
A Novel Approach to Reducing Lung Metastasis in Osteosarcoma: Increasing Cell Stiffness with Carbenoxolone

 , , ,
, , ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Fluorescence Staining and Analysis of F-Actin
2.3. Measurement of Cell Stiffness
Evaluation of Cell Processes In Vitro
2.4. Proliferation Assay
2.5. Migration Assay
2.6. Invasion Assay
2.7. Adhesion Assay
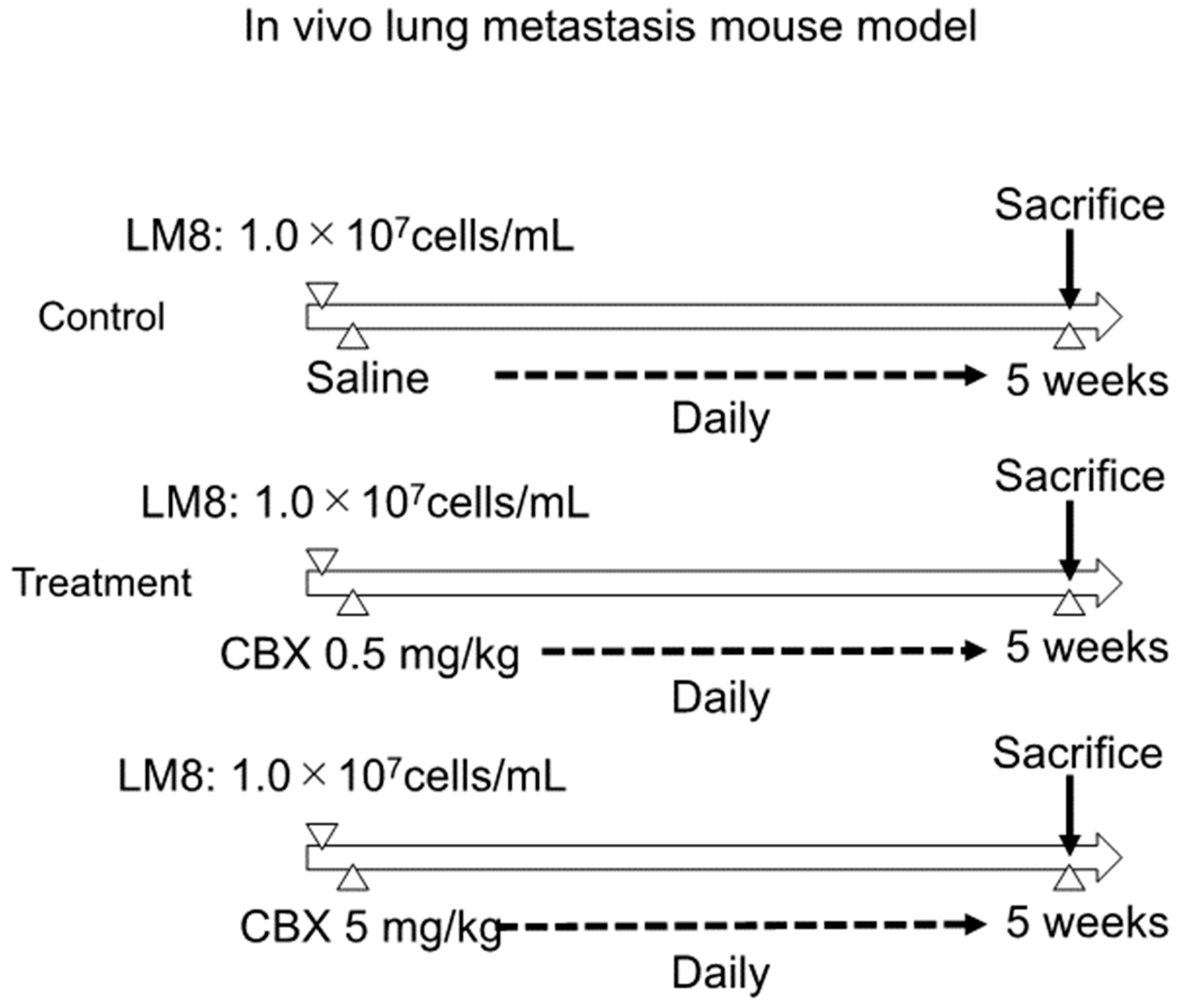
2.8. In Vivo Lung Metastasis Mouse Model
2.9. Statistical Analyses
3. Results
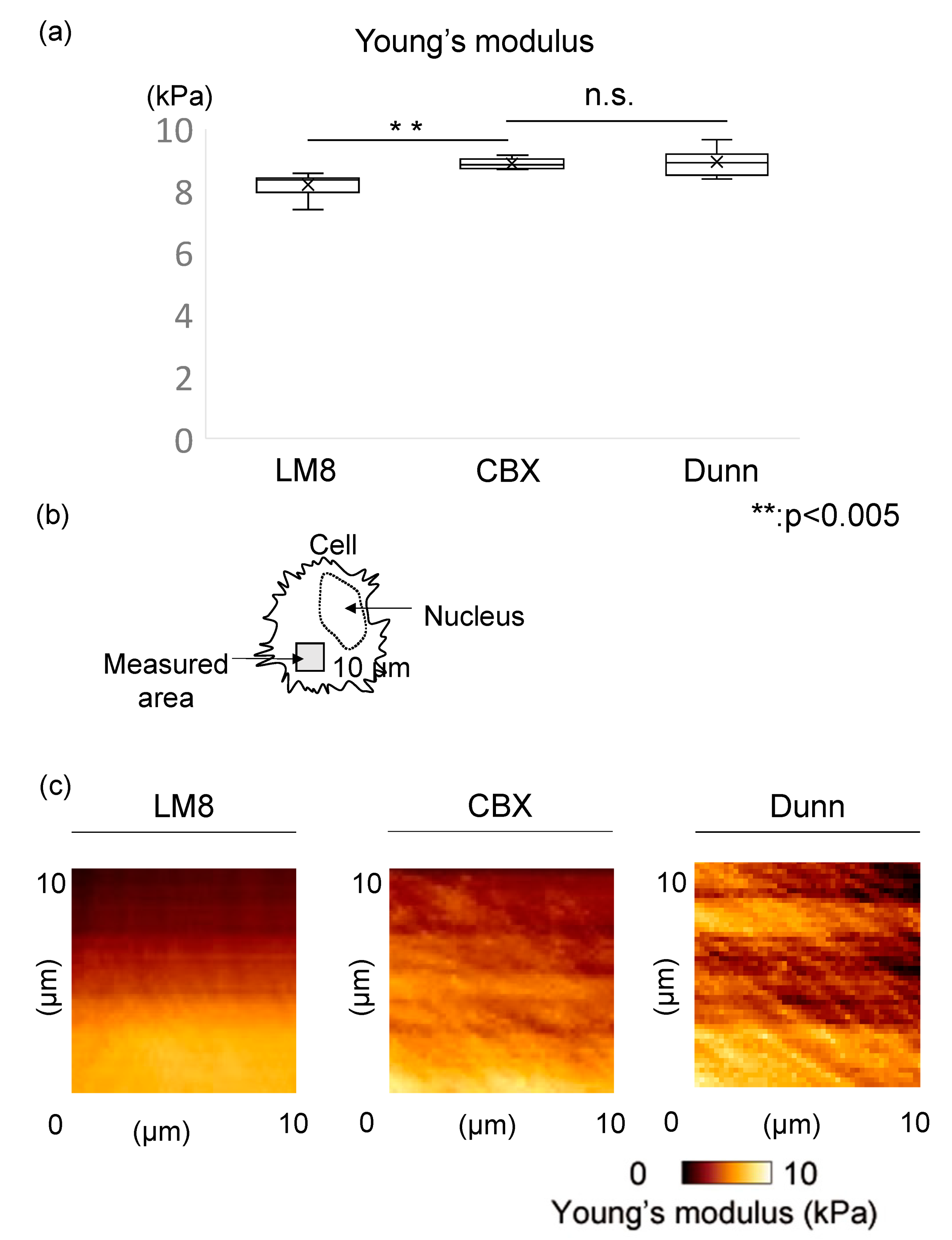
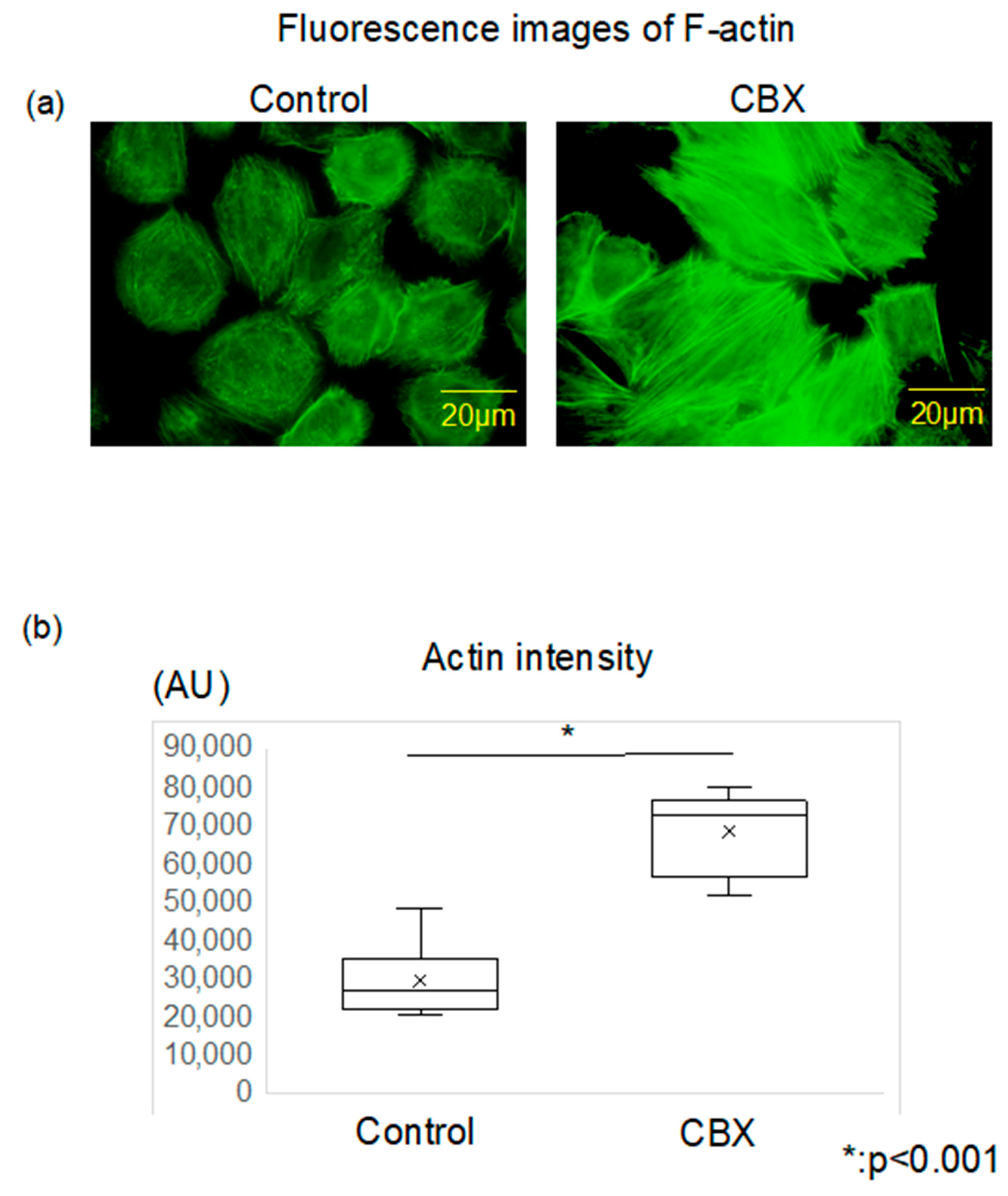
3.1. CBX Increases Cell Stiffness and Actin Polymerization
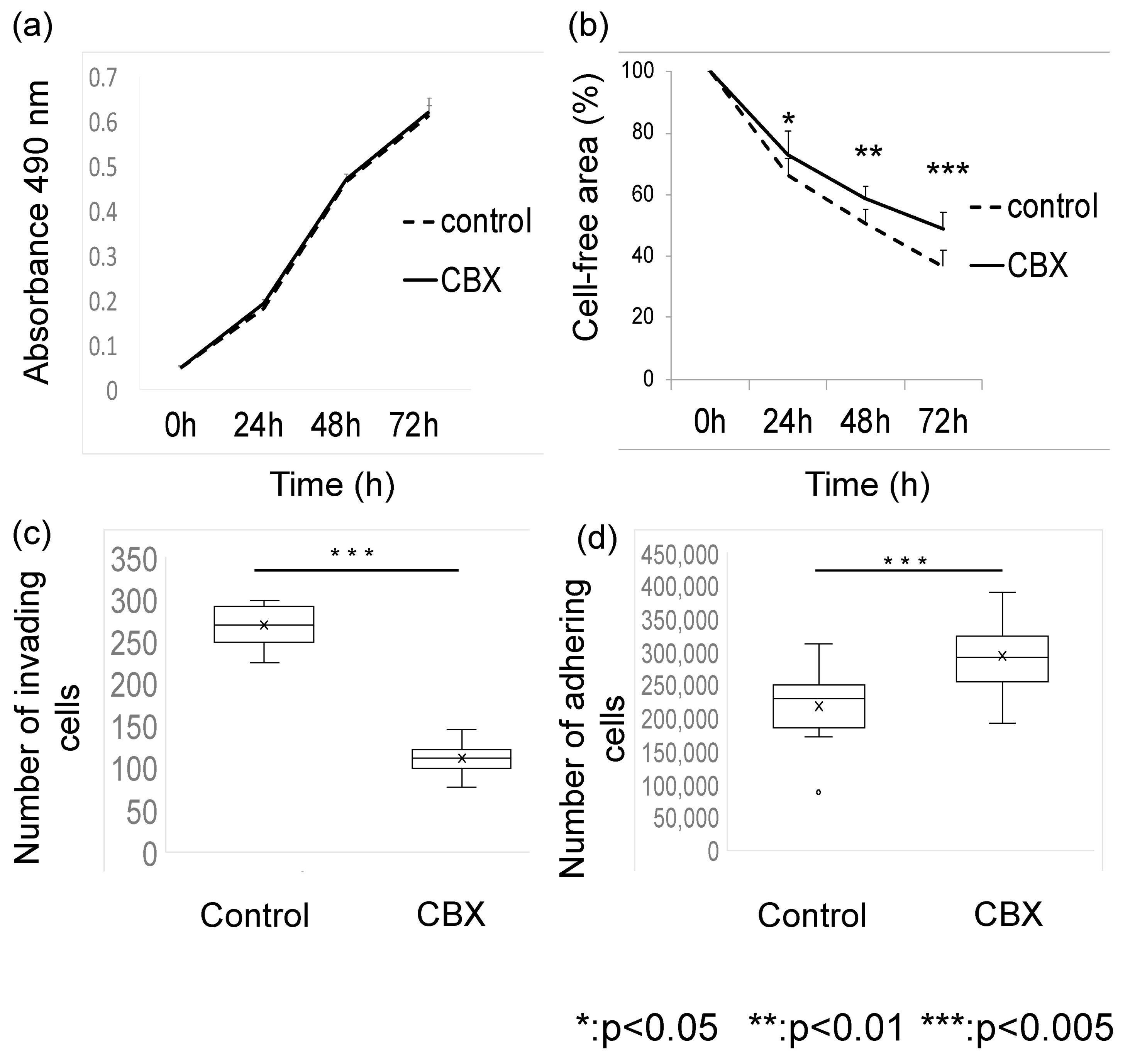
3.2. CBX Affects Metastatic Potential
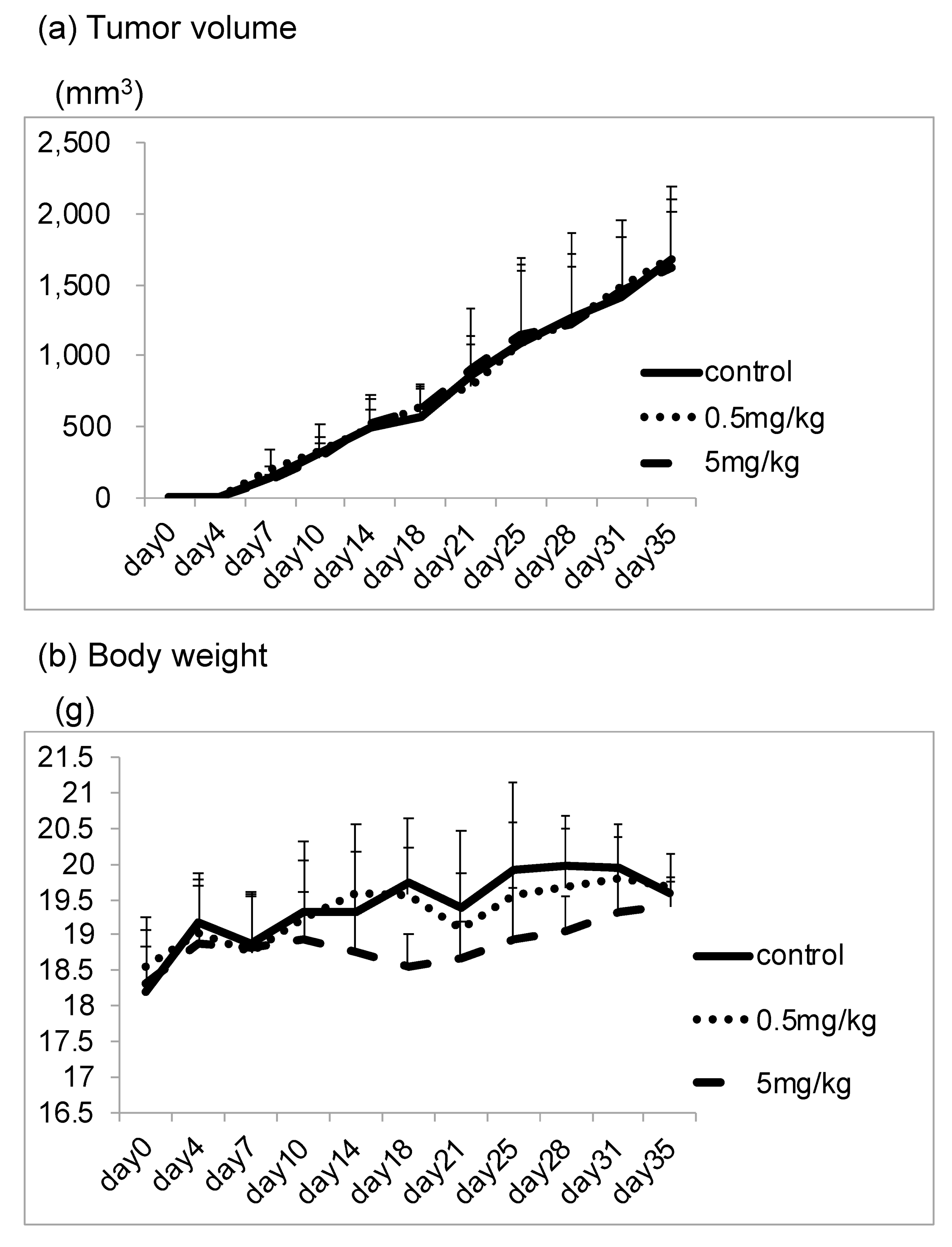
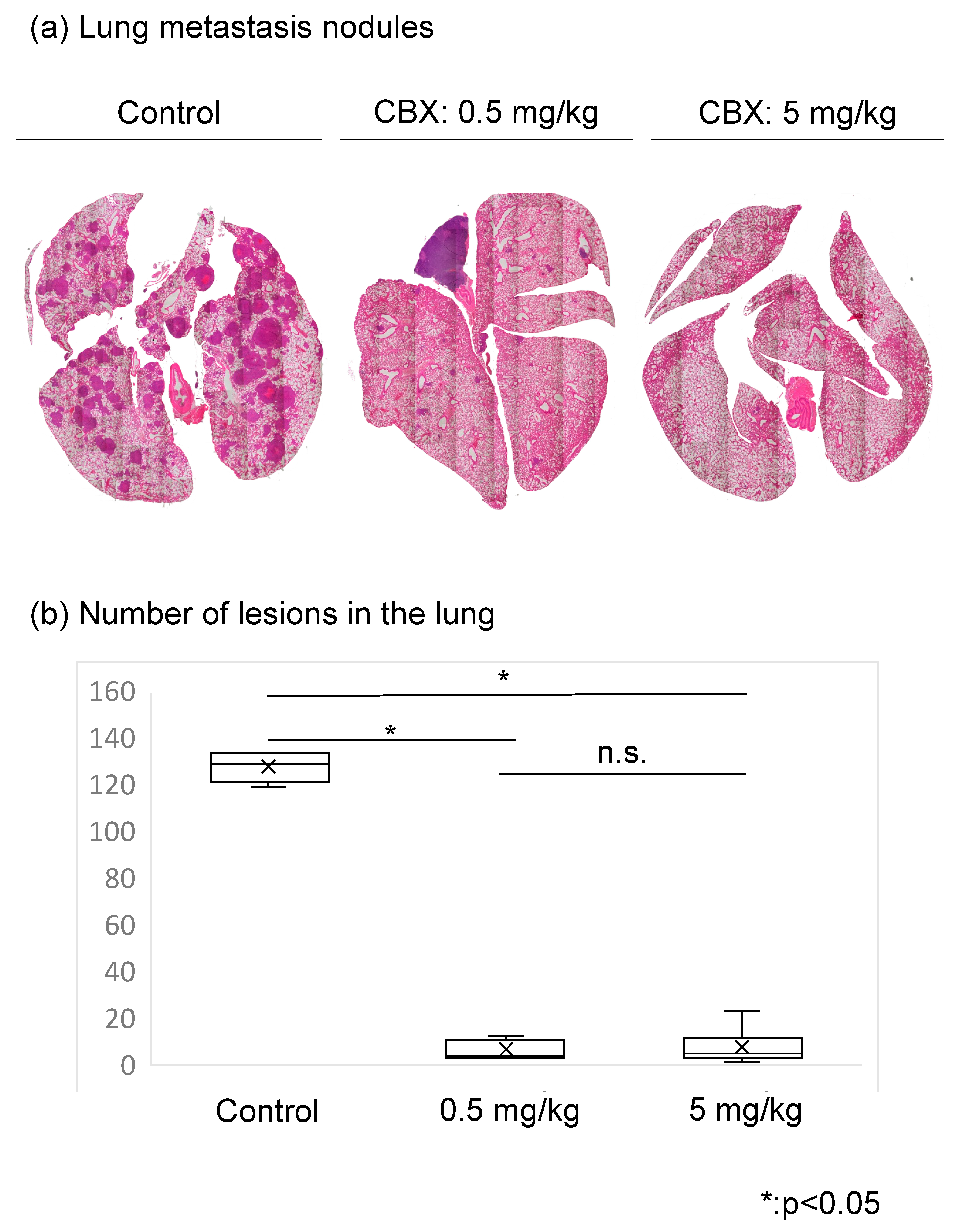
3.3. CBX Reduces the Number of Lesions in the Lung
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaya, M.; Wada, T.; Kawaguchi, S.; Nagoya, S.; Yamashita, T.; Abe, Y.; Hiraga, H.; Isu, K.; Shindoh, M.; Higashino, F.; et al. Increased pre-therapeutic serum vascular endothelial growth factor in patients with early clinical relapse of osteosarcoma. Br. J. Cancer 2002, 86, 864–869. