
Temperature-Promoted Giant Unilamellar Vesicle (GUV) Aggregation: A Way of Multicellular Formation

Abstract
:1. Introduction
2. Materials and Methods
2.1. GUVs Preparation
2.2. MLVs Preparation
2.3. GUVs Solutions Incubation and Imaging
2.4. Zeta Potential Measurements
2.5. ATR-FTIR
2.6. Hydrolysis of Phospholipid Molecules
2.7. GUVs Solution pH Measurement
2.8. Date Analysis
3. Results
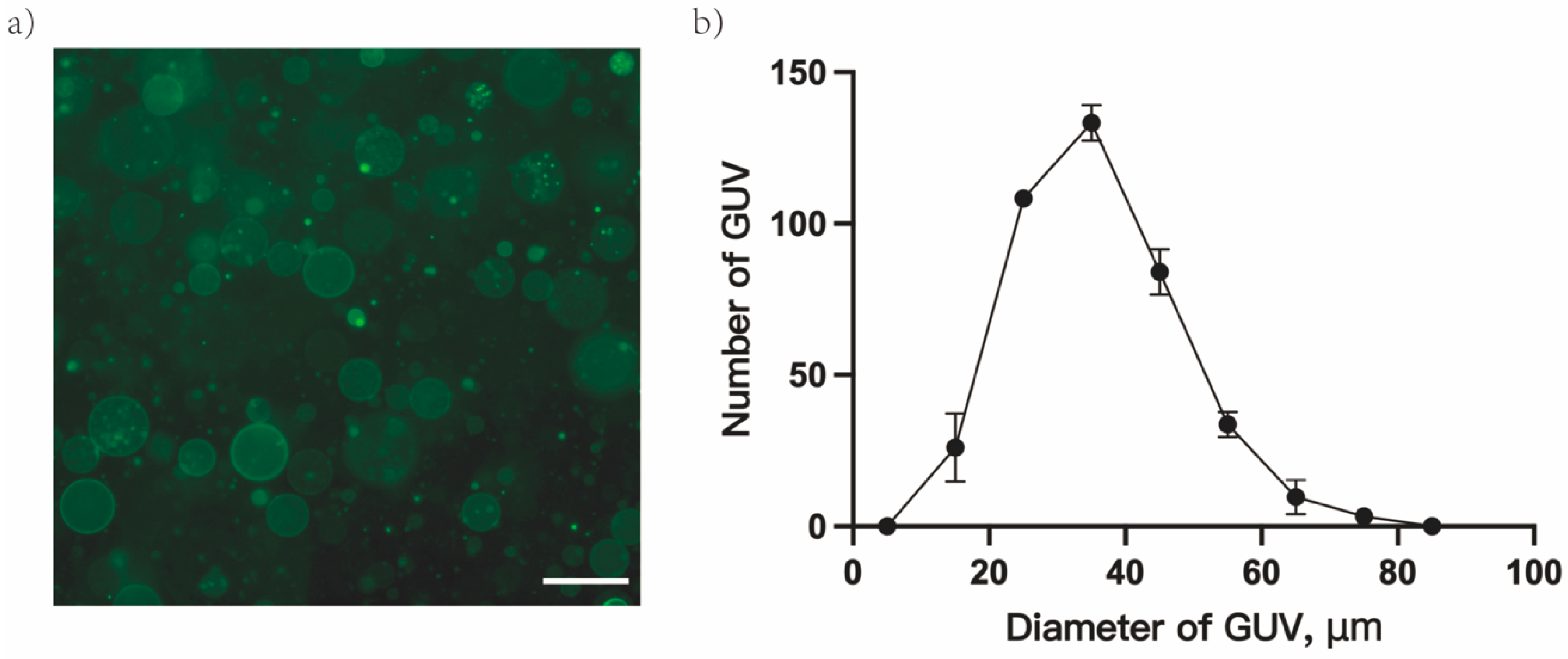
3.1. Characterization of GUVs
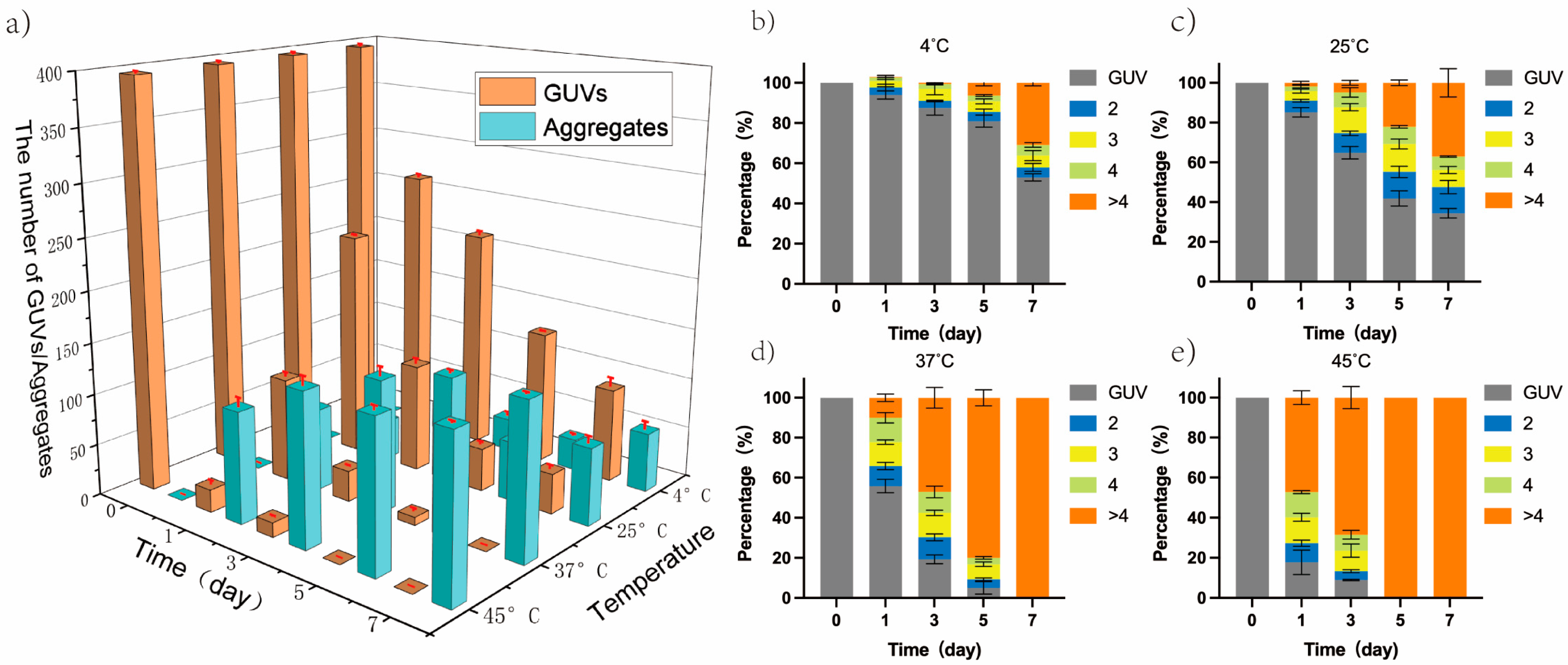
3.2. GUVs Aggregation by Increasing Temperature
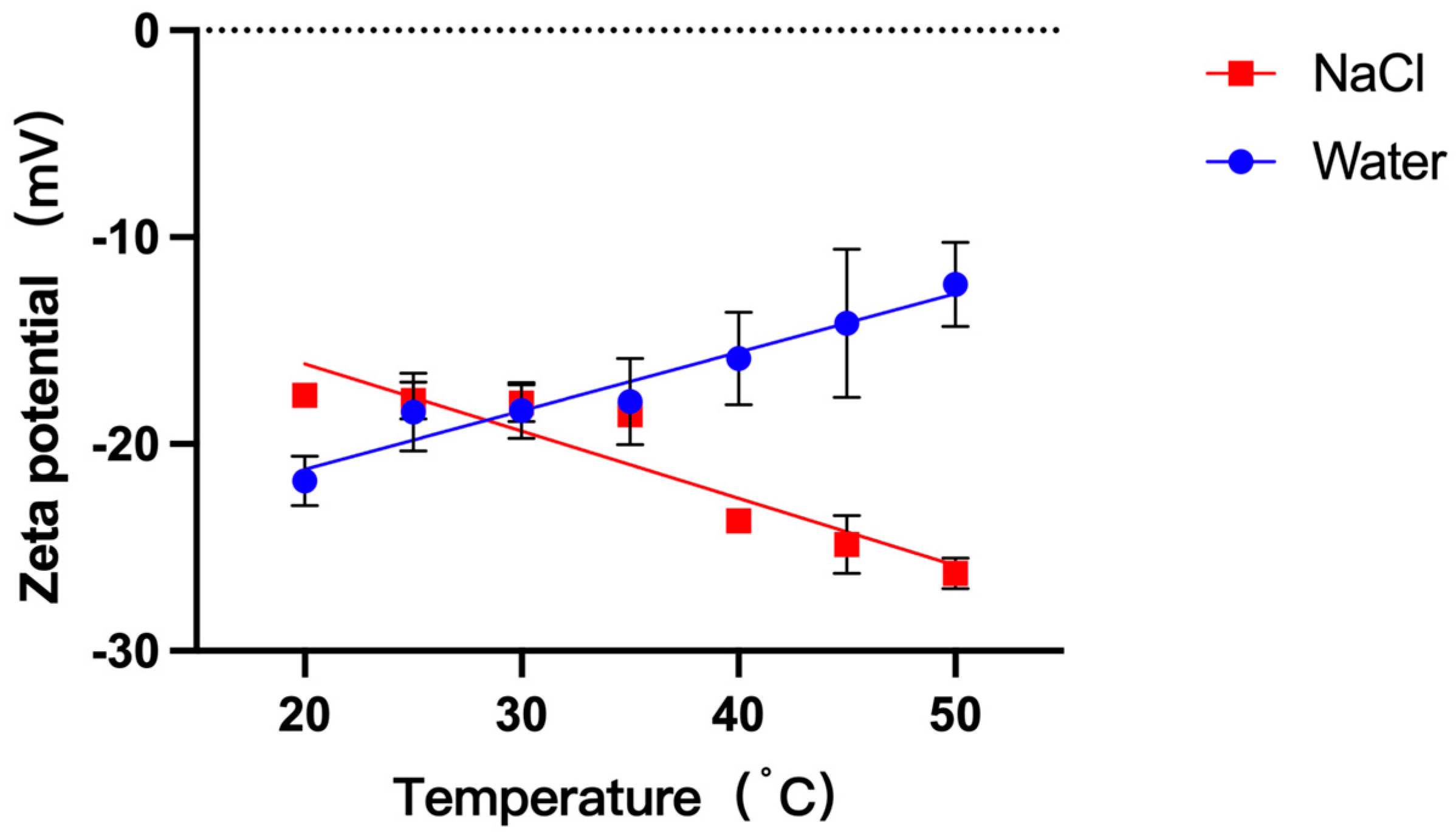
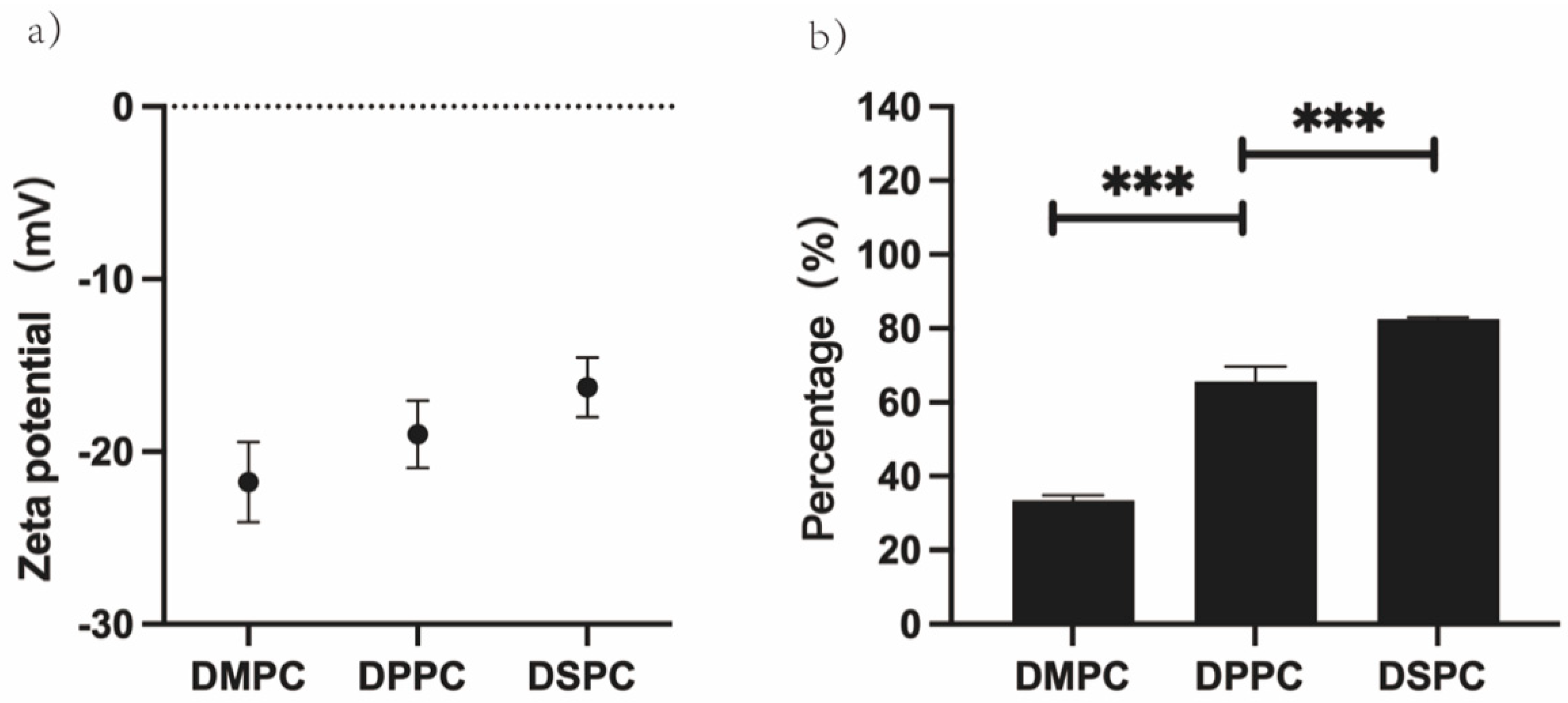
3.3. Effects of Increasing Temperature on GUVs Zeta Potential
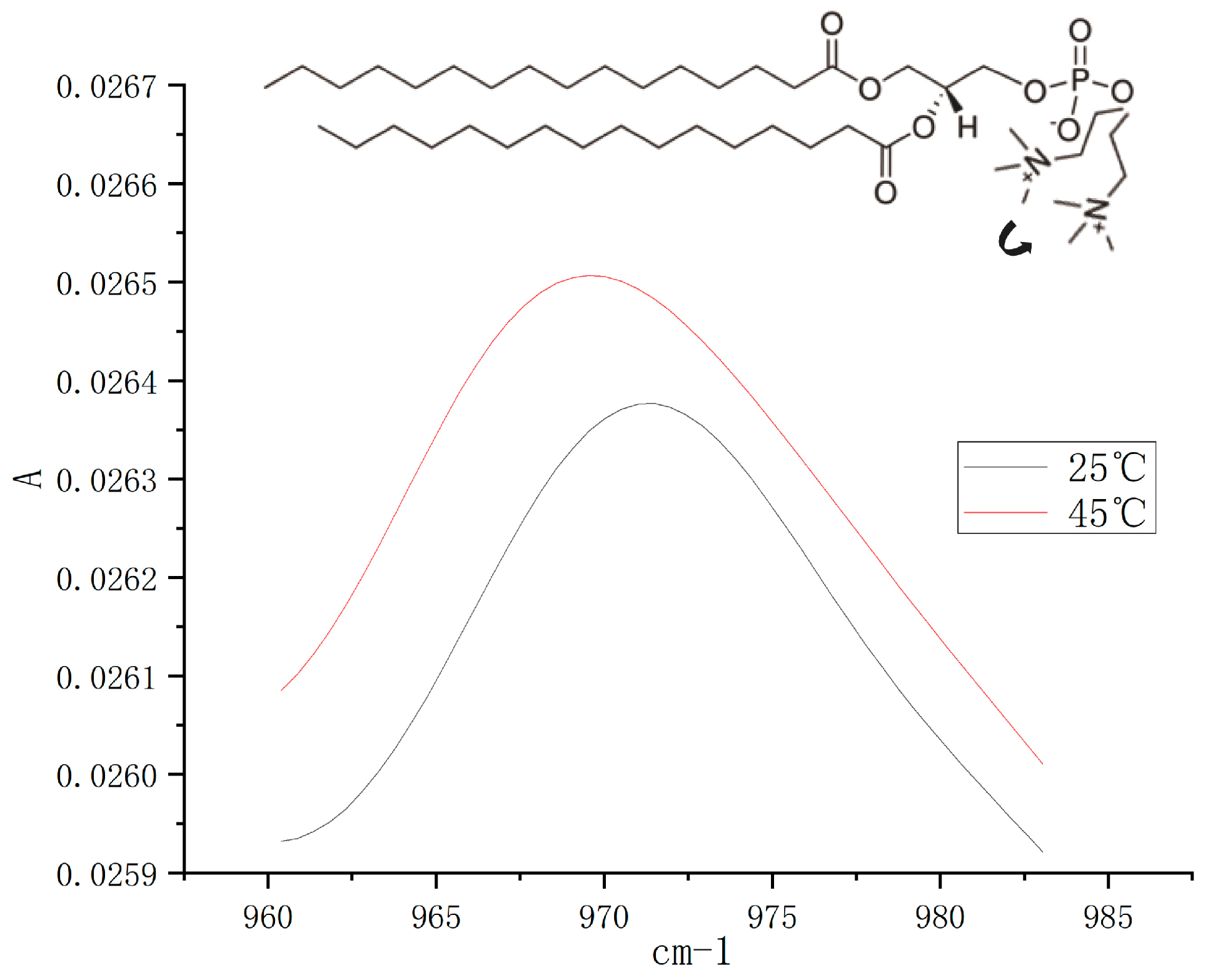
3.4. Effect of Increasing Temperature on the Orientation of the Phospholipid Headgroup
3.5. Effect of Zeta Potential on the Degree of GUVs Aggregation
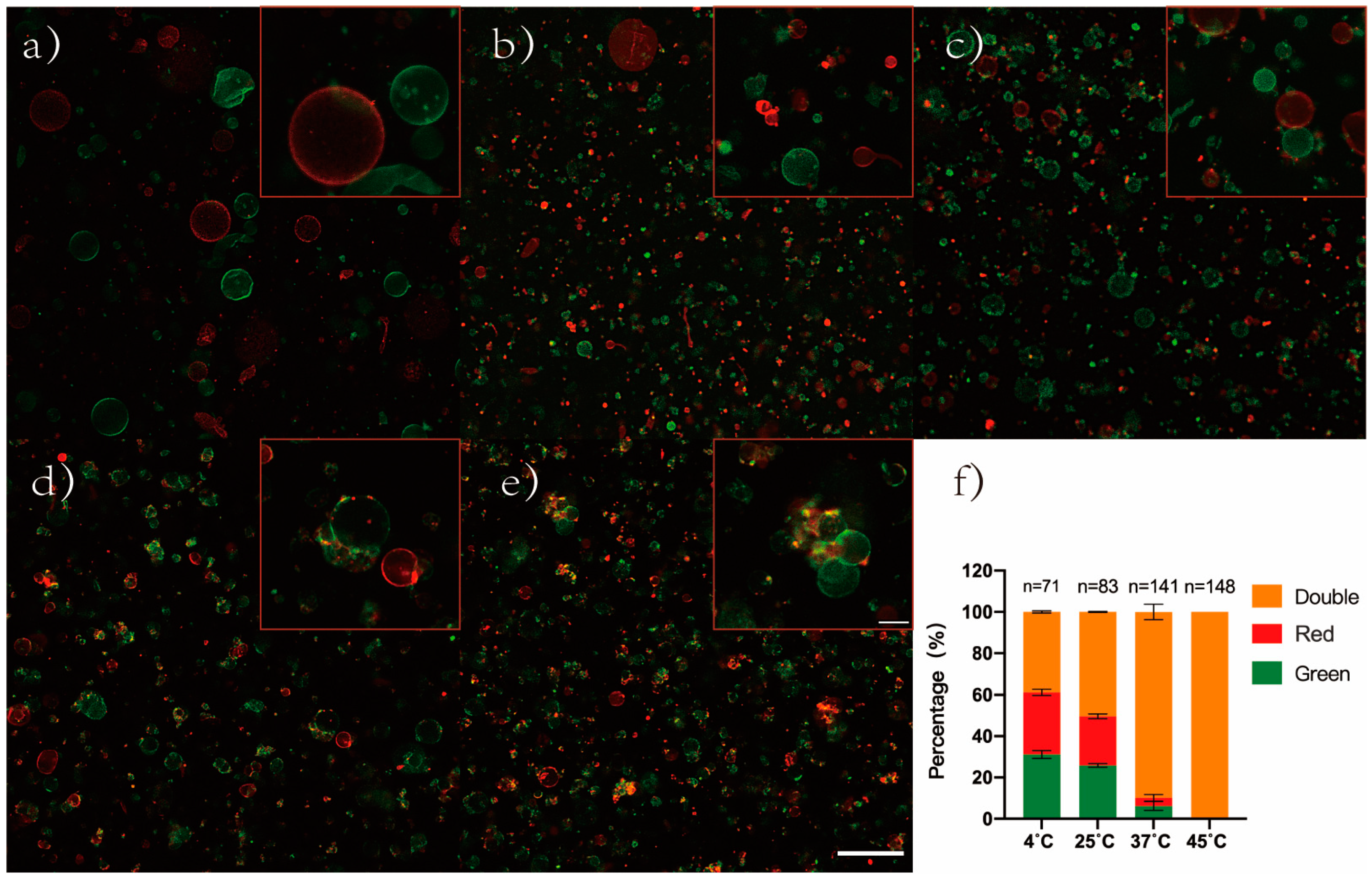
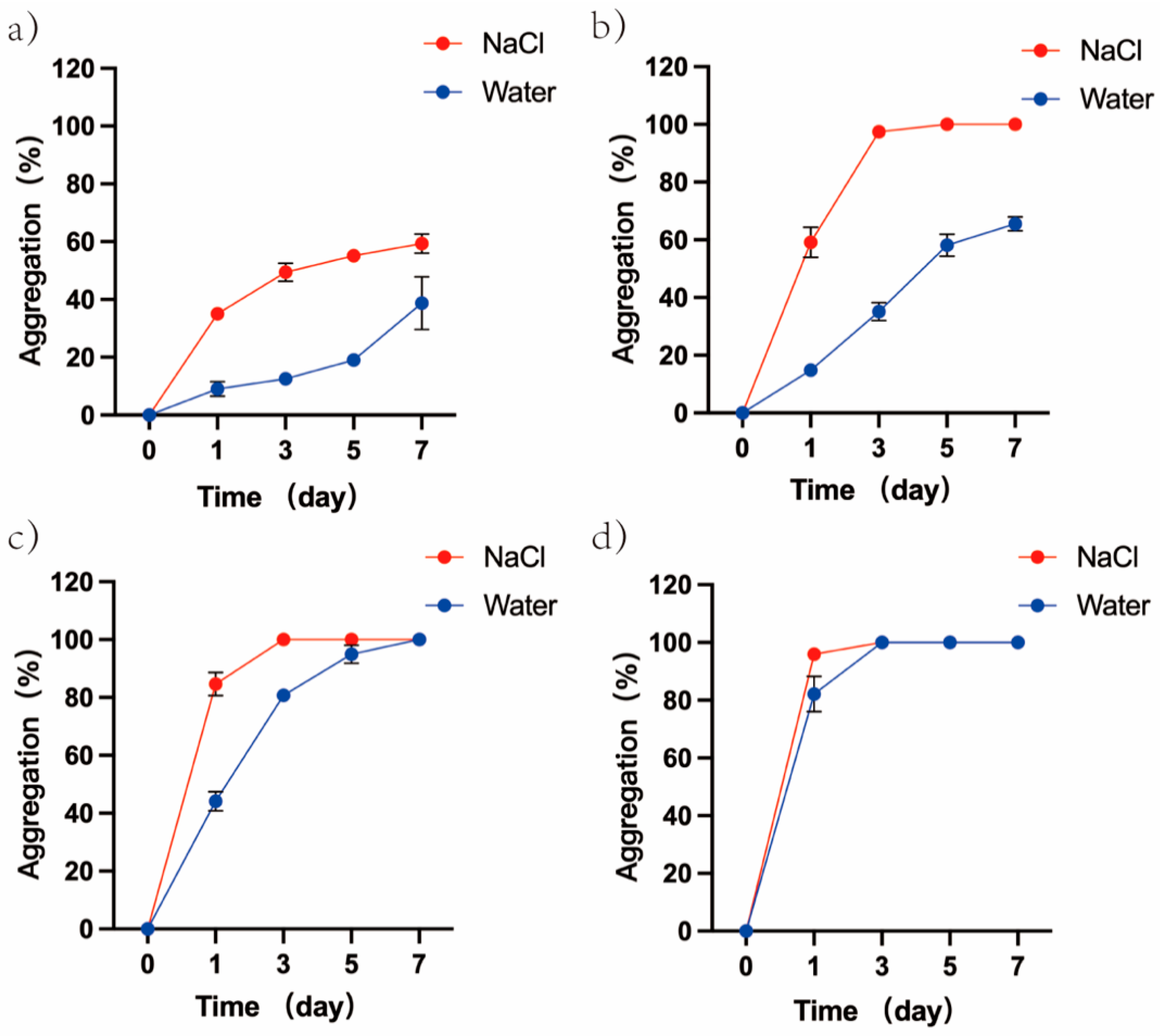
3.6. Effect of Ion on the Degree of GUVs Aggregation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niklas, K.J.; Wayne, R.; Benítez, M.; Newman, S.A. Polarity, Planes of Cell Division, and the Evolution of Plant Multicellularity. Protoplasma 2019, 256, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Altmayer, S.; Köhler, L.; Bielytskyi, P.; Gärtner, W.; Matysik, J.; Wiebeler, C.; Song, C. Light- and PH-Dependent Structural Changes in Cyanobacteriochrome AnPixJg2. Photochem. Photobiol. Sci. 2022, 21, 447–469. [Google Scholar] [CrossRef]
- Sogabe, S.; Hatleberg, W.L.; Kocot, K.M.; Say, T.E.; Stoupin, D.; Roper, K.E.; Fernandez-Valverde, S.L.; Degnan, S.M.; Degnan, B.M. Pluripotency and the Origin of Animal Multicellularity. Nature 2019, 570, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Cooney, D.B.; Rossine, F.W.; Morris, D.H.; Levin, S.A. A PDE Model for Protocell Evolution and the Origin of Chromosomes via Multilevel Selection. Bull. Math. Biol. 2022, 84, 109. [Google Scholar] [CrossRef]
- Godino, E.; López, J.N.; Foschepoth, D.; Cleij, C.; Doerr, A.; Castellà, C.F.; Danelon, C. De Novo Synthesized Min Proteins Drive Oscillatory Liposome Deformation and Regulate FtsA-FtsZ Cytoskeletal Patterns. Nat. Commun. 2019, 10, 4969. [Google Scholar] [CrossRef]
