

Natural Products for the Treatment of Pulmonary Hypertension: Mechanism, Progress, and Future Opportunities


Abstract
:1. Introduction
2. Pathological Mechanisms of PH
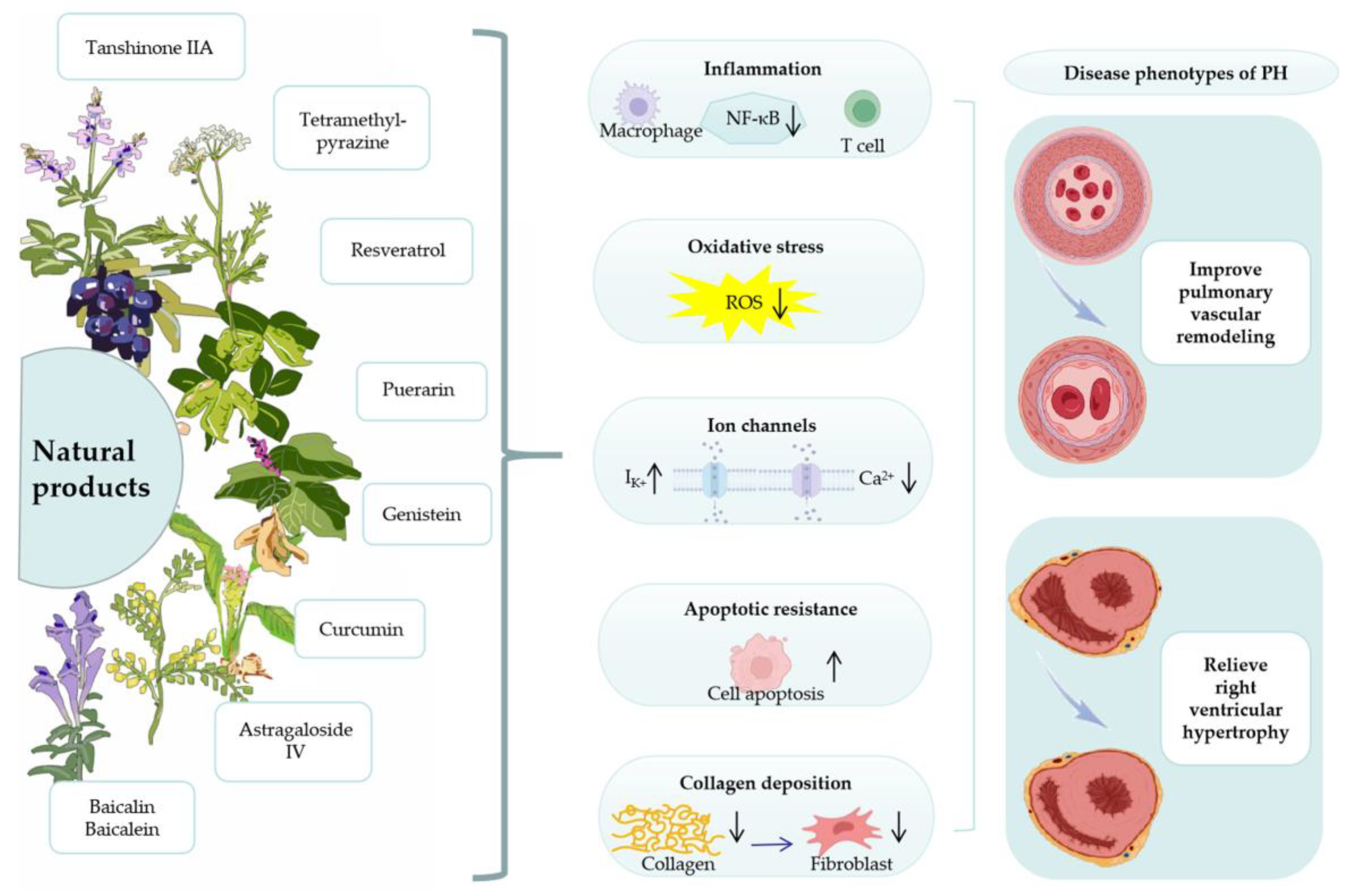
3. Natural Products for the Treatment of PH
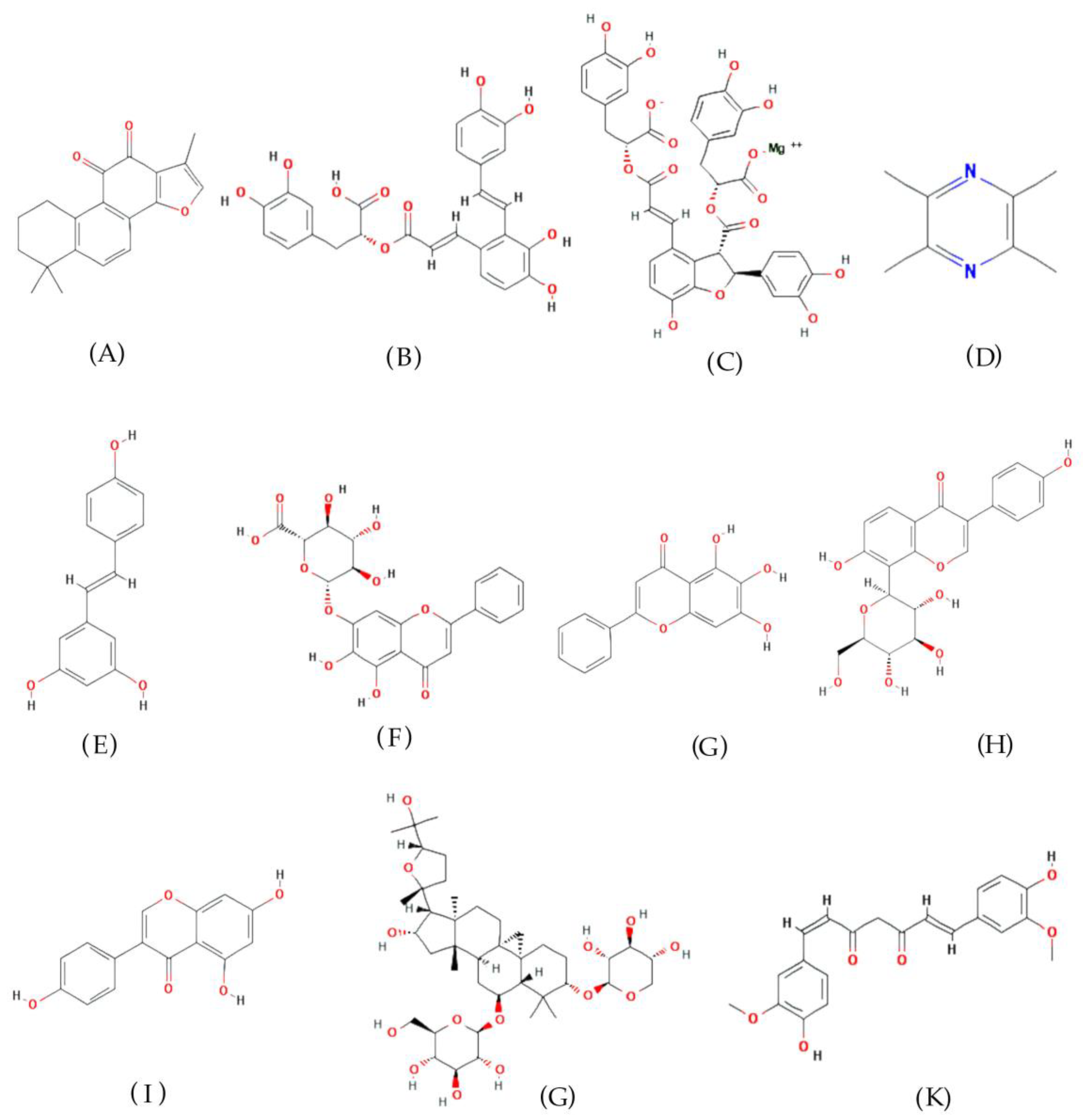
3.1. Tanshinone IIA
3.2. Other Active Natural Products Derived from Danshen
3.3. Tetramethylpyrazine
3.4. Resveratrol
3.5. Baicalin and Baicalein
3.6. Puerarin
3.7. Genistein
3.8. Astragaloside IV
3.9. Curcumin
4. Mechanism of Natural Products for the Treatment of PH
4.1. Anti-Inflammatory
4.2. Oxidative Stress
4.3. Ion Channels
4.4. Apoptotic Resistance
4.5. Collagen Deposition
4.6. Right Ventricle Protection
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poch, D.; Mandel, J. Pulmonary Hypertension. Ann. Intern. Med. 2021, 174, Itc49–Itc64. [Google Scholar] [CrossRef]
- Thenappan, T.; Ormiston, M.L.; Ryan, J.J.; Archer, S.L. Pulmonary arterial hypertension: Pathogenesis and clinical management. BMJ 2018, 360, j5492. [Google Scholar] [CrossRef]
- Levine, D.J. Pulmonary arterial hypertension: Updates in epidemiology and evaluation of patients. Am. J. Manag. Care 2021, 27, S35–S41. [Google Scholar] [CrossRef]
- Zolty, R. Pulmonary arterial hypertension specific therapy: The old and the new. Pharmacol. Ther. 2020, 214, 107576. [Google Scholar] [CrossRef]
- Sommer, N.; Ghofrani, H.A.; Pak, O.; Bonnet, S.; Provencher, S.; Sitbon, O.; Rosenkranz, S.; Hoeper, M.M.; Kiely, D.G. Current and future treatments of pulmonary arterial hypertension. Br. J. Pharmacol. 2021, 178, 6–30. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Chen, J.; Gao, Y.; Deng, B.; Liu, K. Endothelin receptor antagonists for pulmonary arterial hypertension. Cochrane Database Syst. Rev. 2021, 3, Cd004434. [Google Scholar] [CrossRef]
- Cristo Ropero, M.J.; Cruz-Utrilla, A.; Escribano-Subias, M.P. Epoprostenol for the treatment of pulmonary arterial hypertension. Expert Rev. Clin. Pharmacol. 2021, 14, 1005–1013. [Google Scholar] [CrossRef]
- Bhogal, S.; Khraisha, O.; Al Madani, M.; Treece, J.; Baumrucker, S.J.; Paul, T.K. Sildenafil for Pulmonary Arterial Hypertension. Am. J. Ther. 2019, 26, e520–e526. [Google Scholar] [CrossRef] [Green Version]
- de Araujo, A.; Mantovani, A.; Cerski, C.T.S.; Lopes, A.B.; Bortoncello, L.C.; Gazzana, M.B.; Cheinquer, H. Bosentan Inducing Autoimmune Hepatitis in a Patient with Idiopathic Pulmonary Arterial Hypertension. J. Gastrointestin. Liver Dis. 2018, 27, 89–92. [Google Scholar] [CrossRef] [Green Version]
- Abman, S.H.; Kinsella, J.P.; Rosenzweig, E.B.; Krishnan, U.; Kulik, T.; Mullen, M.; Wessel, D.L.; Steinhorn, R.; Adatia, I.; Hanna, B.; et al. Implications of the U.S. Food and Drug Administration warning against the use of sildenafil for the treatment of pediatric pulmonary hypertension. Am. J. Respir Crit. Care Med. 2013, 187, 572–575. [Google Scholar] [CrossRef]
- Xiang, L.; Li, Y.; Deng, X.; Kosanovic, D.; Schermuly, R.T.; Li, X. Natural plant products in treatment of pulmonary arterial hypertension. Pulm. Circ. 2018, 8, 2045894018784033. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.R.; Ouyang, X.; Hou, C.; Yang, Q.F.; Wu, Y.; Lu, W.J.; Liu, C.L.; Yang, K.; Zhong, N.S.; Wang, J.; et al. Natural ingredients from Chinese materia medica for pulmonary hypertension. Chin. J. Nat. Med. 2021, 19, 801–814. [Google Scholar] [CrossRef]
- Wang, T.; Hou, J.; Xiao, W.; Zhang, Y.; Zhou, L.; Yuan, L.; Yin, X.; Chen, X.; Hu, Y. Chinese medicinal plants for the potential management of high-altitude pulmonary oedema and pulmonary hypertension. Pharm. Biol. 2020, 58, 815–827. [Google Scholar] [CrossRef]
- Chang, Z.; Zhang, P.; Zhang, M.; Jun, F.; Hu, Z.; Yang, J.; Wu, Y.; Zhou, R. Aloperine suppresses human pulmonary vascular smooth muscle cell proliferation via inhibiting inflammatory response. Chin. J. Physiol. 2019, 62, 157–165. [Google Scholar] [CrossRef]
- Xu, D.; Hu, Y.H.; Gou, X.; Li, F.Y.; Yang, X.Y.; Li, Y.M.; Chen, F. Oxidative Stress and Antioxidative Therapy in Pulmonary Arterial Hypertension. Molecules 2022, 27, 3724. [Google Scholar] [CrossRef]
- Bourgeois, A.; Omura, J.; Habbout, K.; Bonnet, S.; Boucherat, O. Pulmonary arterial hypertension: New pathophysiological insights and emerging therapeutic targets. Int. J. Biochem. Cell. Biol. 2018, 104, 9–13. [Google Scholar] [CrossRef]
- Santos-Gomes, J.; Le Ribeuz, H.; Brás-Silva, C.; Antigny, F.; Adão, R. Role of Ion Channel Remodeling in Endothelial Dysfunction Induced by Pulmonary Arterial Hypertension. Biomolecules 2022, 12, 484. [Google Scholar] [CrossRef]
- Thenappan, T.; Chan, S.Y.; Weir, E.K. Role of extracellular matrix in the pathogenesis of pulmonary arterial hypertension. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1322–H1331. [Google Scholar] [CrossRef]
- Evans, C.E.; Cober, N.D.; Dai, Z.; Stewart, D.J.; Zhao, Y.Y. Endothelial cells in the pathogenesis of pulmonary arterial hypertension. Eur. Respir J. 2021, 58, 2003957. [Google Scholar] [CrossRef]
- Kylhammar, D.; Rådegran, G. The principal pathways involved in the in vivo modulation of hypoxic pulmonary vasoconstriction, pulmonary arterial remodelling and pulmonary hypertension. Acta Physiol. 2017, 219, 728–756. [Google Scholar] [CrossRef]
- Dunham-Snary, K.J.; Wu, D.; Sykes, E.A.; Thakrar, A.; Parlow, L.R.G.; Mewburn, J.D.; Parlow, J.L.; Archer, S.L. Hypoxic Pulmonary Vasoconstriction: From Molecular Mechanisms to Medicine. Chest 2017, 151, 181–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuder, R.M. Pulmonary vascular remodeling in pulmonary hypertension. Cell Tissue Res. 2017, 367, 643–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guignabert, C.; Tu, L.; Girerd, B.; Ricard, N.; Huertas, A.; Montani, D.; Humbert, M. New molecular targets of pulmonary vascular remodeling in pulmonary arterial hypertension: Importance of endothelial communication. Chest 2015, 147, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Gorelova, A.; Berman, M.; Al Ghouleh, I. Endothelial-to-Mesenchymal Transition in Pulmonary Arterial Hypertension. Antioxid Redox Signal 2021, 34, 891–914. [Google Scholar] [CrossRef] [PubMed]
- Rawal, H.; Suman, A.; Bhoite, R.R.; Kanwal, A.; Young, R.K.; Aronow, W.S.; Lavie, C.; Ghosh, R.K. Anticoagulation in Pulmonary Arterial Hypertension: Do We Know the Answer? Curr. Probl. Cardiol. 2021, 46, 100738. [Google Scholar] [CrossRef]
