
Magnolia kobus Extract Suppresses Porphyromonas gingivalis LPS-Induced Proinflammatory Cytokine and MMP Expression in HGF-1 Cells and Regulates Osteoclastogenesis in RANKL-Stimulated RAW264.7 Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Magnolia kobus Extract (MKE)
2.2. Cell Culture
2.3. Cell Viability Measurement
2.4. TRAP Staining and Activity Analysis
2.5. RNA Extraction and qPCR
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
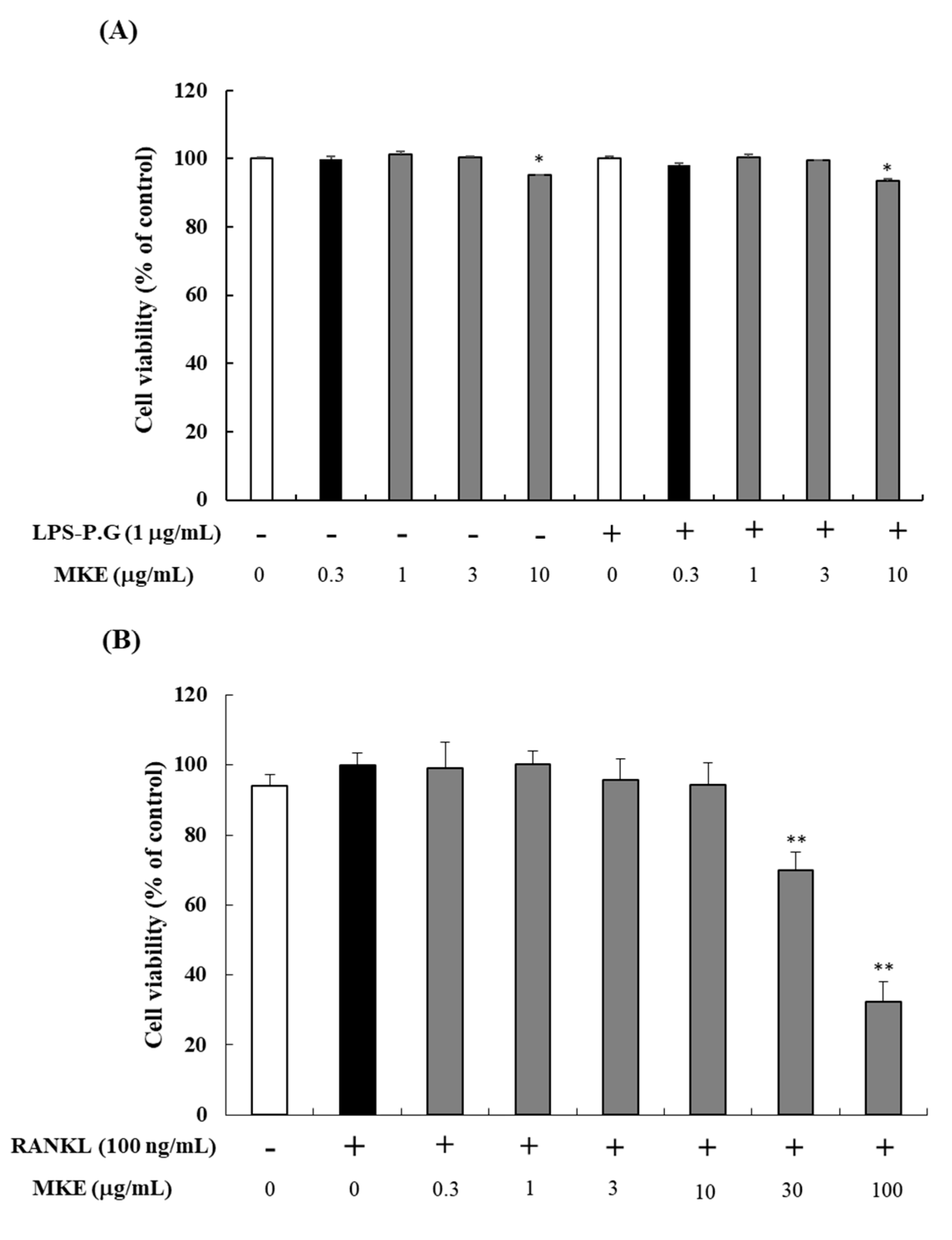
3.1. Effects of MKE on Cell Viability in HGF-1 Cells and RANKL-Induced Osteoclasts
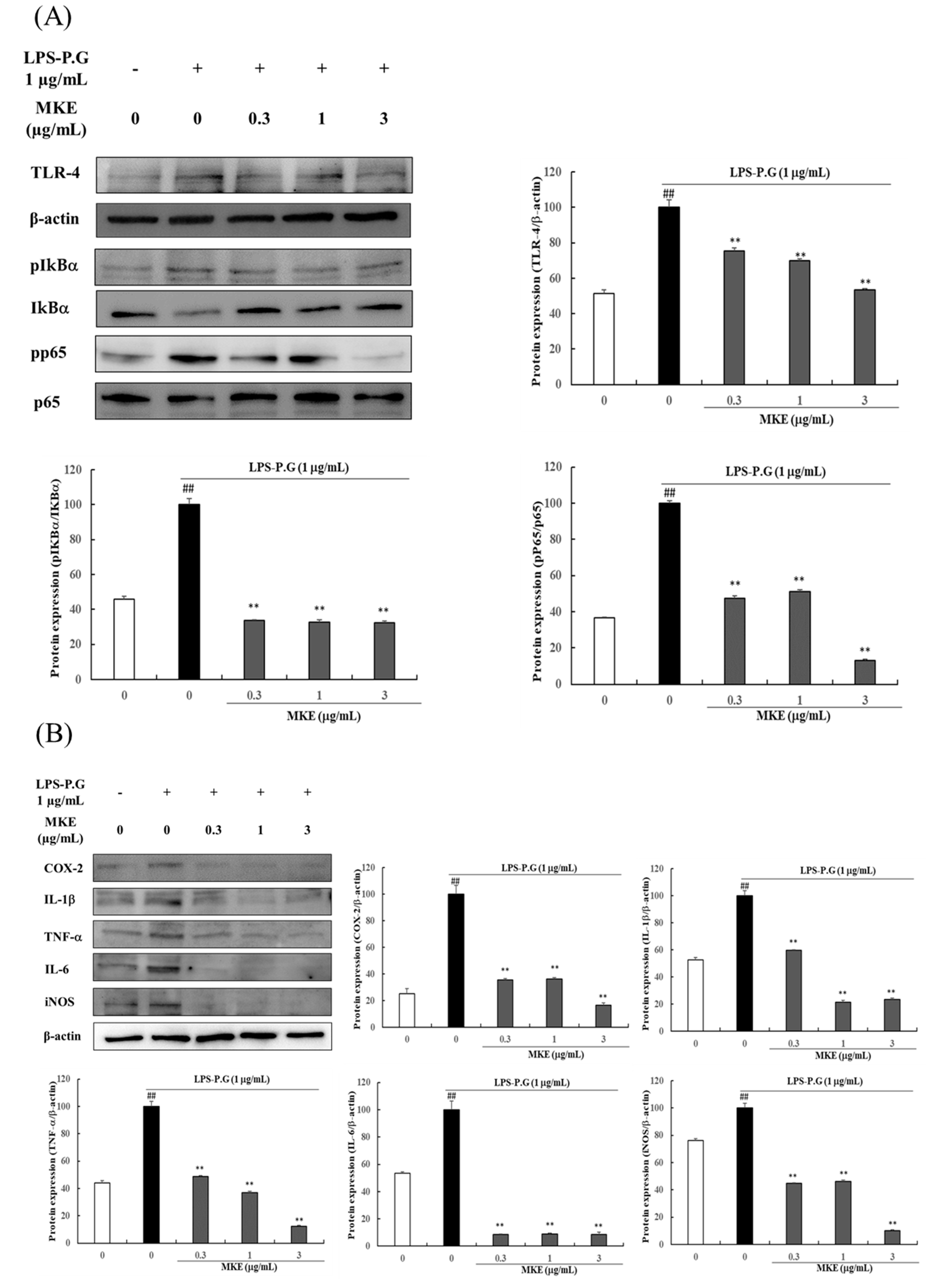
3.2. MKE Reduces Gingival Inflammation by Regulating the TLR4/NF-κB Pathway in LPS-PG-Treated HGF-1 Cells
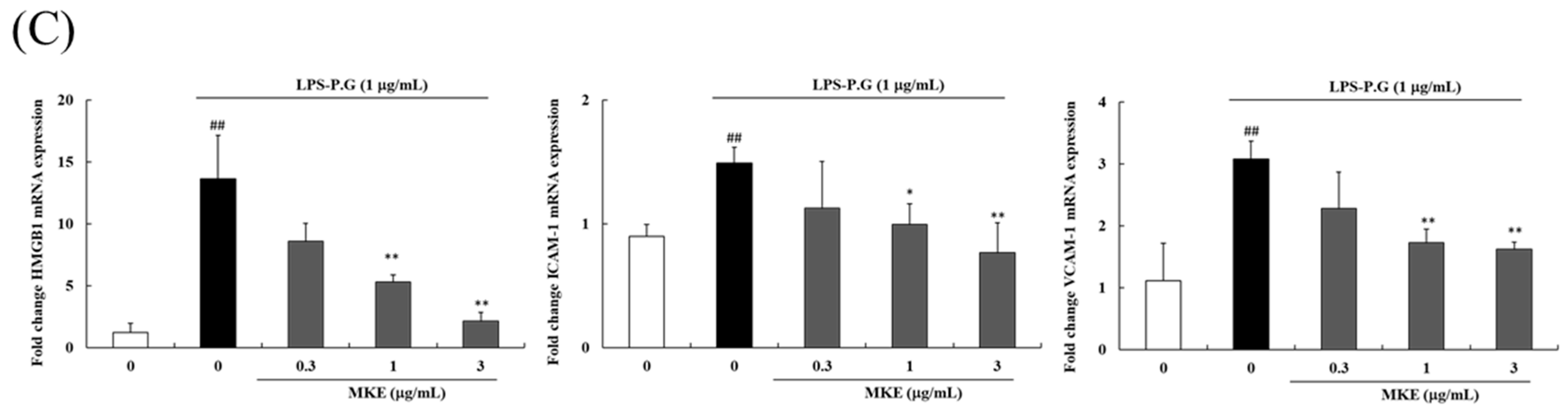
3.3. MKE Inhibits LPS-PG-Stimulated Gingival Inflammation and Decreases HMGB1, ICAM-1, and VCAM-1 Expression in HGF-1 Cells
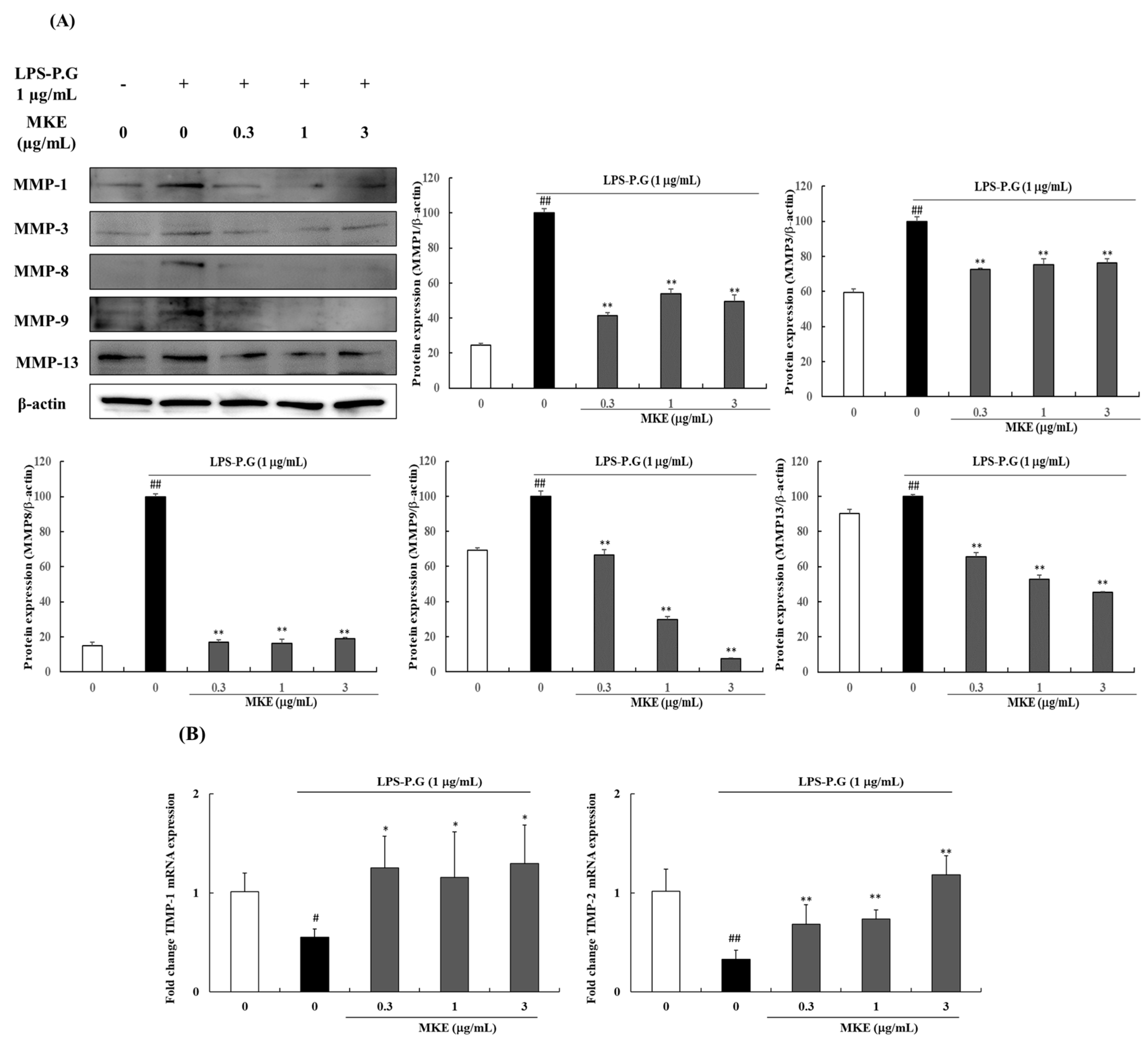
3.4. MKE Downregulates MMP Expression and Upregulates TIMP Expression in LPS-PG-Stimulated HGF-1 Cells
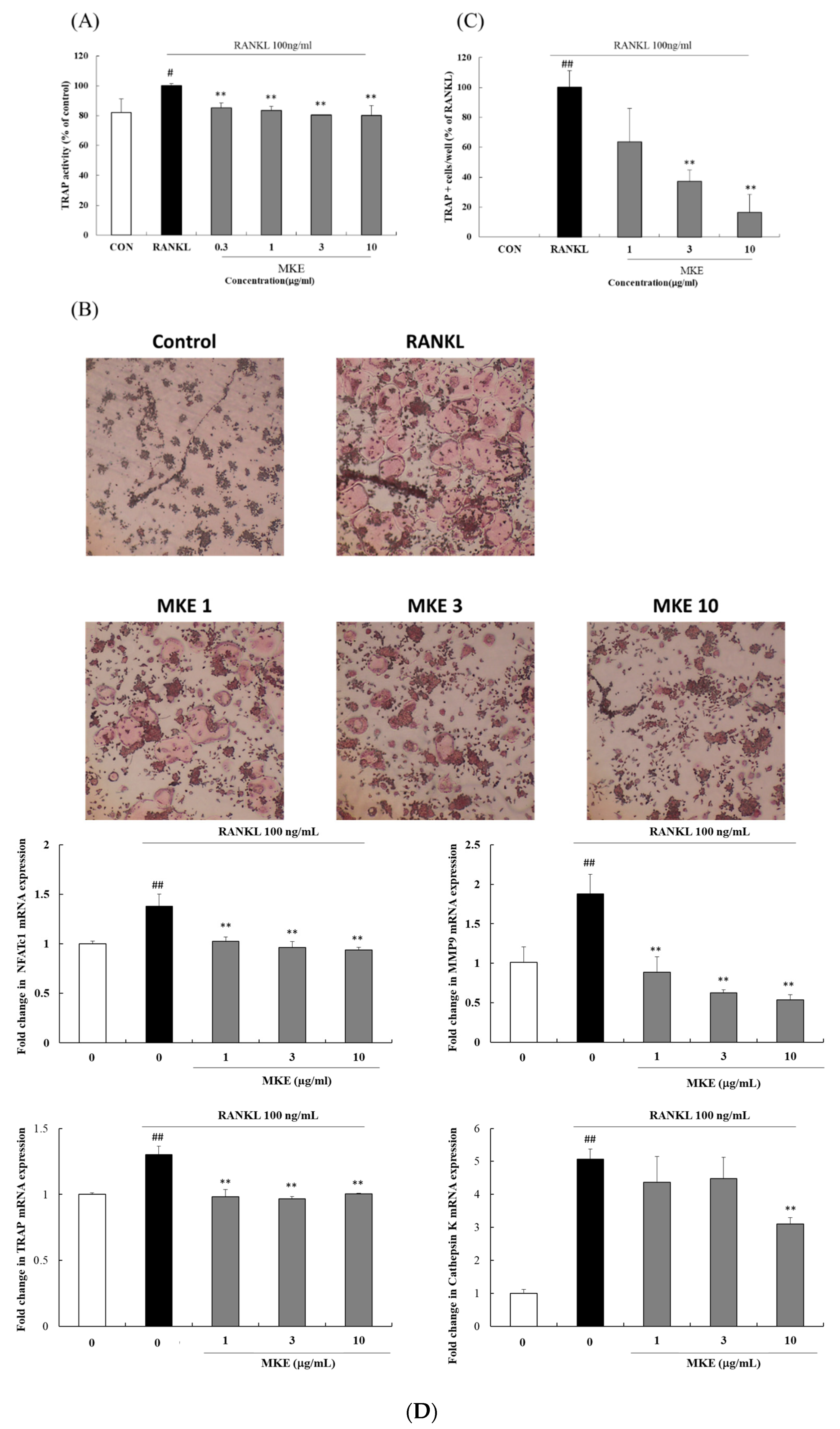
3.5. MKE Inhibits Osteoclastogenesis in RAW264.7 Cells Induced by RANKL
3.6. MKE Downregulates Osteoclast Differentiation-Related Gene Expression in RANKL-Stimulated RAW264.7 Cells
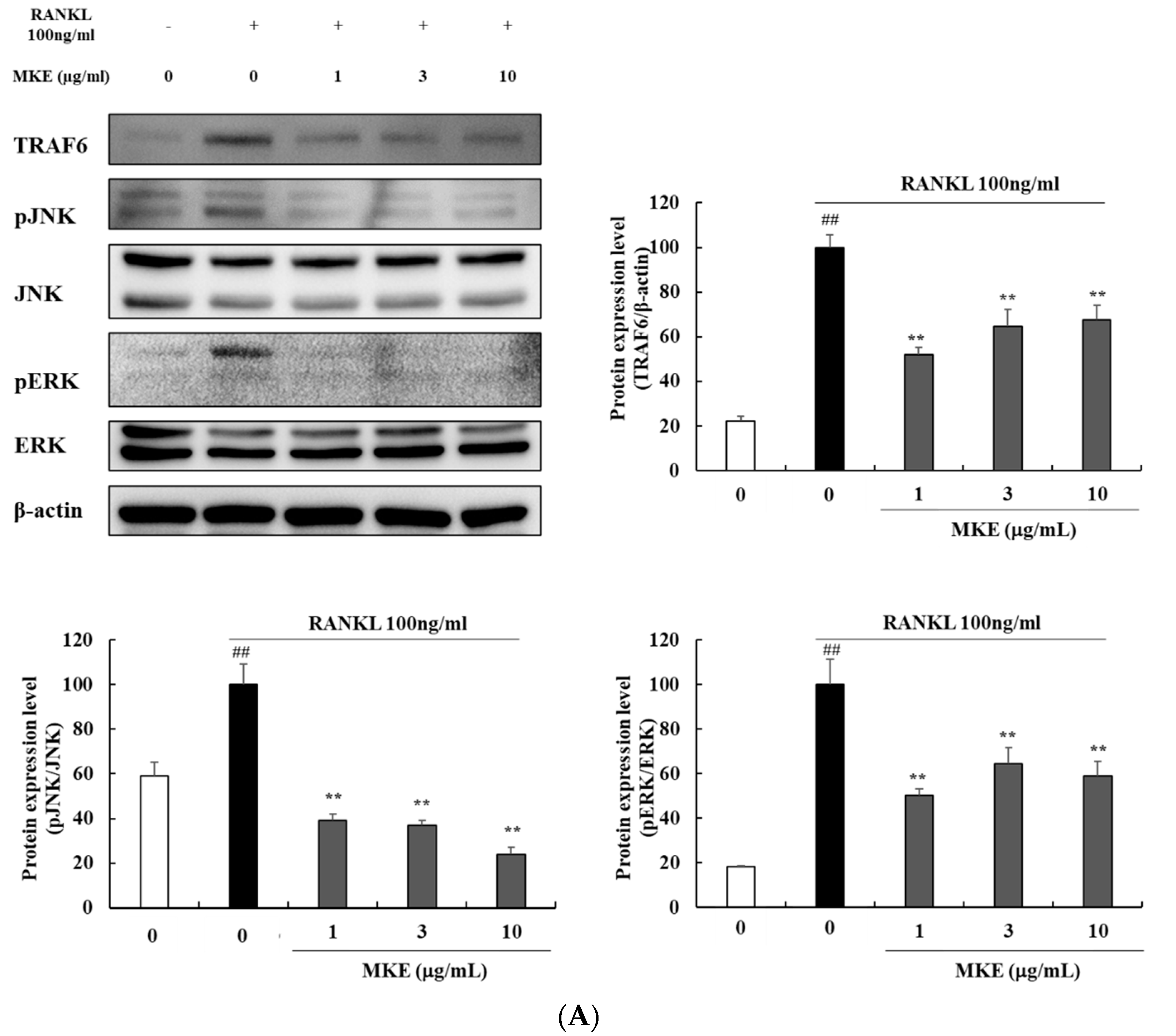
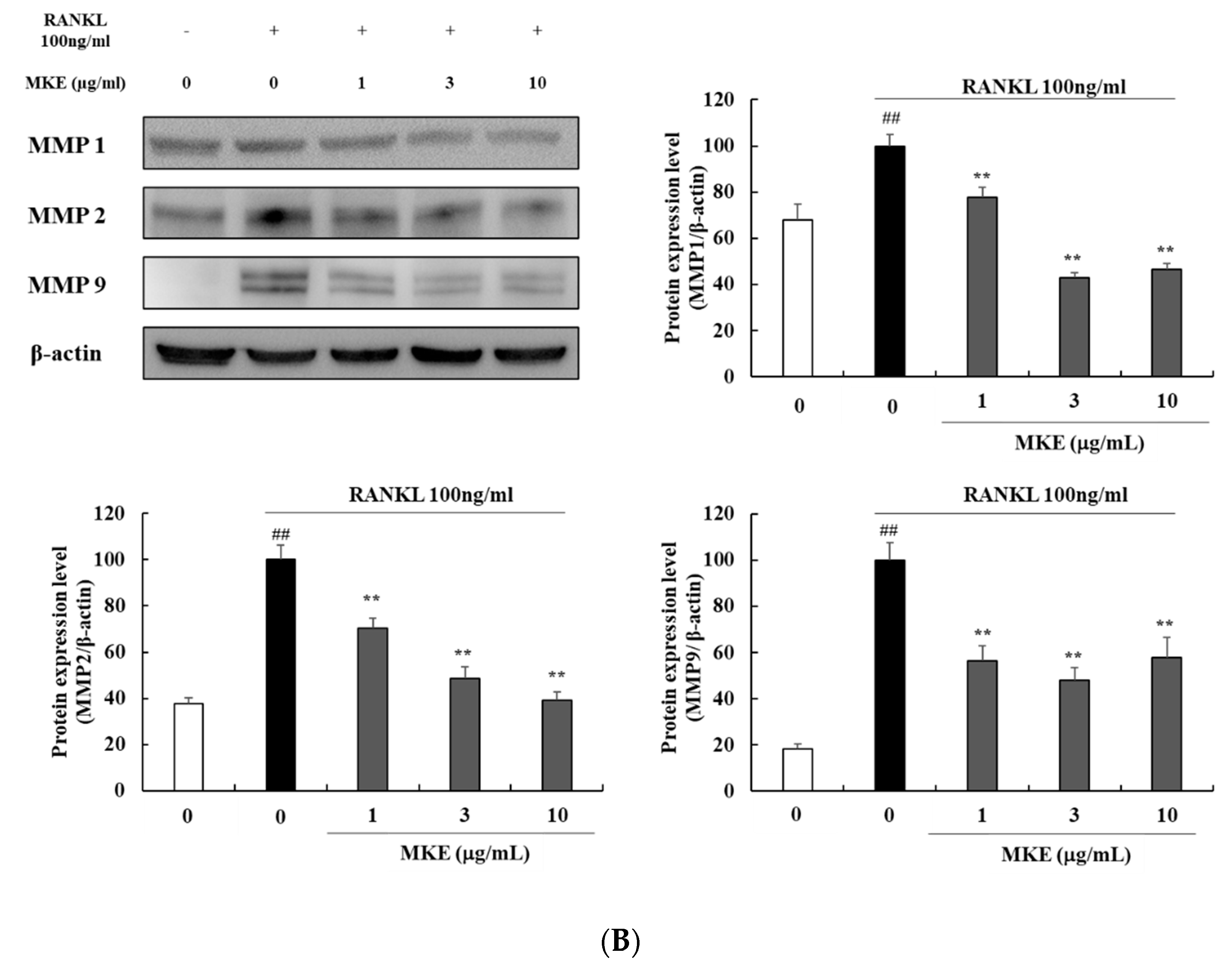
3.7. MKE Downregulates Osteoclast Differentiation-Related Protein Expression in RANKL-Stimulated RAW264.7 Cells
3.8. MKE Downregulates TRAF6 and MAPK Protein Expression in RANKL-Stimulated RAW264.7 Cells
3.9. MKE Downregulates MMP Protein Expression in RANKL-Stimulated RAW264.7 Cells
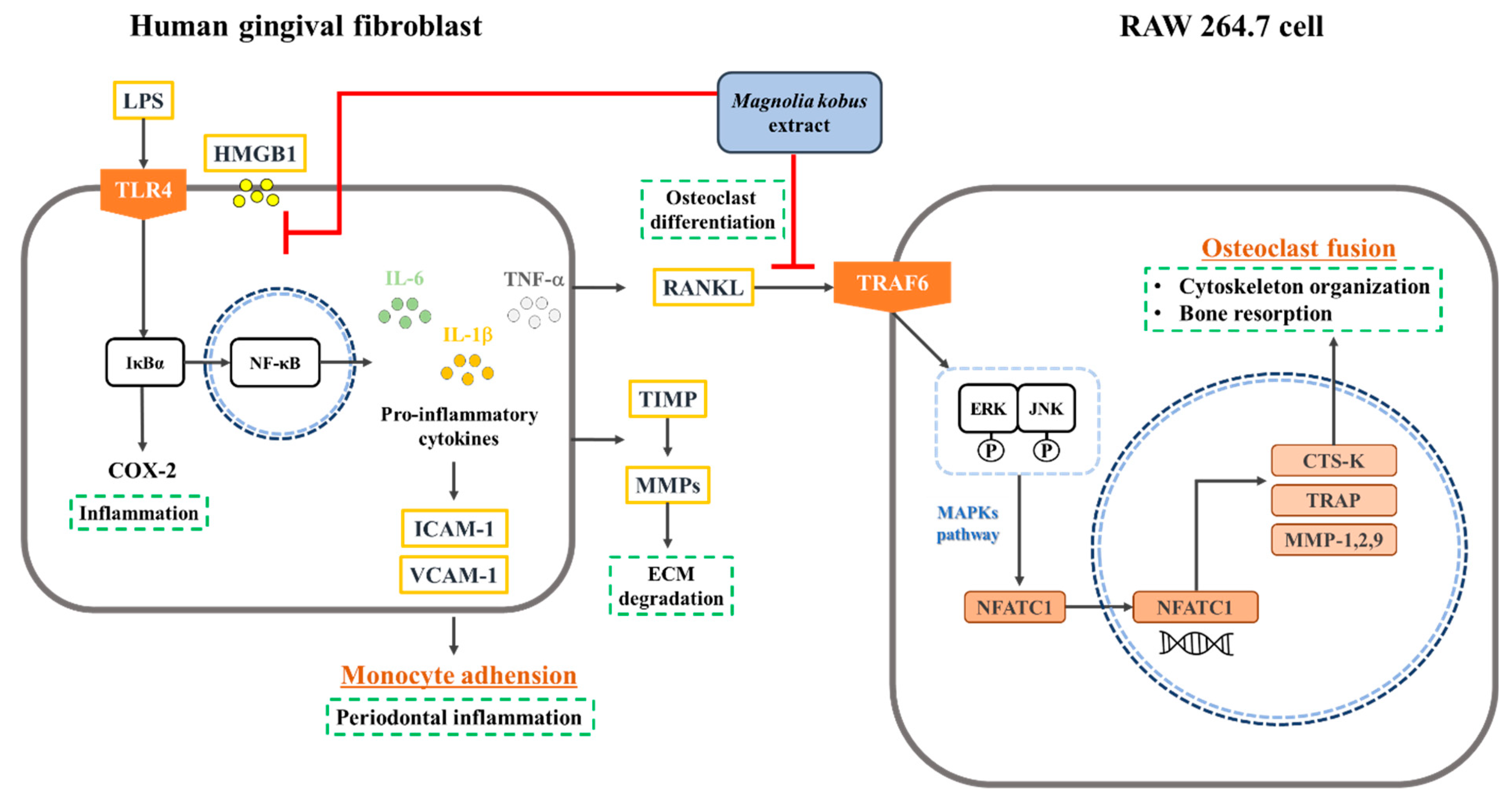