- Ratcliff, W.C.; Herron, M.D.; Howell, K.; Pentz, J.T.; Rosenzweig, F.; Travisano, M. Experimental Evolution of an Alternating Uni- and Multicellular Life Cycle in Chlamydomonas Reinhardtii. Nat. Commun. 2013, 4, 2742. [Google Scholar] [CrossRef]
- Grainger, T.N.; Levine, J.M. Rapid Evolution of Life-history Traits in Response to Warming, Predation and Competition: A Meta-analysis. Ecol. Lett. 2022, 25, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Nandy, G.; Aditya, G. Temperature Dependent Variations of Life History Traits of the Land Snail Allopeas Gracile (Hutton, 1834) (Gastropoda: Subulinidae). J. Therm. Biol. 2022, 108, 103297. [Google Scholar] [CrossRef] [PubMed]
- Boussau, B.; Blanquart, S.; Necsulea, A.; Lartillot, N.; Gouy, M. Parallel Adaptations to High Temperatures in the Archaean Eon. Nature 2008, 456, 942–945. [Google Scholar] [CrossRef]
- Ragland, G.J.; Kingsolver, J.G. The Effect of Fluctuating Temperatures on Ectotherm Life-History Traits: Comparisons among Geographic Populations of Wyeomyia Smithii. Evol. Ecol. Res. 2008, 10, 29–44. [Google Scholar]
- Ma, X.; Zhang, J.; Zhang, Y.; Liu, J. Adsorption Promoted Aggregation-Induced Emission Showing Strong Dye Lateral Interactions. Langmuir 2019, 35, 16304–16311. [Google Scholar] [CrossRef] [PubMed]
- Villamil Giraldo, A.-M.; Eriksson, I.; Wennmalm, S.; Fyrner, T.; Ederth, T.; Öllinger, K. Interactions of the Lysosomotropic Detergent O-Methyl-Serine Dodecylamide Hydrochloride (MSDH) with Lipid Bilayer Membranes-Implications for Cell Toxicity. Int. J. Mol. Sci. 2020, 21, 3136. [Google Scholar] [CrossRef] [PubMed]
- Maleš, P.; Pem, B.; Petrov, D.; Jurašin, D.D.; Bakarić, D. Deciphering the Origin of the Melting Profile of Unilamellar Phosphatidylcholine Liposomes by Measuring the Turbidity of Its Suspensions. Soft Matter 2022, 18, 6703–6715. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Yang, H.; Pang, D.; Wei, L.; Deng, C. Effects of Mono- and Di-Valent Metal Cations on the Morphology of Lipid Vesicles. Chem. Phys. Lipids 2018, 217, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Kisak, E.T.; Kennedy, M.T.; Trommeshauser, D.; Zasadzinski, J.A. Self-Limiting Aggregation by Controlled Ligand−Receptor Stoichiometry. Langmuir 2000, 16, 2825–2831. [Google Scholar] [CrossRef]