- Shen, T.; Shi, J.; Wang, N.; Yu, X.; Zhang, C.; Li, J.; Wei, L.; Ma, C.; Zhao, X.; Lian, M.; et al. 15-Lipoxygenase and 15-hydroxyeicosatetraenoic acid regulate intravascular thrombosis in pulmonary hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L449–L462. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Wang, Z.; Jiang, X.; Zhao, Q.; Shen, J. Targeted Drugs for Treatment of Pulmonary Arterial Hypertension: Past, Present, and Future Perspectives. J. Med. Chem. 2020, 63, 15153–15186. [Google Scholar] [CrossRef]
- Yu, Z.; Xiao, J.; Chen, X.; Ruan, Y.; Chen, Y.; Zheng, X.; Wang, Q. Bioactivities and mechanisms of natural medicines in the management of pulmonary arterial hypertension. Chin. Med. 2022, 17, 13. [Google Scholar] [CrossRef]
- Li, X.; Xiang, D.; Shu, Y.; Zeng, X.; Li, Y. Mitigating effect of tanshinone IIA on ventricular remodeling in rats with pressure overload-induced heart failure. Acta Cir. Bras. 2019, 34, e201900807. [Google Scholar] [CrossRef]
- Guo, R.; Li, L.; Su, J.; Li, S.; Duncan, S.E.; Liu, Z.; Fan, G. Pharmacological Activity and Mechanism of Tanshinone IIA in Related Diseases. Drug Des. Devel. Ther. 2020, 14, 4735–4748. [Google Scholar] [CrossRef]
- Wang, J.; Weigand, L.; Wang, W.; Sylvester, J.T.; Shimoda, L.A. Chronic hypoxia inhibits Kv channel gene expression in rat distal pulmonary artery. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, L1049–L1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Weigand, L.; Lu, W.; Sylvester, J.T.; Semenza, G.L.; Shimoda, L.A. Hypoxia inducible factor 1 mediates hypoxia-induced TRPC expression and elevated intracellular Ca2+ in pulmonary arterial smooth muscle cells. Circ. Res. 2006, 98, 1528–1537. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Liu, M.; Wei, M.; Liu, Y.; Dong, M.; Luo, Y.; Zhao, P.; Dong, H.; Niu, W.; Yan, Z.; et al. Tanshinone IIA attenuates hypoxic pulmonary hypertension via modulating KV currents. Respir. Physiol. Neurobiol. 2015, 205, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, Q.; Wan, L.; Yang, K.; Zhang, Y.; Chen, Y.; Wang, E.; Lai, N.; Zhao, L.; Jiang, H.; et al. Sodium tanshinone IIA sulfonate inhibits canonical transient receptor potential expression in pulmonary arterial smooth muscle from pulmonary hypertensive rats. Am. J. Respir Cell. Mol. Biol. 2013, 48, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Q.; Lu, W.; Yang, K.; Hadadi, C.; Fu, X.; Chen, Y.; Yun, X.; Zhang, J.; Li, M.; Xu, L.; et al. Sodium tanshinone IIA sulfonate inhibits hypoxia-induced enhancement of SOCE in pulmonary arterial smooth muscle cells via the PKG-PPAR-γ signaling axis. Am. J. Physiol. Cell Physiol. 2016, 311, C136–C149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Liu, W.; Lu, W.; Luo, X.; Lin, Y.; Liu, S.; Qian, J.; Zhang, C.; Chen, H.; Li, Y.; et al. Sodium tanshinone IIA sulfonate enhances the BMP9-BMPR2-Smad1/5/9 signaling pathway in rat pulmonary microvascular endothelial cells and human embryonic stem cell-derived endothelial cells. Biochem. Pharmacol. 2022, 199, 114986. [Google Scholar] [CrossRef]
- Xu, Y.; Sun, Z.; Wang, Q.; Wang, T.; Liu, Y.; Yu, F. Stox1 induced the proliferation and cell cycle arrest in pulmonary artery smooth muscle cells via AKT signaling pathway. Vascul. Pharmacol. 2019, 120, 106568. [Google Scholar] [CrossRef]
- Bao, Y.R.; Chen, J.W.; Jiang, Y.; Wang, L.H.; Xue, R.; Qian, J.X.; Zhang, G.X. Sodium Tanshinone II Sulfonate A Ameliorates Hypoxia-Induced Pulmonary Hypertension. Front. Pharmacol. 2020, 11, 687. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Wang, W.; Zhang, N.; Wu, H.; Liu, C.; Chen, X.; Chen, Y.; Chen, Y.; Jiang, Q.; et al. Promising therapeutic effects of sodium tanshinone IIA sulfonate towards pulmonary arterial hypertension in patients. J. Thorac. Dis. 2013, 5, 169–172. [Google Scholar] [CrossRef]
- Suzuki, T.; Carrier, E.J.; Talati, M.H.; Rathinasabapathy, A.; Chen, X.; Nishimura, R.; Tada, Y.; Tatsumi, K.; West, J. Isolation and characterization of endothelial-to-mesenchymal transition cells in pulmonary arterial hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, L118–L126. [Google Scholar] [CrossRef]
- Yuan, T.; Chen, Y.; Zhang, H.; Fang, L.; Du, G. Salvianolic Acid A, a Component of Salvia miltiorrhiza, Attenuates Endothelial-Mesenchymal Transition of HPAECs Induced by Hypoxia. Am. J. Chin. Med. 2017, 45, 1185–1200. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yuan, T.; Zhang, H.; Yan, Y.; Wang, D.; Fang, L.; Lu, Y.; Du, G. Activation of Nrf2 Attenuates Pulmonary Vascular Remodeling via Inhibiting Endothelial-to-Mesenchymal Transition: An Insight from a Plant Polyphenol. Int. J. Biol. Sci. 2017, 13, 1067–1081. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Luo, X.J.; Wang, E.L.; Li, N.S.; Zhang, X.J.; Song, F.L.; Yang, J.F.; Liu, B.; Peng, J. Magnesium lithospermate B prevents phenotypic transformation of pulmonary arteries in rats with hypoxic pulmonary hypertension through suppression of NADPH oxidase. Eur. J. Pharmacol. 2019, 847, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Peng, J.J.; Wang, E.L.; Li, N.S.; Song, F.L.; Yang, J.F.; Luo, X.J.; Liu, B.; Peng, J. Magnesium Lithospermate B Derived from Salvia miltiorrhiza Ameliorates Right Ventricle Remodeling in Pulmonary Hypertensive Rats via Inhibition of NOX/VPO1 Pathway. Planta Med. 2019, 85, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Duo, D.; Yan, Y.; He, R.; Wu, X. Magnesium lithospermate B ameliorates hypobaric hypoxia-induced pulmonary arterial hypertension by inhibiting endothelial-to-mesenchymal transition and its potential targets. Biomed. Pharmacother. 2020, 130, 110560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Dong, M.; Luo, Y.; Zhao, F.; Li, Y. Danshensu prevents hypoxic pulmonary hypertension in rats by inhibiting the proliferation of pulmonary artery smooth muscle cells via TGF-β-smad3-associated pathway. Eur. J. Pharmacol. 2018, 820, 1–7. [Google Scholar] [CrossRef]
- Wang, Y.; Duo, D.; Yan, Y.; He, R.; Wang, S.; Wang, A.; Wu, X. Extract of Salvia przewalskii Repair Tissue Damage in Chronic Hypoxia Maybe through the RhoA-ROCK Signalling Pathway. Biol. Pharm. Bull. 2020, 43, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Liu, Y.; Shi, D. Cardiovascular Actions and Therapeutic Potential of Tetramethylpyrazine (Active Component Isolated from Rhizoma Chuanxiong): Roles and Mechanisms. Biomed. Res. Int. 2016, 2016, 2430329. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Lu, W.; Yang, K.; Duan, X.; Li, M.; Chen, X.; Zhang, J.; Kuang, M.; Liu, S.; Wu, X.; et al. Tetramethylpyrazine: A promising drug for the treatment of pulmonary hypertension. Br. J. Pharmacol. 2020, 177, 2743–2764. [Google Scholar] [CrossRef]