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, D.; Konstantopoulos, K.; Searson, P.C. The physics of cancer: The role of physical interactions and mechanical forces in metastasis. Nat. Rev. Cancer 2011, 11, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qin, G.; Liang, X.; Wang, W.; Wang, Z.; Liao, D.; Zhong, L.; Zhang, R.; Zeng, Y.X.; Wu, Y.; et al. Targeting the CK1α/CBX4 axis for metastasis in osteosarcoma. Nat. Commun. 2020, 11, 1141. [Google Scholar] [CrossRef] [PubMed]
- Yui, Y.; Kumai, J.; Watanabe, K.; Wakamatsu, T.; Sasagawa, S. Lung fibrosis is novel therapeutic target to suppress lung metastasis of osteosarcoma. Int. J. Cancer 2022, 151, 739–751. [Google Scholar] [CrossRef]
- Bendas, G.; Borsig, L. Heparanase in Cancer Metastasis—Heparin as a Potential Inhibitor of Cell Adhesion Molecules. Adv. Exp. Med. Biol. 2020, 1221, 309–329. [Google Scholar]
- Holenstein, C.N.; Horvath, A.; Schar, B.; Schoenenberger, A.D.; Bollhalder, M.; Goedecke, N.; Bartalena, G.; Otto, O.; Herbig, M.; Guck, J.; et al. The relationship between metastatic potential and in vitro mechanical properties of osteosarcoma cells. Mol. Biol. Cell 2019, 30, 887–898. [Google Scholar] [CrossRef]