- Chiruvolu, S.; Walker, S.; Israelachvili, J.; Schmitt, F.-J.; Leckband, D.; Zasadzinski, J.A. Higher Order Self-Assembly of Vesicles by Site-Specific Binding. Science 1994, 264, 1753–1756. [Google Scholar] [CrossRef]
- Wu, C.-M.; Chen, H.-L.; Liou, W.; Lin, T.-L.; Jeng, U.-S. DNA-Induced Aggregation of Zwitterionic Oligolamellar Liposome. Biomacromolecules 2004, 5, 2324–2328. [Google Scholar] [CrossRef]
- Maruyama, T.; Yamamura, H.; Hiraki, M.; Kemori, Y.; Takata, H.; Goto, M. Directed Aggregation and Fusion of Lipid Vesicles Induced by DNA-Surfactants. Colloids Surf. B Biointerfaces 2008, 66, 119–124. [Google Scholar] [CrossRef]
- Ryu, Y.-S.; Yoo, D.; Wittenberg, N.J.; Jordan, L.R.; Lee, S.-D.; Parikh, A.N.; Oh, S.-H. Lipid Membrane Deformation Accompanied by Disk-to-Ring Shape Transition of Cholesterol-Rich Domains. J. Am. Chem. Soc. 2015, 137, 8692–8695. [Google Scholar] [CrossRef] [PubMed]
- de Souza, T.P.; Bossa, G.V.; Stano, P.; Steiniger, F.; May, S.; Luisi, P.L.; Fahr, A. Vesicle Aggregates as a Model for Primitive Cellular Assemblies. Phys. Chem. Chem. Phys. 2017, 19, 20082–20092. [Google Scholar] [CrossRef]
- Suzuki, K.; Kurihara, K.; Okura, Y.; Toyota, T.; Sugawara, T. PH-Induced Switchable Vesicular Aggregation of Zwitterionic and Anionic Phospholipids. Chem. Lett. 2012, 41, 1084–1086. [Google Scholar] [CrossRef]
- Quemeneur, F.; Rammal, A.; Rinaudo, M.; Pépin-Donat, B. Large and Giant Vesicles “Decorated” with Chitosan: Effects of PH, Salt or Glucose Stress, and Surface Adhesion. Biomacromolecules 2007, 8, 2512–2519. [Google Scholar] [CrossRef] [PubMed]
- Makino, K.; Yamada, T.; Kimura, M.; Oka, T.; Ohshima, H.; Kondo, T. Temperature- and Ionic Strength-Induced Conformational Changes in the Lipid Head Group Region of Liposomes as Suggested by Zeta Potential Data. Biophys. Chem. 1991, 41, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Huang, J.; Gao, Y.; Fu, H. Temperature-Controlled Vesicle Aggregation in the Mixed System of Sodium n -Dodecyl Sulfate/ n -Dodecyltributylammonium Bromide. Langmuir 2005, 21, 2656–2659. [Google Scholar] [CrossRef]