- Huang, H.; Kong, L.; Luan, S.; Qi, C.; Wu, F. Ligustrazine Suppresses Platelet-Derived Growth Factor-BB-Induced Pulmonary Artery Smooth Muscle Cell Proliferation and Inflammation by Regulating the PI3K/AKT Signaling Pathway. Am. J. Chin. Med. 2021, 49, 437–459. [Google Scholar] [CrossRef]
- Zhang, H.; Chang, Z.; Mehmood, K.; Yang, M.K.; Liu, Z.; Duan, Z.; Yuan, F.; Ali, M.M.; Adnan, M.; Qasim, M.U.; et al. Tetramethylpyrazine inhibited hypoxia-induced expression of calcium-sensing receptors in pulmonary artery smooth muscle cells in chickens. J. Biol. Regul. Homeost Agents 2018, 32, 489–495. [Google Scholar]
- Zhang, L.; Deng, M.; Zhou, S. Tetramethylpyrazine inhibits hypoxia-induced pulmonary vascular leakage in rats via the ROS-HIF-VEGF pathway. Pharmacology 2011, 87, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.P.; Dong, W.P.; Yang, Y.C.; Zeng, Y.Y.; Liu, Y.; Dong, Z.; Ma, X.M.; Cao, Y.Q.; Bai, Y.Z.; Yang, B.; et al. Tetramethylpyrazine Improves Monocrotaline-Induced Pulmonary Hypertension through the ROS/iNOS/PKG-1 Axis. J. Healthc. Eng. 2022, 2022, 1890892. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, P.; Acosta, M.F.; Gomez, A.I.; Grijalva, C.; Tang, H.; Yuan, J.X.; Mansour, H.M. Design and Comprehensive Characterization of Tetramethylpyrazine (TMP) for Targeted Lung Delivery as Inhalation Aerosols in Pulmonary Hypertension (PH): In Vitro Human Lung Cell Culture and In Vivo Efficacy. Antioxidants 2021, 10, 427. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Duan, S.F. The acute effects of ligustrazini on hemodynamics and right cardiac function in pulmonary arterial hypertension secondary to chronic obstructive pulmonary disease. J. Tongji Med. Univ. 1989, 9, 100–102. [Google Scholar] [CrossRef]
- Breuss, J.M.; Atanasov, A.G.; Uhrin, P. Resveratrol and Its Effects on the Vascular System. Int. J. Mol. Sci. 2019, 20, 1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirhadi, E.; Roufogalis, B.D.; Banach, M.; Barati, M.; Sahebkar, A. Resveratrol: Mechanistic and therapeutic perspectives in pulmonary arterial hypertension. Pharmacol. Res. 2021, 163, 105287. [Google Scholar] [CrossRef]
- Chen, Y.; Yuan, T.; Chen, D.; Liu, S.; Guo, J.; Fang, L.; Du, G. Systematic analysis of molecular mechanism of resveratrol for treating pulmonary hypertension based on network pharmacology technology. Eur. J. Pharmacol. 2020, 888, 173466. [Google Scholar] [CrossRef]
- Csiszar, A.; Labinskyy, N.; Olson, S.; Pinto, J.T.; Gupte, S.; Wu, J.M.; Hu, F.; Ballabh, P.; Podlutsky, A.; Losonczy, G.; et al. Resveratrol prevents monocrotaline-induced pulmonary hypertension in rats. Hypertension 2009, 54, 668–675. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Zhai, C.; Feng, W.; Wang, J.; Zhu, Y.; Li, S.; Wang, Q.; Zhang, Q.; Yan, X.; Chai, L.; et al. Resveratrol inhibits monocrotaline-induced pulmonary arterial remodeling by suppression of SphK1-mediated NF-κB activation. Life Sci. 2018, 210, 140–149. [Google Scholar] [CrossRef]
- Chen, B.; Xue, J.; Meng, X.; Slutzky, J.L.; Calvert, A.E.; Chicoine, L.G. Resveratrol prevents hypoxia-induced arginase II expression and proliferation of human pulmonary artery smooth muscle cells via Akt-dependent signaling. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, L317–L325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal-Ramírez, J.; Silva-Platas, C.; Jerjes-Sánchez, C.; Ramos-González, M.R.; Vázquez-Garza, E.; Chapoy-Villanueva, H.; Ramírez-Rivera, A.; Zarain-Herzberg, Á.; García, N.; García-Rivas, G. Resveratrol Prevents Right Ventricle Dysfunction, Calcium Mishandling, and Energetic Failure via SIRT3 Stimulation in Pulmonary Arterial Hypertension. Oxid. Med. Cell Longev. 2021, 2021, 9912434. [Google Scholar] [CrossRef]
- Zhou, S.; Li, M.T.; Jia, Y.Y.; Liu, J.J.; Wang, Q.; Tian, Z.; Liu, Y.T.; Chen, H.Z.; Liu, D.P.; Zeng, X.F. Regulation of Cell Cycle Regulators by SIRT1 Contributes to Resveratrol-Mediated Prevention of Pulmonary Arterial Hypertension. Biomed. Res. Int. 2015, 2015, 762349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paffett, M.L.; Lucas, S.N.; Campen, M.J. Resveratrol reverses monocrotaline-induced pulmonary vascular and cardiac dysfunction: A potential role for atrogin-1 in smooth muscle. Vascul. Pharmacol. 2012, 56, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhao, B.; Qu, H.; Chen, S.; Hao, X.; Chen, S.; Qin, Z.; Chen, G.; Fan, Y. Sera and lungs metabonomics reveals key metabolites of resveratrol protecting against PAH in rats. Biomed. Pharmacother. 2021, 133, 110910. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Yuan, T.; Zeng, Z.; Liu, D.; Liu, C.; Guo, J.; Chen, Y. Mechanistic and therapeutic perspectives of baicalin and baicalein on pulmonary hypertension: A comprehensive review. Biomed. Pharmacother. 2022, 151, 113191. [Google Scholar] [CrossRef]
- Xue, X.; Zhang, S.; Jiang, W.; Wang, J.; Xin, Q.; Sun, C.; Li, K.; Qi, T.; Luan, Y. Protective effect of baicalin against pulmonary arterial hypertension vascular remodeling through regulation of TNF-α signaling pathway. Pharmacol. Res. Perspect. 2021, 9, e00703. [Google Scholar] [CrossRef]
- Yan, S.; Wang, Y.; Liu, P.; Chen, A.; Chen, M.; Yao, D.; Xu, X.; Wang, L.; Huang, X. Baicalin Attenuates Hypoxia-Induced Pulmonary Arterial Hypertension to Improve Hypoxic Cor Pulmonale by Reducing the Activity of the p38 MAPK Signaling Pathway and MMP-9. Evid. Based Complement Alternat. Med. 2016, 2016, 2546402. [Google Scholar] [CrossRef] [Green Version]
- Yan, G.; Wang, J.; Yi, T.; Cheng, J.; Guo, H.; He, Y.; Shui, X.; Wu, Z.; Huang, S.; Lei, W. Baicalin prevents pulmonary arterial remodeling in vivo via the AKT/ERK/NF-κB signaling pathways. Pulm. Circ. 2019, 9, 2045894019878599. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Wu, P.; Huang, F.; Xu, M.; Chen, M.; Huang, K.; Li, G.P.; Xu, M.; Yao, D.; Wang, L. Baicalin attenuates chronic hypoxia-induced pulmonary hypertension via adenosine A(2A) receptor-induced SDF-1/CXCR4/PI3K/AKT signaling. J. Biomed. Sci. 2017, 24, 52. [Google Scholar] [CrossRef] [Green Version]