- Xu, W.; Mezencev, R.; Kim, B.; Wang, L.; McDonald, J.F.; Sulchek, T. Cell Stiffness Is a Biomarker of the Metastatic Potential of Ovarian Cancer Cells. PLoS ONE 2012, 7, e46609. [Google Scholar] [CrossRef]
- Kita, K.; Asanuma, K.; Okamoto, T.; Kawamoto, E.; Nakamura, K.; Hagi, T.; Nakamura, T.; Shimaoka, M.; Sudo, A. Cytoskeletal Actin Structure in Osteosarcoma Cells Determines Metastatic Phenotype via Regulating Cell Stiffness, Migration, and Transmigration. Curr. Issues Mol. Biol. 2021, 43, 1255–1266. [Google Scholar] [CrossRef]
- Okamoto, T.; Kawamoto, E.; Takagi, Y.; Akita, N.; Hayashi, T.; Park, E.J.; Suzuki, K.; Shimaoka, M. Gap junction-mediated regulation of endothelial cellular stiffness. Sci. Rep. 2017, 7, 6134. [Google Scholar] [CrossRef]
- Qiu, X.; Cheng, J.C.; Klausen, C.; Chang, H.M.; Fan, Q.; Leung, P.C. EGF-Induced Connexin43 Negatively Regulates Cell Proliferation in Human Ovarian Cancer. J. Cell Physiol. 2016, 231, 111–119. [Google Scholar] [CrossRef]