- de Lange, N.; Leermakers, F.A.M.; Kleijn, J.M. Self-Limiting Aggregation of Phospholipid Vesicles. Soft Matter 2020, 16, 2379–2389. [Google Scholar] [CrossRef]
- Dhiman, N.; Sarvaiya, J.; Mohindroo, P. A Drift on Liposomes to Proliposomes: Recent Advances and Promising Approaches. J. Liposome Res. 2022, 32, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Wei, Z.; Wang, Y.; Hu, N.; Sun, S.; Bai, J.; Fang, L.; Wang, Z. Focused Characteristics and Effects of Light Reflected from Spherical Lipid Membrane of Giant Unilamellar Vesicles. Colloids Surf. B Biointerfaces 2020, 189, 110828. [Google Scholar] [CrossRef]
- Toyran, N.; Severcan, F. Infrared Spectroscopic Studies on the Dipalmitoyl Phosphatidylcholine Bilayer Interactions with Calcium Phosphate: Effect of Vitamin D2. Spectroscopy 2002, 16, 399–408. [Google Scholar] [CrossRef]
- Le Saux, A.; Ruysschaert, J.M.; Goormaghtigh, E. Membrane Molecule Reorientation in an Electric Field Recorded by Attenuated Total Reflection Fourier-Transform Infrared Spectroscopy. Biophys. J. 2001, 80, 324–330. [Google Scholar] [CrossRef]
- Terakosolphan, W.; Trick, J.L.; Royall, P.G.; Rogers, S.E.; Lamberti, O.; Lorenz, C.D.; Forbes, B.; Harvey, R.D. Glycerol Solvates DPPC Headgroups and Localizes in the Interfacial Regions of Model Pulmonary Interfaces Altering Bilayer Structure. Langmuir 2018, 34, 6941–6954. [Google Scholar] [CrossRef]
- Bonforti, A.; Solé, R. Unicellular-Multicellular Evolutionary Branching Driven by Resource Limitations. J. R. Soc. Interface 2022, 19, 20220018. [Google Scholar] [CrossRef]
- Penkov, N.V.; Yashin, V.A.; Belosludtsev, K.N. Hydration Shells of DPPC Liposomes from the Point of View of Terahertz Time-Domain Spectroscopy. Appl. Spectrosc. 2021, 75, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Fenz, S.F.; Sengupta, K. Giant Vesicles as Cell Models. Integr. Biol. 2012, 4, 982. [Google Scholar] [CrossRef]