- Hsu, W.L.; Lin, Y.C.; Jeng, J.R.; Chang, H.Y.; Chou, T.C. Baicalein Ameliorates Pulmonary Arterial Hypertension Caused by Monocrotaline through Downregulation of ET-1 and ET(A)R in Pneumonectomized Rats. Am. J. Chin. Med. 2018, 46, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Wei, Z.; Zhu, D.; Fu, N.; Wang, C.; Yin, S.; Liang, Y.; Xing, J.; Wang, X.; Wang, Y. Baicalein attenuates monocrotaline-induced pulmonary arterial hypertension by inhibiting vascular remodeling in rats. Pulm. Pharmacol. Ther. 2018, 48, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Zhu, D.; Wei, Z.; Fu, N.; Wang, C.; Liu, L.; Zhang, H.; Liang, Y.; Xing, J.; Wang, X.; et al. Baicalein attenuates monocrotaline-induced pulmonary arterial hypertension by inhibiting endothelial-to-mesenchymal transition. Life Sci. 2018, 207, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Teng, C.; Li, B.; Lin, C.; Xing, X.; Huang, F.; Yang, Y.; Li, Y.; Azevedo, H.S.; He, W. Targeted delivery of baicalein-p53 complex to smooth muscle cells reverses pulmonary hypertension. J. Control Release 2022, 341, 591–604. [Google Scholar] [CrossRef]
- Wei, S.Y.; Chen, Y.; Xu, X.Y. Progress on the pharmacological research of puerarin: A review. Chin. J. Nat. Med. 2014, 12, 407–414. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Q.; Zhang, C.; Sheng, J.; Li, S.; Li, W.; Yang, X.; Wang, X.; He, S.; Bai, J.; et al. Puerarin prevents progression of experimental hypoxia-induced pulmonary hypertension via inhibition of autophagy. J. Pharmacol. Sci. 2019, 141, 97–105. [Google Scholar] [CrossRef]
- Yuan, T.; Zhang, H.; Chen, D.; Chen, Y.; Lyu, Y.; Fang, L.; Du, G. Puerarin protects pulmonary arteries from hypoxic injury through the BMPRII and PPARγ signaling pathways in endothelial cells. Pharmacol. Rep. 2019, 71, 855–861. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, H.F.; Yuan, T.Y.; Sun, S.C.; Wang, R.R.; Wang, S.B.; Fang, L.H.; Lyu, Y.; Du, G.H. Puerarin-V prevents the progression of hypoxia- and monocrotaline-induced pulmonary hypertension in rodent models. Acta Pharmacol. Sin. 2022, 43, 2325–2339. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, Y.; Liu, P.; Di, P.; Li, M.; Wang, C. Puerarin exhibits antiinflammatory properties in gunpowder smog-induced acute lung injury in rats via regulation of the renin-angiotensin system and the NFκB signaling pathway. Exp. Ther. Med. 2021, 22, 809. [Google Scholar] [CrossRef]
- Wang, X.; Yan, J.; Xu, X.; Duan, C.; Xie, Z.; Su, Z.; Ma, H.; Ma, H.; Wei, X.; Du, X. Puerarin prevents LPS-induced acute lung injury via inhibiting inflammatory response. Microb. Pathog. 2018, 118, 170–176. [Google Scholar] [CrossRef]
- Cai, D.; Zhao, Y.; Yu, F. Puerarin ameliorates acute lung injury by modulating NLRP3 inflammasome-induced pyroptosis. Cell Death Discov. 2022, 8, 368. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Sanches Silva, A.; Sánchez-Machado, D.I.; López-Cervantes, J.; Daglia, M.; Nabavi, S.F.; Nabavi, S.M. Hypotensive effects of genistein: From chemistry to medicine. Chem. Biol. Interact. 2017, 268, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, K.; Humayoun Akhtar, M. An updated review of dietary isoflavones: Nutrition, processing, bioavailability and impacts on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1280–1293. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, D.; Liu, S.; Yuan, T.; Guo, J.; Fang, L.; Du, G. Systematic Elucidation of the Mechanism of Genistein against Pulmonary Hypertension via Network Pharmacology Approach. Int. J. Mol. Sci. 2019, 20, 5569. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Wu, Y.; Wang, M.; Wang, Y.; Tausif, R.; Yang, Y. Genistein rescues hypoxia-induced pulmonary arterial hypertension through estrogen receptor and β-adrenoceptor signaling. J. Nutr. Biochem. 2018, 58, 110–118. [Google Scholar] [CrossRef]
- Yang, Y.; Nie, W.; Yuan, J.; Zhang, B.; Wang, Z.; Wu, Z.; Guo, Y. Genistein activates endothelial nitric oxide synthase in broiler pulmonary arterial endothelial cells by an Akt-dependent mechanism. Exp. Mol. Med. 2010, 42, 768–776. [Google Scholar] [CrossRef] [Green Version]
- Qiu, L.; Yin, G.; Cheng, L.; Fan, Y.; Xiao, W.; Yu, G.; Xing, M.; Jia, R.; Sun, R.; Ma, X.; et al. Astragaloside IV ameliorates acute pancreatitis in rats by inhibiting the activation of nuclear factor-κB. Int. J. Mol. Med. 2015, 35, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Feng, Y.; Gao, X.; Hao, X.; Zhang, J.; Long, Y.; Zhang, S.; Li, L. Astragaloside IV in Hypoxic Pulmonary Hypertension: An In Vivo and In Vitro Experiments. Appl. Biochem. Biotechnol. 2022, 194, 6319–6334. [Google Scholar] [CrossRef]
- Jin, H.; Jiao, Y.; Guo, L.; Ma, Y.; Zhao, R.; Li, X.; Shen, L.; Zhou, Z.; Kim, S.C.; Liu, J. Astragaloside IV blocks monocrotaline-induced pulmonary arterial hypertension by improving inflammation and pulmonary artery remodeling. Int. J. Mol. Med. 2021, 47, 595–606. [Google Scholar] [CrossRef]
- Sun, Y.; Lu, M.; Sun, T.; Wang, H. Astragaloside IV attenuates inflammatory response mediated by NLRP-3/calpain-1 is involved in the development of pulmonary hypertension. J. Cell Mol. Med. 2021, 25, 586–590. [Google Scholar] [CrossRef]
- Li, C.; Zhu, H.; Zhang, S.; Meng, F.; Li, S.; Li, G.; Zha, J.; Wu, S.; Zhu, L.; Dai, A. Astragaloside IV ameliorates pulmonary vascular remodeling in hypoxia-induced pulmonary hypertension by restraining the T follicular helper cell response and expanding T follicular regulatory cell response. Phytomedicine 2022, 102, 154171. [Google Scholar] [CrossRef]
- Yao, J.; Fang, X.; Zhang, C.; Yang, Y.; Wang, D.; Chen, Q.; Zhong, G. Astragaloside IV attenuates hypoxia-induced pulmonary vascular remodeling via the Notch signaling pathway. Mol. Med. Rep. 2021, 23, 89. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Jiao, G.; Chen, Y.; Han, M.; Wang, X.; Liu, W. Astragaloside IV attenuates chronic intermittent hypoxia-induced myocardial injury by modulating Ca(2+) homeostasis. Cell Biochem. Funct. 2020, 38, 710–720. [Google Scholar] [CrossRef]
- Mirzaei, H.; Shakeri, A.; Rashidi, B.; Jalili, A.; Banikazemi, Z.; Sahebkar, A. Phytosomal curcumin: A review of pharmacokinetic, experimental and clinical studies. Biomed. Pharmacother. 2017, 85, 102–112. [Google Scholar] [CrossRef]
- Li, K.X.; Wang, Z.C.; Machuki, J.O.; Li, M.Z.; Wu, Y.J.; Niu, M.K.; Yu, K.Y.; Lu, Q.B.; Sun, H.J. Benefits of Curcumin in the Vasculature: A Therapeutic Candidate for Vascular Remodeling in Arterial Hypertension and Pulmonary Arterial Hypertension? Front. Physiol. 2022, 13, 848867. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, W.; Zhu, F.; Wang, Q.; Yang, H.; Wu, J. Curcumin Improves Pulmonary Hypertension Rats by Regulating Mitochondrial Function. Biomed. Res. Int. 2021, 2021, 1078019. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.; Yousefvand, S.; Jamialahmadi, T.; Johnston, T.P.; Sahebkar, A. Protective Effects of Curcumin on Pulmonary Arterial Hypertension. Adv. Exp. Med. Biol. 2021, 1328, 213–221. [Google Scholar] [CrossRef]
