I. Structure and function of cytokinin oxidase/dehydrogenase genes of maize, rice, Arabidopsis and other species. J. Plant Res. 2003, 116, 241–252. [Google Scholar] [CrossRef]
- Bilyeu, K.D.; Frébort, I. Vacuolar and cytosolic cytokinin dehydrogenases of Arabidopsis thaliana: Heterologous expression, purification and properties. Phytochemistry 2010, 71, 1970–1978. [Google Scholar] [CrossRef]
- Köllmer, I.; Novák, O.; Strnad, M.; Schmülling, T.; Werner, T. Overexpression of the cytosolic cytokinin oxidase/dehydrogenase (CKX7) from Arabidopsis causes specific changes in root growth and xylem differentiation. Plant J. 2014, 78, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, L.S.; Ma, G.J.; Yang, C.C.; Lee, P.D. Cloning, expression, site-directed mutagenesis and immunolocalization of phenylalanine ammonia-lyase in Bambusa oldhamii. Phytochemistry 2010, 71, 1999–2009. [Google Scholar] [CrossRef]
- Hsieh, L.S.; Hsieh, Y.L.; Yeh, C.S.; Cheng, C.Y.; Yang, C.C.; Lee, P.D. Molecular characterization of a phenylalanine ammonia-lyase gene (BoPAL1) from Bambusa oldhamii. Mol. Biol. Rep. 2011, 38, 283–290. [Google Scholar] [CrossRef]
- Hsieh, C.Y.; Huang, Y.H.; Lin, Z.Y.; Hsieh, L.S. Insights into the substrate selectivity of Bambusa oldhamii phenylalanine ammonia-lyase 1 and 2 through mutational analysis. Phytochem. Lett. 2020, 38, 140–143. [Google Scholar] [CrossRef]
- Hsiao, C.J.; Hsieh, C.Y.; Hsieh, L.S. Cloning and characterization of the Bambusa oldhamii BoMDH-encoded malate dehydrogenase. Protein Expr. Purif. 2020, 174, 105665. [Google Scholar] [CrossRef]
- Hong, P.Y.; Huang, Y.H.; Lim, G.G.C.W.; Chen, Y.P.; Hsiao, C.J.; Chen, L.H.; Ciou, J.Y.; Hsieh, L.S. Production of trans-cinnamic acid by immobilization of the Bambusa oldhamii BoPAL1 and BoPAL2 phenylalanine ammonia-lyases on electrospun nanofibers. Int. J. Mol. Sci. 2021, 22, 11184. [Google Scholar] [CrossRef]
- Hsieh, C.Y.; Huang, Y.H.; Yeh, H.H.; Hong, P.Y.; Hsiao, C.J.; Hsieh, L.S. Phenylalanine, tyrosine, and DOPA are bona fide substrates for Bambusa oldhamii BoPAL4. Catalysts 2021, 11, 1263. [Google Scholar] [CrossRef]
- Huang, Y.H.; You, W.C.; Hong, P.Y.; Chang, Y.T.; Ciou, J.Y.; Hsieh, L.S. Molecular characterization of the Bambusa oldhamii BoPAL3-encoded phenylalanine ammonia-lyase. Phytochem. Lett. 2022, 48, 15–18. [Google Scholar] [CrossRef]
- Gamuyao, R.; Nagai, K.; Ayano, M.; Mori, Y.; Minami, A.; Kojima, M.; Suzuki, T.; Sakakibara, H.; Higashiyama, T.; Ashikari, M.; et al. Hormone distribution and transcriptome profiles in bamboo shoots provide insights on bamboo stem emergence and growth. Plant Cell Physiol. 2017, 58, 702–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakočiūnė, D.; Moodley, A. A rapid bacteriophage DNA extraction method. Methods Protoc. 2018, 1, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; Jung, E.; Brunak, S. Prediction of N-glycosylation Sites in Human Proteins. 2004. Available online: https://www.cbs.dtu.dk/services/NetNGlyc/ (accessed on 15 March 2020).