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, H.; Kim, M.-B.; Kim, C.; Hwang, J.-K. Inhibitory effects of panduratin A on periodontitis-induced inflammation and osteoclastogenesis through inhibition of MAPK pathways in vitro. J. Microbiol. Biotechnol. 2018, 28, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, X.; Luan, Q.-X. Hyperresponsiveness of human gingival fibroblasts from patients with aggressive periodontitis against bacterial lipopolysaccharide. Exp. Ther. Med. 2021, 21, 417. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, Q.; Qin, X.; Li, J.; Zhao, Y.; Xia, Y. Superparamagnetic Iron Oxide Nanoparticles Protect Human Gingival Fibroblasts from Porphyromonas gingivalis Invasion and Inflammatory Stimulation. Int. J. Nanomed. 2022, 17, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Herath, T.D.; Wang, Y.; Seneviratne, C.J.; Darveau, R.P.; Wang, C.-Y.; Jin, L. The expression and regulation of matrix metalloproteinase-3 is critically modulated by Porphyromonas gingivalis lipopolysaccharide with heterogeneous lipid A structures in human gingival fibroblasts. BMC Microbiol. 2013, 13, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, H.-S.; Kim, H.J.; Park, C.-M. Anti-inflammatory activity of various seaweeds in LPS-PG stimulated HGF-1 cells. J. Korean Soc. Oral Health Sci. 2020, 8, 25–30. [Google Scholar] [CrossRef]
- Park, C.M.; Yoon, H.-S. Chlorogenic acid as a positive regulator in LPS-PG-induced inflammation via TLR4/MyD88-mediated NF-κB and PI3K/MAPK signaling cascades in human gingival fibroblasts. Mediat. Inflamm. 2022, 2022, 2127642. [Google Scholar] [CrossRef]
- AlQranei, M.S.; Chellaiah, M.A. Osteoclastogenesis in periodontal diseases: Possible mediators and mechanisms. J. Oral Biosci. 2020, 62, 123–130. [Google Scholar] [CrossRef]