- Boban, Z.; Puljas, A.; Kovač, D.; Subczynski, W.K.; Raguz, M. Effect of Electrical Parameters and Cholesterol Concentration on Giant Unilamellar Vesicles Electroformation. Cell Biochem. Biophys. 2020, 78, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yan, D.; Dong, W.; Tian, Y. Temperature-Responsive Phase Transition of Polymer Vesicles: Real-Time Morphology Observation and Molecular Mechanism. J. Phys. Chem. B 2007, 111, 1262–1270. [Google Scholar] [CrossRef]
- Yin, H.; Zhou, Z.; Huang, J.; Zheng, R.; Zhang, Y. Temperature-Induced Micelle to Vesicle Transition in the Sodium Dodecylsulfate/Dodecyltriethylammonium Bromide System. Angew. Chem. Int. Ed. 2003, 42, 2188–2191. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, K.; Nakanishi, H.; Sakai, H.; Abe, M. Temperature-Dependent Vesicle Formation of Aqueous Solutions of Mixed Cationic and Anionic Surfactants. Langmuir 2004, 20, 2117–2122. [Google Scholar] [CrossRef]
- Schenkman, S.; Araujo, P.S.; Dukman, R.; Quina, F.H.; Chaimovich, H. Effects of Temperature and Lipid Composition on the Serum Albumin-Induced Aggregation and Fusion of Small Unilamellar Vesicles. Biochim. Biophys. Acta—Biomembr. 1981, 649, 633–641. [Google Scholar] [CrossRef]
- Andes-Koback, M.; Keating, C.D. Complete Budding and Asymmetric Division of Primitive Model Cells To Produce Daughter Vesicles with Different Interior and Membrane Compositions. J. Am. Chem. Soc. 2011, 133, 9545–9555. [Google Scholar] [CrossRef]
- Carrara, P.; Stano, P.; Luisi, P.L. Giant Vesicles “Colonies”: A Model for Primitive Cell Communities. ChemBioChem 2012, 13, 1497–1502. [Google Scholar] [CrossRef]
- Herzog, M.; Dwivedi, M.; Kumar Harishchandra, R.; Bilstein, A.; Galla, H.-J.; Winter, R. Effect of Ectoine, Hydroxyectoine and β-Hydroxybutyrate on the Temperature and Pressure Stability of Phospholipid Bilayer Membranes of Different Complexity. Colloids Surf. B Biointerfaces 2019, 178, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Huang, R.; Corti, D.S.; Franses, E.I. Colloidal Dispersion Stability of Unilamellar DPPC Vesicles in Aqueous Electrolyte Solutions and Comparisons to Predictions of the DLVO Theory. J. Colloid Interface Sci. 2010, 342, 300–310. [Google Scholar] [CrossRef]
- Inoue, T.; Minami, H.; Shimozawa, R.; Sugihara, G. Stability of DLPA/DLPC Mixed Vesicles against Divalent Cation-Induced Aggregation: Importance of the Hydration Force. J. Colloid Interface Sci. 1992, 152, 493–506. [Google Scholar] [CrossRef]