- Li, W.; Li, J.; Sama, A.E.; Wang, H. Carbenoxolone blocks endotoxin-induced protein kinase R (PKR) activation and high mobility group box 1 (HMGB1) release. Mol. Med. 2013, 19, 203–211. [Google Scholar] [CrossRef]
- Poste, G.; Fidler, I.J. The pathogenesis of cancer metastasis. Nature 1980, 283, 139–146. [Google Scholar] [CrossRef]
- Asai, T.; Ueda, T.; Itoh, K.; Yoshioka, K.; Aoki, Y.; Mori, S.; Yoshikawa, H. Establishment and characterization of a murine osteosarcoma cell line (LM8) with high metastatic potential to the lung. Int. J. Cancer 1998, 76, 418–422. [Google Scholar] [CrossRef]
- Luo, Q.; Kuang, D.; Zhang, B.; Song, G. Cell stiffness determined by atomic force microscopy and its correlation with cell motility. Biochim. Biophys. Acta 2016, 1860, 1953–1960. [Google Scholar] [CrossRef]
- Kawamoto, E.; Nago, N.; Okamoto, T.; Gaowa, A.; Masui-Ito, A.; Sakakura, Y.; Akama, Y.; Soe, Z.Y.; Prajuabjinda, O.; Darkwah, S.; et al. Anti-adhesive effects of human soluble thrombomodulin and its domains. Biochem. Biophys. Res. Commun. 2019, 511, 312–317. [Google Scholar] [CrossRef]