- Usui, M.; Onizuka, S.; Sato, T.; Kokabu, S.; Ariyoshi, W.; Nakashima, K. Mechanism of alveolar bone destruction in periodontitis—Periodontal bacteria and inflammation. Jpn. Dent. Sci. Rev. 2021, 57, 201–208. [Google Scholar] [CrossRef]
- Kang, W.; Wang, T.; Hu, Z.; Liu, F.; Sun, Y.; Ge, S. Metformin inhibits Porphyromonas gingivalis lipopolysaccharide-influenced inflammatory response in human gingival fibroblasts via regulating activating transcription factor-3 expression. J. Periodontol. 2017, 88, e169–e178. [Google Scholar] [CrossRef]
- Huang, J.; Xiong, T.; Zhang, Z.; Tan, Y.; Guo, L. Inhibition of the receptor for advanced glycation inhibits lipopolysaccharide-mediated High mobility group protein B1 and Interleukin-6 synthesis in human gingival fibroblasts through the NF-κB signaling pathway. Arch. Oral Biol. 2019, 105, 81–87. [Google Scholar] [CrossRef]
- Cronin, J.G.; Turner, M.L.; Goetze, L.; Bryant, C.E.; Sheldon, I.M. Toll-like receptor 4 and MYD88-dependent signaling mechanisms of the innate immune system are essential for the response to lipopolysaccharide by epithelial and stromal cells of the bovine endometrium. Biol. Reprod. 2012, 86, 51–59. [Google Scholar] [CrossRef]
- Chitanuwat, A.; Laosrisin, N.; Dhanesuan, N. Role of HMGB1 in proliferation and migration of human gingival and periodontal ligament fibroblasts. J. Oral Sci. 2013, 55, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramya, R.; Appukuttan, D.; Subramanian, S.; Prakash, P. Role of High Mobility Group Box 1 and Receptor for Advanced Glycation End Products in the Periodontal Disease Pathogenesis-A Review. J. Clin. Diagn. Res. 2022, 16, 6. [Google Scholar] [CrossRef]
- Kim, J.E.; Takanche, J.S.; Yun, B.S.; Yi, H.K. Anti-inflammatory character of Phelligridin D modulates periodontal regeneration in l ipopolysaccharide-induced human periodontal ligament cells. J. Periodontal Res. 2018, 53, 816–824. [Google Scholar] [CrossRef]