- Yin, H.; Lin, Y.; Huang, J.; Ye, J. Temperature-Induced Vesicle Aggregation in Catanionic Surfactant Systems: The Effects of the Headgroup and Counterion. Langmuir 2007, 23, 4225–4230. [Google Scholar] [CrossRef]
- Carvalho, K.; Ramos, L.; Roy, C.; Picart, C. Giant Unilamellar Vesicles Containing Phosphatidylinositol(4,5)Bisphosphate: Characterization and Functionality. Biophys. J. 2008, 95, 4348–4360. [Google Scholar] [CrossRef] [PubMed]
- Salipante, P.F.; Vlahovska, P.M. Vesicle Deformation in DC Electric Pulses. Soft Matter 2014, 10, 3386–3393. [Google Scholar] [CrossRef]
- Dimova, R.; Riske, K.A.; Aranda, S.; Bezlyepkina, N.; Knorr, R.L.; Lipowsky, R. Giant Vesicles in Electric Fields. Soft Matter 2007, 3, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Chibowski, E.; Szcześ, A. Zeta Potential and Surface Charge of DPPC and DOPC Liposomes in the Presence of PLC Enzyme. Adsorption 2016, 22, 755–765. [Google Scholar] [CrossRef]
- Sęk, A.; Perczyk, P.; Wydro, P.; Gruszecki, W.I.; Szcześ, A. Effect of Trace Amounts of Ionic Surfactants on the Zeta Potential of DPPC Liposomes. Chem. Phys. Lipids 2021, 235, 105059. [Google Scholar] [CrossRef]
- Morini, M.A.; Sierra, M.B.; Pedroni, V.I.; Alarcon, L.M.; Appignanesi, G.A.; Disalvo, E.A. Influence of Temperature, Anions and Size Distribution on the Zeta Potential of DMPC, DPPC and DMPE Lipid Vesicles. Colloids Surf. B Biointerfaces 2015, 131, 54–58. [Google Scholar] [CrossRef]
- Verde, A.R.; Sierra, M.B.; Alarcón, L.M.; Pedroni, V.I.; Appignanesi, G.A.; Morini, M.A. Experimental and Computational Studies of the Effects of Free DHA on a Model Phosphatidylcholine Membrane. Chem. Phys. Lipids 2018, 217, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Umemura, J.; Cameron, D.G.; Mantsch, H.H. A Fourier Transform Infrared Spectroscopic Study of the Molecular Interaction of Cholesterol with 1,2-Dipalmitoyl-Sn-Glycero-3-Phosphocholine. Biochim. Et Biophys. Acta—Biomembr. 1980, 602, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Doux, J.P.F.; Hall, B.A.; Killian, J.A. How Lipid Headgroups Sense the Membrane Environment: An Application of 14N NMR. Biophys. J. 2012, 103, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Monnard, P.