- Atsumi, S.; Matsumine, A.; Toyoda, H.; Niimi, R.; Iino, T.; Nakamura, T.; Matsubara, T.; Asanuma, K.; Komada, Y.; Uchida, A.; et al. Oncolytic virotherapy for human bone and soft tissue sarcomas using live attenuated poliovirus. Int. J. Oncol. 2012, 41, 893–902. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transpl. 2013, 48, 452–458. [Google Scholar] [CrossRef]
- Tian, H.; Guan, D.; Li, J. Identifying osteosarcoma metastasis associated genes by weighted gene coexpression network analysis (WGCNA). Medicine 2018, 97, e10781. [Google Scholar] [CrossRef]
- Anderson, R.L.; Balasas, T.; Callaghan, J.; Coombes, R.C.; Evans, J.; Hall, J.A.; Kinrade, S.; Jones, D.; Jones, P.S.; Jones, R.; et al. A framework for the development of effective anti-metastatic agents. Nat. Rev. Clin. Oncol. 2019, 16, 185–204. [Google Scholar] [CrossRef]
- Fidler, I.J.; Kripke, M.L. The challenge of targeting metastasis. Cancer Metastasis Rev. 2015, 34, 635–641. [Google Scholar] [CrossRef]
- Schaefer, A.; Hordijk, P.L. Cell-stiffness-induced mechanosignaling—A key driver of leukocyte transendothelial migration. J. Cell Sci. 2015, 128, 2221–2230. [Google Scholar] [CrossRef] [PubMed]