- Pinte, S.; Caetano, B.; Le Bras, A.; Havet, C.; Villain, G.; Dernayka, R.; Duez, C.; Mattot, V.; Soncin, F. Endothelial cell activation is regulated by epidermal growth factor-like domain 7 (Egfl7) during inflammation. J. Biol. Chem. 2016, 291, 24017–24028. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Nakae, H.; Matsuo, T. Cytokines differentially regulate ICAM-1 and VCAM-1 expression on human gingival fibroblasts. Clin. Exp. Immunol. 2006, 144, 494–502. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.-H.; Ning, W.-C. Evaluation of ICAM-1 and VCAM-1 gene polymorphisms in patients with periodontal disease. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 2386. [Google Scholar] [CrossRef] [Green Version]
- Ra, H.-J.; Parks, W.C. Control of matrix metalloproteinase catalytic activity. Matrix Biol. 2007, 26, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.-Y. Inhibitory effect of myricetin on matrix metalloproteinase expression and activity in periodontal inflammation. Int. J. Oral Biol. 2016, 41, 163–173. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Lee, N.K.; Lee, S.Y. Current understanding of RANK signaling in osteoclast differentiation and maturation. Mol. Cells 2017, 40, 706. [Google Scholar] [PubMed] [Green Version]
- Gravallese, E.M.; Galson, D.L.; Goldring, S.R.; Auron, P.E. The role of TNF-receptor family members and other TRAF-dependent receptors in bone resorption. Arthritis Res. Ther. 2000, 3, 1–7. [Google Scholar]