-A.; Apel, C.L.; Kanavarioti, A.; Deamer, D.W. Influence of Ionic Inorganic Solutes on Self-Assembly and Polymerization Processes Related to Early Forms of Life: Implications for a Prebiotic Aqueous Medium. Astrobiology 2002, 2, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Wiącek, A.E. Comparison of N-Tetradecane/Electrolyte Emulsions Properties Stabilized by DPPC and DPPC Vesicles in the Electrolyte Solution. Colloids Surf. B Biointerfaces 2011, 83, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Knecht, V.; Klasczyk, B. Specific Binding of Chloride Ions to Lipid Vesicles and Implications at Molecular Scale. Biophys. J. 2013, 104, 818–824. [Google Scholar] [CrossRef]
- Zheng, L.-Q.; Shui, L.; Shen, Q.; Li, G.-Z.; Baba, T.; Minamikawa, H.; Hato, M. PH and Salt-Induced Reversible Aggregation of Nonionic Synthetic Glycolipid Vesicles. Colloids Surf. A Physicochem. Eng. Asp. 2002, 207, 215–221. [Google Scholar] [CrossRef]
- Rodriguez, J.R.; García, A.E. Concentration Dependence of NaCl Ion Distributions around DPPC Lipid Bilayers. Interdiscip. Sci. Comput. Life Sci. 2011, 3, 272–282. [Google Scholar] [CrossRef]
- Smistad, G.; Bøyum, S.; Alund, S.J.; Samuelsen, A.B.C.; Hiorth, M. The Potential of Pectin as a Stabilizer for Liposomal Drug Delivery Systems. Carbohydr. Polym. 2012, 90, 1337–1344. [Google Scholar] [CrossRef]
- Jiménez-Rojo, N.; Viguera, A.R.; Collado, M.I.; Sims, K.H.; Constance, C.; Hill, K.; Shaw, W.A.; Goñi, F.M.; Alonso, A. Sphingosine Induces the Aggregation of Imine-Containing Peroxidized Vesicles. Biochim. Biophys. Acta—Biomembr. 2014, 1838, 2071–2077. [Google Scholar] [CrossRef]
- Grit, M.; Crommelin, D.J.A. Chemical Stability of Liposomes: Implications for Their Physical Stability. Chem. Phys. Lipids 1993, 64, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Scotese, C.R. An Atlas of Phanerozoic Paleogeographic Maps: The Seas Come In and the Seas Go Out. Annu. Rev. Earth Planet. Sci. 2021, 49, 679–728. [Google Scholar] [CrossRef]
- Collins, P. Size and Cycle: An Essay on the Structure of Biology; Princeton University Press: Princeton, NJ, USA, 1965. [Google Scholar]