- Kruangtip, O.; Chootip, K.; Temkitthawon, P.; Changwichit, K.; Chuprajob, T.; Changtam, C.; Suksamrarn, A.; Khorana, N.; Scholfield, C.N.; Ingkaninan, K. Curcumin analogues inhibit phosphodiesterase-5 and dilate rat pulmonary arteries. J. Pharm. Pharmacol. 2015, 67, 87–95. [Google Scholar] [CrossRef]
- Rice, K.M.; Manne, N.D.; Kolli, M.B.; Wehner, P.S.; Dornon, L.; Arvapalli, R.; Selvaraj, V.; Kumar, A.; Blough, E.R. Curcumin nanoparticles attenuate cardiac remodeling due to pulmonary arterial hypertension. Artif. Cells Nanomed. Biotechnol. 2016, 44, 1909–1916. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Ma, Z.; Zhang, X.; Zhou, F. Evaluation of Effect of Curcumin on Psychological State of Patients with Pulmonary Hypertension by Magnetic Resonance Image under Deep Learning. Contrast Media Mol. Imaging 2021, 2021, 9935754. [Google Scholar] [CrossRef]
- Hu, Y.; Chi, L.; Kuebler, W.M.; Goldenberg, N.M. Perivascular Inflammation in Pulmonary Arterial Hypertension. Cells 2020, 9, 2338. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Desai, A.A.; Black, S.M.; Tang, H. Cytokines, Chemokines, and Inflammation in Pulmonary Arterial Hypertension. Adv. Exp. Med. Biol. 2021, 1303, 275–303. [Google Scholar] [CrossRef]
- Klouda, T.; Yuan, K. Inflammation in Pulmonary Arterial Hypertension. Adv. Exp. Med. Biol. 2021, 1303, 351–372. [Google Scholar] [CrossRef]
- Mercurio, V.; Cuomo, A.; Naranjo, M.; Hassoun, P.M. Inflammatory Mechanisms in the Pathogenesis of Pulmonary Arterial Hypertension: Recent Advances. Compr. Physiol. 2021, 11, 1805–1829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Y.; Liang, M.T.; Gong, Y.; Yang, Y.; Bu, P.L.; Zhang, M.; Yao, H.C. HGF Reduces Disease Severity and Inflammation by Attenuating the NF-κB Signaling in a Rat Model of Pulmonary Artery Hypertension. Inflammation 2018, 41, 924–931. [Google Scholar] [CrossRef]
- Yang, J.M.; Zhou, R.; Zhang, M.; Tan, H.R.; Yu, J.Q. Betaine Attenuates Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats via Inhibiting Inflammatory Response. Molecules 2018, 23, 1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, U.H.; Suh, S.J.; Chang, H.W.; Son, J.K.; Lee, S.H.; Son, K.H.; Chang, Y.C.; Kim, C.H. Tanshinone IIA from Salvia miltiorrhiza BUNGE inhibits human aortic smooth muscle cell migration and MMP-9 activity through AKT signaling pathway. J. Cell Biochem. 2008, 104, 15–26. [Google Scholar] [CrossRef]
- Hu, G.l.; Song, Y.l.; Ke, S.l.; Cao, H.B.; Zhang, C.Y.; Deng, G.F.; Yang, F.; Zhou, S.H.; Liu, P.; Guo, X.; et al. Tanshinone IIA protects against pulmonary arterial hypertension in broilers. Poult Sci. 2017, 96, 11321138. [Google Scholar] [CrossRef]
- Langer, F.; Schramm, R.; Bauer, M.; Tscholl, D.; Kunihara, T.; Schäfers, H.J. Cytokine response to pulmonary thromboendarterectomy. Chest 2004, 126, 135–141. [Google Scholar] [CrossRef]
- Chun, C.; Yang, W.; Xueding, C.; Qi, Z.; Xiaoying, H.; Honglei, X.; Fangyou, Y.; Chan, C.; Yuanyuan, L.; Weixi, Z.; et al. Resveratrol downregulates acute pulmonary thromboembolism-induced pulmonary artery hypertension via p38 mitogen-activated protein kinase and monocyte chemoattractant protein-1 signaling in rats. Life Sci. 2012, 90, 721–727. [Google Scholar] [CrossRef]
- Lin, J.W.; Yang, L.H.; Ren, Z.C.; Mu, D.G.; Li, Y.Q.; Yan, J.P.; Wang, L.X.; Chen, C. Resveratrol downregulates TNF-α-induced monocyte chemoattractant protein-1 in primary rat pulmonary artery endothelial cells by P38 mitogen-activated protein kinase signaling. Drug Des. Devel Ther. 2019, 13, 1843–1853. [Google Scholar] [CrossRef] [Green Version]
- Groth, A.; Vrugt, B.; Brock, M.; Speich, R.; Ulrich, S.; Huber, L.C. Inflammatory cytokines in pulmonary hypertension. Respir Res. 2014, 15, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Wang, H.; Zhao, J.; Yan, J.; Meng, H.; Zhan, H.; Chen, L.; Yuan, L. Grape seed proanthocyanidin inhibits monocrotaline-induced pulmonary arterial hypertension via attenuating inflammation: In vivo and in vitro studies. J. Nutr. Biochem. 2019, 67, 72–77. [Google Scholar] [CrossRef]
- Sánchez-Gloria, J.L.; Martínez-Olivares, C.E.; Rojas-Morales, P.; Hernández-Pando, R.; Carbó, R.; Rubio-Gayosso, I.; Arellano-Buendía, A.S.; Rada, K.M.; Sánchez-Muñoz, F.; Osorio-Alonso, H. Anti-Inflammatory Effect of Allicin Associated with Fibrosis in Pulmonary Arterial Hypertension. Int. J. Mol. Sci. 2021, 22, 8600. [Google Scholar] [CrossRef]
- Toshner, M.; Church, C.; Harbaum, L.; Rhodes, C.; Villar Moreschi, S.S.; Liley, J.; Jones, R.; Arora, A.; Batai, K.; Desai, A.A.; et al. Mendelian randomisation and experimental medicine approaches to interleukin-6 as a drug target in pulmonary arterial hypertension. Eur. Respir J. 2022, 59, 2002463. [Google Scholar] [CrossRef] [PubMed]
- Florentin, J.; Zhao, J.; Tai, Y.Y.; Vasamsetti, S.B.; O’Neil, S.P.; Kumar, R.; Arunkumar, A.; Watson, A.; Sembrat, J.; Bullock, G.C.; et al. Interleukin-6 mediates neutrophil mobilization from bone marrow in pulmonary hypertension. Cell Mol. Immunol. 2021, 18, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Brock, M.; Trenkmann, M.; Gay, R.E.; Michel, B.A.; Gay, S.; Fischler, M.; Ulrich, S.; Speich, R.; Huber, L.C. Interleukin-6 modulates the expression of the bone morphogenic protein receptor type II through a novel STAT3-microRNA cluster 17/92 pathway. Circ. Res. 2009, 104, 1184–1191. [Google Scholar] [CrossRef] [Green Version]
- Scott, T.E.; Kemp-Harper, B.K.; Hobbs, A.J. Inflammasomes: A novel therapeutic target in pulmonary hypertension? Br. J. Pharmacol. 2019, 176, 1880–1896. [Google Scholar] [CrossRef]
- Ornatowski, W.; Lu, Q.; Yegambaram, M.; Garcia, A.E.; Zemskov, E.A.; Maltepe, E.; Fineman, J.R.; Wang, T.; Black, S.M. Complex interplay between autophagy and oxidative stress in the development of pulmonary disease. Redox. Biol. 2020, 36, 101679. [Google Scholar] [CrossRef]
- Joppa, P.; Petrásová, D.; Stancák, B.; Dorková, Z.; Tkácová, R. Oxidative stress in patients with COPD and pulmonary hypertension. Wien. Klin. Wochenschr. 2007, 119, 428–434. [Google Scholar] [CrossRef]