- Belardi, B.; Hamkins-Indik, T.; Harris, A.R.; Kim, J.; Xu, K.; Fletcher, D.A. A Weak Link with Actin Organizes Tight Junctions to Control Epithelial Permeability. Dev. Cell 2020, 54, 792–804. [Google Scholar] [CrossRef] [PubMed]
- Cartagena-Rivera, A.X.; Van Itallie, C.M.; Anderson, J.M.; Chadwick, R.S. Apical surface supracellular mechanical properties in polarized epithelium using noninvasive acoustic force spectroscopy. Nat. Commun. 2017, 8, 1030. [Google Scholar] [CrossRef] [PubMed]
- Mierke, C.T.; Kollmannsberger, P.; Zitterbart, D.P.; Smith, J.; Fabry, B.; Goldmann, W.H. Mechano-coupling and regulation of contractility by the vinculin tail domain. Biophys. J. 2008, 94, 661–670. [Google Scholar] [CrossRef]
- Batra, N.; Burra, S.; Siller-Jackson, A.J.; Gu, S.; Xia, X.; Weber, G.F.; DeSimone, D.; Bonewald, L.F.; Lafer, E.M.; Sprague, E.; et al. Mechanical stress-activated integrin α5β1 induces opening of connexin 43 hemichannels. Proc. Natl. Acad. Sci. USA 2012, 109, 3359–3364. [Google Scholar] [CrossRef]
- Shimaoka, M.; Kawamoto, E.; Gaowa, A.; Okamoto, T.; Park, E.J. Connexins and Integrins in Exosomes. Cancers 2019, 11, 106. [Google Scholar] [CrossRef]