- Darnay, B.G.; Haridas, V.; Ni, J.; Moore, P.A.; Aggarwal, B.B. Characterization of the intracellular domain of receptor activator of NF-κB (RANK): Interaction with tumor necrosis factor receptor-associated factors and activation of NF-κB and c-Jun N-terminal kinase. J. Biol. Chem. 1998, 273, 20551–20555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Woo, K.; Kim, S.; Kim, H.-M.; Kwack, K.; Lee, Z.; Kim, H.-H. The phosphatidylinositol 3-kinase, p38, and extracellular signal-regulated kinase pathways are involved in osteoclast differentiation. Bone 2002, 30, 71–77. [Google Scholar] [CrossRef]
- Yan, L.; Lu, L.; Hu, F.; Shetti, D.; Wei, K. Piceatannol attenuates RANKL-induced osteoclast differentiation and bone resorption by suppressing MAPK, NF-κB and AKT signalling pathways and promotes Caspase3-mediated apoptosis of mature osteoclasts. R. Soc. Open Sci. 2019, 6, 190360. [Google Scholar] [CrossRef] [Green Version]
- Chiou, W.-F.; Huang, Y.-L.; Liu, Y.-W. (+)-Vitisin A inhibits osteoclast differentiation by preventing TRAF6 ubiquitination and TRAF6-TAK1 formation to suppress NFATc1 activation. PLoS ONE 2014, 9, e89159. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.-H.; Hsia, Y.-J.; Shih, K.-C.; Chou, T.-C. Fucoidan prevents RANKL-stimulated osteoclastogenesis and LPS-induced inflammatory bone loss via regulation of Akt/GSK3β/PTEN/NFATc1 signaling pathway and calcineurin activity. Mar. Drugs 2019, 17, 345. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Lee, D.-R.; Choi, B.-K.; Yang, S.H. Antiperiodontitis effects of magnolia biondii extract on ligature-induced periodontitis in Rats. Nutrients 2019, 11, 934. [Google Scholar] [CrossRef] [Green Version]
- Oshima, N.; Kume, H.; Umeda, T.; Takito, H.; Tsukimoto, M.; Hada, N. Structures and Inhibitory Activities for Interleukin-2 Production of Seasonally Variable Constituents in Flower Parts of Magnolia kobus at Different Growth Stages. Chem. Pharm. Bull. 2020, 68, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Pang, E.C.; Xue, C.C.; Zhao, Z.; Lin, J.; Li, C.G. Inhibitions of mast cell-derived histamine release by different Flos Magnoliae species in rat peritoneal mast cells. Phytomedicine 2008, 15, 808–814. [Google Scholar] [CrossRef]