- Hoshikawa, Y.; Ono, S.; Suzuki, S.; Tanita, T.; Chida, M.; Song, C.; Noda, M.; Tabata, T.; Voelkel, N.F.; Fujimura, S. Generation of oxidative stress contributes to the development of pulmonary hypertension induced by hypoxia. J. Appl. Physiol. (1985) 2001, 90, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Luo, P.; Shi, W.; Guo, J.; Huo, S.; Yan, D.; Peng, L.; Zhang, C.; Lv, J.; Lin, L.; et al. S-Nitroso-L-Cysteine Ameliorated Pulmonary Hypertension in the MCT-Induced Rats through Anti-ROS and Anti-Inflammatory Pathways. Oxid. Med. Cell Longev. 2021, 2021, 6621232. [Google Scholar] [CrossRef] [PubMed]
- Brandt, M.; Giokoglu, E.; Garlapati, V.; Bochenek, M.L.; Molitor, M.; Hobohm, L.; Schönfelder, T.; Münzel, T.; Kossmann, S.; Karbach, S.H.; et al. Pulmonary Arterial Hypertension and Endothelial Dysfunction Is Linked to NADPH Oxidase-Derived Superoxide Formation in Venous Thrombosis and Pulmonary Embolism in Mice. Oxid. Med. Cell Longev. 2018, 2018, 1860513. [Google Scholar] [CrossRef] [PubMed]
- Suresh, K.; Servinsky, L.; Jiang, H.; Bigham, Z.; Yun, X.; Kliment, C.; Huetsch, J.; Damarla, M.; Shimoda, L.A. Reactive oxygen species induced Ca(2+) influx via TRPV4 and microvascular endothelial dysfunction in the SU5416/hypoxia model of pulmonary arterial hypertension. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, L893–L907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, S.; Boucherat, O. The ROS controversy in hypoxic pulmonary hypertension revisited. Eur. Respir J. 2018, 51, 1800276. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Pagano, P.J. Microvascular NADPH oxidase in health and disease. Free Radic. Biol. Med. 2017, 109, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Nisbet, R.E.; Graves, A.S.; Kleinhenz, D.J.; Rupnow, H.L.; Reed, A.L.; Fan, T.H.; Mitchell, P.O.; Sutliff, R.L.; Hart, C.M. The role of NADPH oxidase in chronic intermittent hypoxia-induced pulmonary hypertension in mice. Am. J. Respir Cell Mol. Biol. 2009, 40, 601–609. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Chang, Z.; Zhao, F.; Zhang, P.; Hao, Y.J.; Yan, L.; Liu, N.; Wang, J.L.; Bo, L.; Ma, P.; et al. Protective Effects of 18β-Glycyrrhetinic Acid on Monocrotaline-Induced Pulmonary Arterial Hypertension in Rats. Front. Pharmacol. 2019, 10, 13. [Google Scholar] [CrossRef]
- Zhang, Y.S.; He, L.; Liu, B.; Li, N.S.; Luo, X.J.; Hu, C.P.; Ma, Q.L.; Zhang, G.G.; Li, Y.J.; Peng, J. A novel pathway of NADPH oxidase/vascular peroxidase 1 in mediating oxidative injury following ischemia-reperfusion. Basic Res. Cardiol. 2012, 107, 266. [Google Scholar] [CrossRef]
- Liu, B.; Luo, X.J.; Yang, Z.B.; Zhang, J.J.; Li, T.B.; Zhang, X.J.; Ma, Q.L.; Zhang, G.G.; Hu, C.P.; Peng, J. Inhibition of NOX/VPO1 pathway and inflammatory reaction by trimethoxystilbene in prevention of cardiovascular remodeling in hypoxia-induced pulmonary hypertensive rats. J. Cardiovasc. Pharmacol. 2014, 63, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Zeng, Z.; Wang, X.; Yuan, T.; Wang, C.; Liu, D.; Guo, J.; Chen, Y. Deciphering the Mechanism of Wogonin, a Natural Flavonoid, on the Proliferation of Pulmonary Arterial Smooth Muscle Cells by Integrating Network Pharmacology and In Vitro Validation. Curr. Issues Mol. Biol. 2023, 45, 555–570. [Google Scholar] [CrossRef]
- Harris, C.; Hansen, J.M. Nrf2-mediated resistance to oxidant-induced redox disruption in embryos. Birth Defects Res. B Dev. Reprod. Toxicol. 2012, 95, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Li, Y.; Zhang, B.; Wang, Y.; Liu, Y.; Luo, Y.; Niu, W.; Dong, M.; Liu, M.; Dong, H.; et al. Resveratrol alleviate hypoxic pulmonary hypertension via anti-inflammation and anti-oxidant pathways in rats. Int. J. Med. Sci. 2016, 13, 942–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, M.; Capuano, V.; Olschewski, A.; Sabourin, J.; Nagaraj, C.; Girerd, B.; Weatherald, J.; Humbert, M.; Antigny, F. Ion Channels in Pulmonary Hypertension: A Therapeutic Interest? Int. J. Mol. Sci. 2018, 19, 3162. [Google Scholar] [CrossRef] [Green Version]
- Lai, N.; Lu, W.; Wang, J. Ca(2+) and ion channels in hypoxia-mediated pulmonary hypertension. Int. J. Clin. Exp. Pathol. 2015, 8, 1081–1092. [Google Scholar] [PubMed]
- Mondéjar-Parreño, G.; Cogolludo, A.; Perez-Vizcaino, F. Potassium (K(+)) channels in the pulmonary vasculature: Implications in pulmonary hypertension Physiological, pathophysiological and pharmacological regulation. Pharmacol. Ther. 2021, 225, 107835. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dong, M.Q.; Liu, M.L.; Xu, D.Q.; Luo, Y.; Zhang, B.; Liu, L.L.; Xu, M.; Zhao, P.T.; Gao, Y.Q.; et al. Tanshinone IIA modulates pulmonary vascular response to agonist and hypoxia primarily via inhibiting Ca2+ influx and release in normal and hypoxic pulmonary hypertension rats. Eur. J. Pharmacol. 2010, 640, 129–138. [Google Scholar] [CrossRef]
- Hong, Z.; Weir, E.K.; Nelson, D.P.; Olschewski, A. Subacute hypoxia decreases voltage-activated potassium channel expression and function in pulmonary artery myocytes. Am. J. Respir Cell Mol. Biol. 2004, 31, 337–343. [Google Scholar] [CrossRef]
- Huang, Y.F.; Liu, M.L.; Dong, M.Q.; Yang, W.C.; Zhang, B.; Luan, L.L.; Dong, H.Y.; Xu, M.; Wang, Y.X.; Liu, L.L.; et al. Effects of sodium tanshinone II A sulphonate on hypoxic pulmonary hypertension in rats in vivo and on Kv2.1 expression in pulmonary artery smooth muscle cells in vitro. J. Ethnopharmacol. 2009, 125, 436–443. [Google Scholar] [CrossRef]
- Wisutthathum, S.; Demougeot, C.; Totoson, P.; Adthapanyawanich, K.; Ingkaninan, K.; Temkitthawon, P.; Chootip, K. Eulophia macrobulbon extract relaxes rat isolated pulmonary artery and protects against monocrotaline-induced pulmonary arterial hypertension. Phytomedicine 2018, 50, 157–165. [Google Scholar] [CrossRef]
- Zhang, R.; Li, Z.; Liu, C.; Yang, Q.; Lu, D.; Ge, R.L.; Ma, S.; Li, Z. Pretreatment with the active fraction of Rhodiola tangutica (Maxim.) S.H. Fu rescues hypoxia-induced potassium channel inhibition in rat pulmonary artery smooth muscle cells. J. Ethnopharmacol. 2022, 283, 114734. [Google Scholar] [CrossRef]
- Guignabert, C.; Dorfmuller, P. Pathology and pathobiology of pulmonary hypertension. Semin. Respir Crit. Care Med. 2013, 34, 551–559. [Google Scholar] [CrossRef]
- Li, M.X.; Jiang, D.Q.; Wang, Y.; Chen, Q.Z.; Ma, Y.J.; Yu, S.S.; Wang, Y. Signal Mechanisms of Vascular Remodeling in the Development of Pulmonary Arterial Hypertension. J. Cardiovasc. Pharmacol. 2016, 67, 182–190. [Google Scholar] [CrossRef]
- Zhu, Z.; Xu, Y.; Zou, H.; Zhang, Z.; Ni, W.; Chen, S. Effects of puerarin on pulmonary vascular remodeling and protein kinase C-alpha in chronic cigarette smoke exposure smoke-exposed rats. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2008, 28, 27–32. [Google Scholar] [CrossRef]