- Bonner, J.T. Perspective: The Size-Complexity Rule. Evolution 2004, 58, 1883–1890. [Google Scholar] [CrossRef] [PubMed]
- Bonner, J.T. The Origins of Multicellularity. Integr. Biol. 1998, 1, 27–36. [Google Scholar] [CrossRef]
- Hall, B.K. A Review of First Signals: The Evolution of Multicellular Development. Evol. Dev. 2001, 3, 223–224. [Google Scholar] [CrossRef]
- Queller, D.C. Relatedness and the Fraternal Major Transitions. Philos. Trans. R. Soc. Lond. B 2000, 355, 1647–1655. [Google Scholar] [CrossRef]
- Dworkin, M.; Bonner, J.T. The Myxobacteria: New Directions in Studies of Procaryotic Development. CRC Crit. Rev. Microbiol. 1972, 1, 435–452. [Google Scholar] [CrossRef]
- Shimkets, L.J. Social and Developmental Biology of the Myxobacteria. Microbiol. Rev. 1990, 54, 473–501. [Google Scholar] [CrossRef]
- Blanton, R.L.; Olive, L.S. Ultrastructure of Aerial Stalk Formation by the Ciliated ProtozoanSorogena Stoianovitchae. Protoplasma 1983, 116, 125–135. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature | N(CH3)3+ Stretching (cm−1) |
|---|---|
| 25 °C | 971.49 ± 0.37 ** |
| 45 °C | 969.84 ± 0.21 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhang, Y.; Xie, M.; Wang, Z.; Qiao, H. Temperature-Promoted Giant Unilamellar Vesicle (GUV) Aggregation: A Way of Multicellular Formation. Curr. Issues Mol. Biol. 2023, 45, 3757-3771. https://doi.org/10.3390/cimb45050242
Wang X, Zhang Y, Xie M, Wang Z, Qiao H. Temperature-Promoted Giant Unilamellar Vesicle (GUV) Aggregation: A Way of Multicellular Formation. Current Issues in Molecular Biology. 2023; 45(5):3757-3771. https://doi.org/10.3390/cimb45050242
Chicago/Turabian StyleWang, Xinmao, Yangruizi Zhang, Maobin Xie, Zhibiao Wang, and Hai Qiao. 2023. "Temperature-Promoted Giant Unilamellar Vesicle (GUV) Aggregation: A Way of Multicellular Formation" Current Issues in Molecular Biology 45, no. 5: 3757-3771. https://doi.org/10.3390/cimb45050242
APA StyleWang, X., Zhang, Y., Xie, M., Wang, Z., & Qiao, H. (2023). Temperature-Promoted Giant Unilamellar Vesicle (GUV) Aggregation: A Way of Multicellular Formation. Current Issues in Molecular Biology, 45(5), 3757-3771. https://doi.org/10.3390/cimb45050242