- Yang, Y.J.; Park, J.I.; Lee, H.-J.; Seo, S.-M.; Lee, O.-K.; Choi, D.-H.; Paik, K.-H.; Lee, M.K. Effects of (+)-eudesmin from the stem bark of magnolia kobus DC. var. borealis Sarg. on neurite outgrowth in PC12 cells. Arch. Pharmacal Res. 2006, 29, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Lee, K.H.; Han, M.H.; Lee, H.; Ahn, J.M.; Han, S.B.; Han, G.; Lee, K.; Park, S.K.; Kim, H.M. Antiinflammatory activity of methanol extract isolated from stem bark of Magnolia kobus. Phytother. Res. 2008, 22, 883–888. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Lee, S.-J.; Lee, S.-K.; Choi, B.-K.; Lee, D.-R. Magnolia kobus Extract Inhibits Periodontitis-Inducing Mediators in Porphyromonas gingivalis Lipopolysaccharide-Activated RAW 264.7 Cells. Curr. Issues Mol. Biol. 2023, 45, 538–554. [Google Scholar] [CrossRef]
- Hienz, S.A.; Paliwal, S.; Ivanovski, S. Mechanisms of bone resorption in periodontitis. J. Immunol. Res. 2015, 2015, 615486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naruishi, K. Biological Roles of Fibroblasts in Periodontal Diseases. Cells 2022, 11, 3345. [Google Scholar] [CrossRef] [PubMed]
- Baek, K.J.; Choi, Y.; Ji, S. Gingival fibroblasts from periodontitis patients exhibit inflammatory characteristics in vitro. Arch. Oral Biol. 2013, 58, 1282–1292. [Google Scholar] [CrossRef]
- Zhou, Z.; Han, J.Y.; Xi, C.X.; Xie, J.X.; Feng, X.; Wang, C.Y.; Mei, L.; Xiong, W.C. HMGB1 regulates RANKL-induced osteoclastogenesis in a manner dependent on RAGE. J. Bone Miner. Res. 2008, 23, 1084–1096. [Google Scholar] [CrossRef]