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; Lee, T.Y.; Huang, H.D.; Huang, H.Y.; Pan, R.L. PlantPAN: Plant promoter analysis navigator, for identifying combinatorial cis-regulatory elements with distance constraint in plant gene group. BMC Genom. 2008, 9, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahmuradov, I.A.; Gammerman, A.J.; Hancock, J.M.; Bramley, P.M.; Solovyev, V.V. PlantProm: A database of plant promoter sequences. Nucleic Acids Res. 2003, 31, 114–117. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.H.; Yu, H.; Goh, C.J. Functional characterisation of a cytokinin oxidase gene DsCKX1 in Dendrobium orchid. Plant Mol. Biol. 2003, 51, 237–248. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Xu, J.; Wang, Y.; Yang, K. Transcription factor ANACo74 binds to NRS1, NRS2, or MybSt1 element in addition to the NACRS to regulate gene expression. Int. J. Mol. Sci. 2018, 19, 3271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, A.; Pati, P.K.; Pati, A.M.; Nagpal, A.K. In-silico analysis of cis-acting regulatory elements of pathogenesis-related proteins of Arabidopsis thaliana and Oryza sativa. PLoS ONE 2017, 12, e0184523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, M.R.; Lindskog, I.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Hochstrasser, D.F.; Appel, R.D. Detailed peptide characterization using PEPTIDEMASS: A World-Wide Web accessible tool. Electrophoresis 1997, 18, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Nakai, K.; Horton, P. PSORT: A program for detecting sorting signals in proteins and predicting their subcellular localization. Trends Biochem. Sci. 1999, 24, 34–36. [Google Scholar] [CrossRef]
- Lam, E.; Cha, N.H. ASF-2: A factor that binds to the cauliflower mosaic virus 35S promoter and a conserved GATA motif in cab promoter. Plant Cell 1989, 1, 1147–1156. [Google Scholar] [CrossRef]
- Benfey, P.N.; Cha, N.H. The cauliflower mosaic virus 35S promoter: Combinatorial regulation of transcription in plants. Science 1990, 250, 959–966. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Aoyama, T.; Oka, A. Arabidopsis ARR1 and ARR2 response regulators operate as transcriptional activators. Plant J. 2000, 24, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.; Honma, T.; Sato, S.; Kato, T.; Tabata, S.; Oka, A. ARR1, a transcription factor for genes immediately responsive to cytokinins. Science 2001, 294, 1519–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BoCKX1 | BoCKX2 | BoCKX3 | |
|---|---|---|---|
| BoCKX1 | 100 | 78 | 92 |
| BoCKX2 | 100 | 79 | |
| BoCKX3 | 100 |
| Gene Name a | No. of Exons | Length (aa) | Mass (kDa) b | Subcellular Localization (PSORT) c | Glycosylation Sites d | Phosphorylation Sites e Ser Thr Tyr | Reference | ||
|---|---|---|---|---|---|---|---|---|---|
| BoCKX1 | 3 | 525 | 57.0 | S f | 7/6 | 11/8 | 4/1 | 6/3 | This study |
| BoCKX2 | 4 | 523 | 57.4 | S | 5/5 | 14/8 | 5/3 | 8/3 | This study |
| BoCKX3 | 3 | 522 | 56.6 | S | 6/5 | 11/8 | 3/1 | 8/4 | This study |
| DsCKX1 | 4 | 536 | 60.4 | S | 2/2 | h - | - | - | [51] |
| OsCKX1 | 3 | 558 | 59.1 | S | 2/1 | - | - | - | [35] |
| OsCKX2 | 4 | 532 | 56.0 | M g | 6/5 | - | - | - | [35] |
| OsCKX3 | 5 | 525 | 58.0 | S | 3/3 | - | - | - | [35] |
| ZmCKX1 | 3 | 534 | 57.2 | S | 8/5 | - | - | - | [22] |
| Cis-Element | Consensus Sequence | Function | References |
|---|---|---|---|
| TATA box | TATAAT | Core promoter cis-element of genes in eukaryotes | [50] |
| CAAT box | CAAT | The CAAT box is a conserved consensus sequence as the binding site of the RNA transcriptional factor | [50] |
| As-1 box | TGACG | Activation sequence 1 (as-1) is a salicylic acid (SA)- and auxin-responsive element | [53,56,57] |
| NGATT | N=G/A/C/T GATT | Arabidopsis cytokinin response regulators ARR1 binding element | [52,58,59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, C.-Y.; Hsieh, L.-S. Cloning of Three Cytokinin Oxidase/Dehydrogenase Genes in Bambusa oldhamii. Curr. Issues Mol. Biol. 2023, 45, 1902-1913. https://doi.org/10.3390/cimb45030123
Hsieh C-Y, Hsieh L-S. Cloning of Three Cytokinin Oxidase/Dehydrogenase Genes in Bambusa oldhamii. Current Issues in Molecular Biology. 2023; 45(3):1902-1913. https://doi.org/10.3390/cimb45030123
Chicago/Turabian StyleHsieh, Chun-Yen, and Lu-Sheng Hsieh. 2023. "Cloning of Three Cytokinin Oxidase/Dehydrogenase Genes in Bambusa oldhamii" Current Issues in Molecular Biology 45, no. 3: 1902-1913. https://doi.org/10.3390/cimb45030123
APA StyleHsieh, C. -Y., & Hsieh, L. -S. (2023). Cloning of Three Cytokinin Oxidase/Dehydrogenase Genes in Bambusa oldhamii. Current Issues in Molecular Biology, 45(3), 1902-1913. https://doi.org/10.3390/cimb45030123