- Wu, X.; Muthuchamy, M.; Reddy, D.S. Atomic force microscopy investigations of fibronectin and alpha5beta1-integrin signaling in neuroplasticity and seizure susceptibility in experimental epilepsy. Epilepsy Res. 2017, 138, 71–80. [Google Scholar] [CrossRef]
- Olk, S.; Turchinovich, A.; Grzendowski, M.; Stuhler, K.; Meyer, H.E.; Zoidl, G.; Dermietzel, R. Proteomic analysis of astroglial connexin43 silencing uncovers a cytoskeletal platform involved in process formation and migration. Glia 2010, 58, 494–505. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, M.; Huang, J.; Li, Y.; Wang, S.; Harrington, C.A.; Qian, D.Z.; Sun, X.X.; Dai, M.S. microRNA-130a suppresses breast cancer cell migration and invasion by targeting FOSL1 and upregulating ZO-1. J. Cell Biochem. 2018, 119, 4945–4956. [Google Scholar] [CrossRef]
- Armanini, D.; Calo, L.; Semplicini, A. Pseudohyperaldosteronism: Pathogenetic mechanisms. Crit. Rev. Clin. Lab. Sci. 2003, 40, 295–335. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kita, K.; Asanuma, K.; Okamoto, T.; Kawamoto, E.; Nakamura, K.; Hagi, T.; Nakamura, T.; Shimaoka, M.; Sudo, A. A Novel Approach to Reducing Lung Metastasis in Osteosarcoma: Increasing Cell Stiffness with Carbenoxolone. Curr. Issues Mol. Biol. 2023, 45, 4375-4388. https://doi.org/10.3390/cimb45050278