- Kwang-Hoon, C.; Chul, J.H.; Sung, K.K.; Tong-Shin, C.; Seo, H.G. Poncirin Inhibits Osteoclast Differentiation and Bone Loss Through Down-Regulation of NFATc1 In Vitro and In Vivo. Biomol. Ther. 2020, 28, 337. [Google Scholar]
- Omi, M.; Mishina, Y. Roles of osteoclasts in alveolar bone remodeling. Genesis 2022, 60, e23490. [Google Scholar] [CrossRef]
- Kim, J.-H.; Ki, J.-Y.; Ann, J.-Y.; Park, H.-J.; Kim, H.-J.; Kwak, H.-B.; Oh, J.-M.; Kim, Y.-K. Inhibitory effects of Achyranthis bidentatae radix on osteoclast differentiation and bone resorption. Korea J. Herbol. 2010, 25, 65–74. [Google Scholar]
- Seo, M.; Baek, M.; Lee, H.J.; Shin, Y.P.; Lee, J.H.; Kim, I.-W.; Kim, M.; Hwang, J.-S. Inhibitory Effects of Tenebrio molitor Larvae Ethanol Extract on RANKL-Induced Osteoclast Differentiation. J. Life Sci. 2020, 30, 983–989. [Google Scholar]
- Choi, K.-H.; Yoo, D.-Y. The effect of Guibi-tang water extract on osteoclast differentiation and osteoblast proliferation. J. Korean Obstet. Gynecol. 2014, 27, 12–27. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.-Y. Myricetin suppresses LPS-induced MMP expression in human gingival fibroblasts and inhibits osteoclastogenesis by downregulating NFATc1 in RANKL-induced RAW 264.7 cells. Arch. Oral Biol. 2012, 57, 1623–1632. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Conditions |
|---|---|---|
| HMGB1 | F: CGC CAT GAG AAC TTC CTA CC R: CAC TTG GCC TTC CCT CTG TA | 40 cycles at the following conditions: 94 °C for 40 s, 55 °C for 40 s, and 72 °C for 1 min |
| ICAM-1 | F: CGT GCC GCA CTG AAC TGG AC R: CCT CAC ACT TCA CG TCA CCT | |
| VCAM-1 | F: ATT GGG AAA AAC AGA AAA GAG R: GGC AAC ATT GAC ATA AAG T | |
| TIMP-1 | F: TTC GTG GGG ACA CCA GAA GTC AAC R: TGG ACA CTG TGC AGG CTT CAG TTC | |