- Piantadosi, C.A.; Suliman, H.B. Mitochondrial Dysfunction in Lung Pathogenesis. Annu. Rev. Physiol. 2017, 79, 495–515. [Google Scholar] [CrossRef]
- He, Y.; Fang, X.; Shi, J.; Li, X.; Xie, M.; Liu, X. Apigenin attenuates pulmonary hypertension by inducing mitochondria-dependent apoptosis of PASMCs via inhibiting the hypoxia inducible factor 1α-KV1.5 channel pathway. Chem. Biol. Interact. 2020, 317, 108942. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zou, L.; Yu, X.; Chen, M.; Guo, R.; Cai, H.; Yao, D.; Xu, X.; Chen, Y.; Ding, C.; et al. Salidroside attenuates chronic hypoxia-induced pulmonary hypertension via adenosine A2a receptor related mitochondria-dependent apoptosis pathway. J. Mol. Cell Cardiol. 2015, 82, 153–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Cai, H.; Yu, C.; Wu, P.; Fu, Y.; Xu, X.; Fan, R.; Xu, C.; Chen, Y.; Wang, L.; et al. Salidroside exerts protective effects against chronic hypoxia-induced pulmonary arterial hypertension via AMPKα1-dependent pathways. Am. J. Transl. Res. 2016, 8, 12–27. [Google Scholar]
- Tang, W.L.; Guo, H.; Yang, J.; Chen, B.; Wang, X. Suppression of tissue inhibitors of metalloproteinases may reverse severe pulmonary arterial hypertension. Cytotherapy 2011, 13, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Yan, S.; Chen, M.; Chen, A.; Yao, D.; Xu, X.; Cai, X.; Wang, L.; Huang, X. Effects of baicalin on collagen Ι and collagen ΙΙΙ expression in pulmonary arteries of rats with hypoxic pulmonary hypertension. Int. J. Mol. Med. 2015, 35, 901–908. [Google Scholar] [CrossRef] [Green Version]
- Salter, R.C.; Ashlin, T.G.; Kwan, A.P.; Ramji, D.P. ADAMTS proteases: Key roles in atherosclerosis? J. Mol. Med. (Berl) 2010, 88, 1203–1211. [Google Scholar] [CrossRef]
- Li, X.W.; Wang, X.M.; Li, S.; Yang, J.R. Effects of chrysin (5,7-dihydroxyflavone) on vascular remodeling in hypoxia-induced pulmonary hypertension in rats. Chin. Med. 2015, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, R.D.; Chartan, C.A.; Mourani, P.M. Intensive care management of right ventricular failure and pulmonary hypertension crises. Pediatr. Pulmonol. 2021, 56, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Zangiabadi, A.; De Pasquale, C.G.; Sajkov, D. Pulmonary hypertension and right heart dysfunction in chronic lung disease. Biomed. Res. Int. 2014, 2014, 739674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmeißer, A.; Rauwolf, T.; Groscheck, T.; Fischbach, K.; Kropf, S.; Luani, B.; Tanev, I.; Hansen, M.; Meißler, S.; Schäfer, K.; et al. Predictors and prognosis of right ventricular function in pulmonary hypertension due to heart failure with reduced ejection fraction. ESC Heart Fail 2021, 8, 2968–2981. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Garza, E.; Bernal-Ramírez, J.; Jerjes-Sánchez, C.; Lozano, O.; Acuña-Morín, E.; Vanoye-Tamez, M.; Ramos-González, M.R.; Chapoy-Villanueva, H.; Pérez-Plata, L.; Sánchez-Trujillo, L.; et al. Resveratrol Prevents Right Ventricle Remodeling and Dysfunction in Monocrotaline-Induced Pulmonary Arterial Hypertension with a Limited Improvement in the Lung Vasculature. Oxid. Med. Cell. Longev. 2020, 2020, 1841527. [Google Scholar] [CrossRef]
- Li, L.; Dong, P.; Hou, C.; Cao, F.; Sun, S.; He, F.; Song, Y.; Li, S.; Bai, Y.; Zhu, D. Hydroxysafflor yellow A (HSYA) attenuates hypoxic pulmonary arterial remodelling and reverses right ventricular hypertrophy in rats. J. Ethnopharmacol. 2016, 186, 224–233. [Google Scholar] [CrossRef]
- Sun, X.; Ku, D.D. Allicin in garlic protects against coronary endothelial dysfunction and right heart hypertrophy in pulmonary hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H2431–H2438. [Google Scholar] [CrossRef] [Green Version]
- Ahmadipour, B.; Kalantar, M.; Schreurs, N.M.; Raza, S.H.A.; Khan, R.; Khan, S.; Abd El-Aziz, A.H.; Memon, S.; Ullah, I.; Samira, A. Flavonoid bioactive compounds of hawthorn extract can promote growth, regulate electrocardiogram waves, and improve cardiac parameters of pulmonary hypertensive chickens. Poult. Sci. 2020, 99, 974–980. [Google Scholar] [CrossRef]
- Yan, L.; Huang, H.; Tang, Q.Z.; Zhu, L.H.; Wang, L.; Liu, C.; Bian, Z.Y.; Li, H. Breviscapine protects against cardiac hypertrophy through blocking PKC-alpha-dependent signaling. J. Cell Biochem. 2010, 109, 1158–1171. [Google Scholar] [CrossRef] [PubMed]
- Kuriyama, S.; Morio, Y.; Toba, M.; Nagaoka, T.; Takahashi, F.; Iwakami, S.; Seyama, K.; Takahashi, K. Genistein attenuates hypoxic pulmonary hypertension via enhanced nitric oxide signaling and the erythropoietin system. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 306, L996–L1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, W.; He, W.; Li, Y.; Chen, Y.; Liang, J.; Lei, H.; Fu, L.; Chen, Y.; Ren, N.; Jiang, Q.; et al. Efficacy and safety of novel-targeted drugs in the treatment of pulmonary arterial hypertension: A Bayesian network meta-analysis. Drug Deliv. 2021, 28, 1007–1019. [Google Scholar] [CrossRef]
- Sharma, S.; Umar, S.; Centala, A.; Eghbali, M. Role of miR206 in genistein-induced rescue of pulmonary hypertension in monocrotaline model. J. Appl. Physiol. (1985) 2015, 119, 1374–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Natural Products | Sources | Model | Dose | mPVP | PVR | RVH | Mechanisms | Phase | Classification |
|---|---|---|---|---|---|---|---|---|---|
| Sodium tanshinone II sulfonate A | Salvia miltiorrhiza Bung | HPH rats | 10 mg/kg, 30 mg/kg, 3 weeks | + | + | + | IL-6↓IL-8↓TNF-α↓PI3K/AKT/mTOR↓ [38] | Clinical trial Phase 3 | Anti-inflammatory |
| Magnesium lithospermate B | Salvia miltiorrhiza Bunge | HPH rats | 5, 15 mg/kg, 30 days | + | + | + | EndMT↓HIF-1α↓NF-κB↓MCP-1↓PCNA↓CDK4↓CyclinD1↓ROCK1, 2↓ [45] | Preclinical trials | |
| Ligustrazine | Rhizoma Chuanxiong | MCT-induced PH rats | 40, 80, 160 mg/kg, 4 weeks | + | + | + | Inhibit inflammation by regulating the PI3K/AKT [50] | Clinical trial Phase 0 | |
| Resveratrol | Grapes, red wine, peanuts | MCT-induced PH rats | 25 mg/kg/, 3 weeks | + | + | + | TNF-α↓IL-1β↓IL-6↓PDGF-α/β↓ [59] | Preclinical trials | |
| Resveratrol | Grapes, red wine, peanuts | MCT-induced PH rats | 25 mg/kg, 4 weeks | + | + | + | SphK1/S1P/NF-κB↓ [60] | Preclinical trials | |
| Baicalin | Scutellaria baicalensis Georgi | MCT-induced PH rats | 100 mg/kg, 6 weeks | + | + | + | Regulate the TNF-α/BMPR2 [67] | Preclinical trials | |
| Baicalin | Scutellaria baicalensis Georgi | MCT-induced PH rats | 20, 100, 200 mg/kg, 29 days | + | + | + | AKT/ERK/NF-κB↓ [69] | Preclinical trials | |