Kita K, Asanuma K, Okamoto T, Kawamoto E, Nakamura K, Hagi T, Nakamura T, Shimaoka M, Sudo A. A Novel Approach to Reducing Lung Metastasis in Osteosarcoma: Increasing Cell Stiffness with Carbenoxolone. Current Issues in Molecular Biology. 2023; 45(5):4375-4388. https://doi.org/10.3390/cimb45050278
Chicago/Turabian StyleKita, Kouji, Kunihiro Asanuma, Takayuki Okamoto, Eiji Kawamoto, Koichi Nakamura, Tomohito Hagi, Tomoki Nakamura, Motomu Shimaoka, and Akihiro Sudo. 2023. "A Novel Approach to Reducing Lung Metastasis in Osteosarcoma: Increasing Cell Stiffness with Carbenoxolone" Current Issues in Molecular Biology 45, no. 5: 4375-4388. https://doi.org/10.3390/cimb45050278
APA StyleKita, K., Asanuma, K., Okamoto, T., Kawamoto, E., Nakamura, K., Hagi, T., Nakamura, T., Shimaoka, M., & Sudo, A. (2023). A Novel Approach to Reducing Lung Metastasis in Osteosarcoma: Increasing Cell Stiffness with Carbenoxolone. Current Issues in Molecular Biology, 45(5), 4375-4388. https://doi.org/10.3390/cimb45050278