| Baicalein | Scutellaria baicalensis Georgi | MCT-induced PH rats | 50,100 mg/kg, 4 weeks | + | + | + | IL-6↓TNF-α↓IL-1β↓ MAPK↓NF-κB↓ [72] | Preclinical trials | |
| Baicalein | Scutellaria baicalensis Georgi | MCT-induced PH rats | 50,100 mg/kg, 4 weeks | + | + | + | NF-κB-BMPR2↓EndMT↓ [73] | Preclinical trials | |
| Astragaloside IV | Astragalus membranaceus | MCT-induced PH rats | 10, 30 mg/kg, 3 weeks | + | + | + | TNF-α↓IL-1β↓HIF-1α↓VEGF↓ [89] | Preclinical trials | |
| Astragaloside IV | Astragalus membranaceus | MCT-induced PH rats | 40 mg/kg, 80 mg/kg, 4 weeks | + | + | + | NLRP-3/calpain-1↓Caspase-1↓ASC↓IL-18↓IL-1β↓ [90] | Preclinical trials | |
| Betaine | Lycium barbarum | MCT-induced PH rats | 100, 200, 400 mg/kg, 6 weeks | + | + | + | MCP-1↓ET-1↓NF-κB↓TNF-α↓IL-1β↓ [107] | Preclinical trials | |
| Grape seed proanthocyanidin | Grape seeds | MCT-induced PH rats | 10 mL/kg, 3 weeks | + | + | + | NF-κB↓IL-1β↓IL-6↓TNF-α↓ [114] | Preclinical trials | |
| Allicin | Allium sativum L. | MCT-induced PH rats | 16 mg/kg, 4 weeks | + | + | + | TNF-α↓IL-6↓IL-1β↓CD68↓NFκB p65↓Iκβ↓TGF-β↓α-SMA↓ [115] | Preclinical trials | |
| Salvianolic acid A | Salvia miltiorrhiza Bunge | MCT-induced PH rats | 0.3, 1, 3 mg/kg, 4 weeks | + | + | + | Nrf2/HO-1↑ROS↓TGFβ1↓EndMT↓ [42] | Preclinical trials | Oxidative stress |
| Magnesium lithospermate B | Salvia miltiorrhiza Bunge | HPH rats | 5, 15 mg/kg, 3 weeks | + | + | + | NOX/ROS/ERK↓NOX2↓NOX4↓ [43] | Preclinical trials | |
| Tetramethylpyrazine | Rhizoma Chuanxiong | MCT-induced PH rats | 5 mg/kg, 4 weeks | + | + | ROS/iNOS/PKG↓ [53] | Clinical trial Phase 0 | ||
| Resveratrol | Grapes, red wine, peanuts | MCT-induced PH rats | 25 mg/kg/, 3 weeks | + | + | + | eNOs↑NOX2↓NOX4↓ [59] | Preclinical trials | |
| Resveratrol | Grapes, red wine, peanuts | HPH rats | 40 mg/kg, 4 weeks | + | + | + | MAPK/ERK1↓PI3K/AKT↓HIF-1 α↓Nrf-2/Trx-1↓ [134] | Preclinical trials | |
| Trimethoxystilbene | Resveratrol analog | HPH rats | 5, 10 mg/kg, 4 weeks | + | + | + | NOX/VPO1↓ [131] | Preclinical trials | |
| 18β-glycyrrhetinic acid | Radix glycyrrhizas | MCT-induced PH rats | 25, 50,100 mg/kg, 3 weeks | + | + | + | Nox2↓Nox4↓ [129] | Preclinical trials | |
| Genistein | Soybeans | CHPH rats | 60 mg/kg, 3 weeks | + | + | + | EPO/EPOR↑NO↑ [162] | Preclinical trials | |
| Tanshinone IIA | Salvia miltiorrhiza Bunge | CHPH rats | 10 mg/kg, 4 weeks | + | + | + | KV2.1↑KV1.5↑ [33] | Clinical trial Phase 3 | Ion channels |
| Sodium tanshinone IIA sulfonate | Salvia miltiorrhiza Bunge | CHPH and MCT-induced PH rats | 10 mg/kg, 3 weeks | + | + | + | SOCE↓[Ca2+]i↓TRPC1↓TRPC6↓ [34] | Clinical trial Phase 3 | |
| Sodium tanshinone IIA sulfonate | Salvia miltiorrhiza Bunge | HPH rats | 30 mg/kg, 3 weeks | + | + | + | TRPC↓SOCE↓[Ca2+]i↓PKG-PPAR-γ↑ [35] | Clinical trial Phase 3 | |
| Sodium tanshinone IIA sulfonate | Salvia milti-orrhiza Bunge | HPH rats | 10 mg/kg, 3 weeks | + | + | + | KV2.1↑ [140] | Clinical trial Phase 3 | |
| Tetramethylpyrazine | Rhizoma Chuanxiong | HPH, MCT-PH rats | 100 mg/kg, 16 weeks | + | + | Inhibition of intracellular calcium homeostasis [49] | Clinical trial Phase 0 | ||
| Sodium tanshinone II sulfonate A | Salvia miltiorrhiza Bunge | HPH rats | 10 mg/kg; 30 mg/kg, 3 weeks | + | + | + | PI3K/AKT/mTOR↓Autophagy↑Bcl-2↓Bax↑ [38] | Clinical trial Phase 3 | Apoptotic resistance |
| Astragaloside IV | Astragalus membranaceus | MCT-induced PH rats | 10, 30 mg/kg, 3 weeks | + | + | + | Bcl-2↓ERK↓HIF-1α↓VEGF↓Caspase-3↑Caspase-9↑Bax↑ [89] | Preclinical trials | |
| Apigenin | Apium graveolens L. var. Dulce DC. | CHPH rats | 50, 100 mg/kg, 4 weeks | + | + | Cytochrome C ↑Bax↑Bcl-2↓Caspase-3↑Caspase-9↑HIF-1α-KV1.5↓ [147] | Preclinical trials | ||
| Salidroside | Rhodiola rosea | CHPH rats | 16, 32, 64 mg/kg, 4 weeks | + | + | + | Bax↑Bcl-2↓Caspase-9↑Cytochrome C↑A2aR↑ [148] | Preclinical trials | |
| Salidroside | Rhodiola rosea | HPH rats | 2, 8, 32 mg/kg, 4 weeks | + | + | Regulate the AMPKα1-P53-Bax/Bcl-2-caspase 9-caspase 3 [149] | Preclinical trials | ||
| Baicalin | Scutellaria baicalensis Georgi | HPH rats | 30 mg/kg, 4 weeks | + | + | + | P38 MAPK/MMP-9↓ [68] | Preclinical trials | Collagen deposition |
| Baicalin | Scutellaria baicalensis Georgi | HPH rats | 30 mg/kg, 4 weeks | + | + | + | ADAMTS-1↑Collagen I↓Collagen III↓ [151] | Preclinical trials | |
| Chrysin | Oroxylum indicum (L.) Vent. honey, and propolis | HPH rats | 50, 100 mg/kg, 4 weeks | + | + | + | Collagen I↓Collagen III↓NOX4↓ [153] | Preclinical trials | |
| Resveratrol | Grapes, red wine, peanuts | MCT-induced PH rats | 20 mg/kg, 6 weeks | + | + | SIRT3↑SERCA↑, prevent right ventricle dysfunction [62] | Preclinical trials | Right ventricle protection | |
| Resveratrol | Grapes, red wine, peanuts | MCT-induced PH rats | 20 mg/kg, 6 weeks | + | + | Sirtuin 1↑, improvement of right ventricle and isolated cardiomyocyte [157] | Preclinical trials | ||
| Curcumin | Curcuma longa L. | MCT-induced PH rats | 50 mg/kg, 4 weeks | + | + | TNF-α↓IL-1β↓Nitrotyrosine↓Fibronectin↓Myosin heavy chain-β↓, attenuate cardiac remodeling [99] | Preclinical trials | ||
| Hydroxysafflor yellow A | Carthamus tinctorius L. | HPH rats | 25, 50, 75,100 mg/kg, 9 days | + | + | + | PCNA↓Ki67↓, reverse right ventricular hypertrophy [158] | Preclinical trials |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Z.; Wang, X.; Cui, L.; Wang, H.; Guo, J.; Chen, Y. Natural Products for the Treatment of Pulmonary Hypertension: Mechanism, Progress, and Future Opportunities. Curr. Issues Mol. Biol. 2023, 45, 2351-2371. https://doi.org/10.3390/cimb45030152
Zeng Z, Wang X, Cui L, Wang H, Guo J, Chen Y. Natural Products for the Treatment of Pulmonary Hypertension: Mechanism, Progress, and Future Opportunities. Current Issues in Molecular Biology. 2023; 45(3):2351-2371. https://doi.org/10.3390/cimb45030152
Chicago/Turabian StyleZeng, Zuomei, Xinyue Wang, Lidan Cui, Hongjuan Wang, Jian Guo, and Yucai Chen. 2023. "Natural Products for the Treatment of Pulmonary Hypertension: Mechanism, Progress, and Future Opportunities" Current Issues in Molecular Biology 45, no. 3: 2351-2371. https://doi.org/10.3390/cimb45030152
APA StyleZeng, Z., Wang, X., Cui, L., Wang, H., Guo, J., & Chen, Y. (2023). Natural Products for the Treatment of Pulmonary Hypertension: Mechanism, Progress, and Future Opportunities. Current Issues in Molecular Biology, 45(3), 2351-2371. https://doi.org/10.3390/cimb45030152