| TIMP-2 | F: AAG CGG TCA GTG AGA AGG AGT GG R: CCT TGG AGG CTT TTT TGC AGT TG | |
| GAPDH | F: TGA AGG TCG GAG TCA ACG GAT TTG GT R: CAT GTG GGC CAT GAG GTC CAC CAC | |
| NFATc1 | F: CCGTTGCTTCCAGAAAATAACA R: TGTGGGATGTGAACTCGGAA | 40 cycles at the following conditions: 95 °C for 5 s, 60 °C for 20 s, and 72 °C for 20 s |
| TRAP | F: CTGGAGTGCACGATGCCAGCGACA R: TCCGTGCTCGGCGATGGACCAGA | |
| GAPDH | F: ACCCAGAAGACTGTGGATGG R: CACATTGGGGGTAGGAACAC | |
| MMP9 | F: GCCCTGGAACTCACACGACA R: TCCGTGCTCGGCGATGGACCAGA | 35 cycles at 95 °C for 5 s and 60 °C for 20 s |
| Catehpsin K | F: CAGCAGAACGGAGGCATTGA R: CCTTTGCCGTGGCGTTATAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.J.; Lee, S.J.; Lee, S.K.; Choi, B.K.; Lee, D.R.; Park, J.-H.; Oh, J.S. Magnolia kobus Extract Suppresses Porphyromonas gingivalis LPS-Induced Proinflammatory Cytokine and MMP Expression in HGF-1 Cells and Regulates Osteoclastogenesis in RANKL-Stimulated RAW264.7 Cells. Curr. Issues Mol. Biol. 2023, 45, 4875-4890. https://doi.org/10.3390/cimb45060310
Lee HJ, Lee SJ, Lee SK, Choi BK, Lee DR, Park J-H, Oh JS. Magnolia kobus Extract Suppresses Porphyromonas gingivalis LPS-Induced Proinflammatory Cytokine and MMP Expression in HGF-1 Cells and Regulates Osteoclastogenesis in RANKL-Stimulated RAW264.7 Cells. Current Issues in Molecular Biology. 2023; 45(6):4875-4890. https://doi.org/10.3390/cimb45060310
Chicago/Turabian StyleLee, Hae Jin, So Jung Lee, Sung Kwon Lee, Bong Keun Choi, Dong Ryung Lee, Ju-Hyoung Park, and Joa Sub Oh. 2023. "Magnolia kobus Extract Suppresses Porphyromonas gingivalis LPS-Induced Proinflammatory Cytokine and MMP Expression in HGF-1 Cells and Regulates Osteoclastogenesis in RANKL-Stimulated RAW264.7 Cells" Current Issues in Molecular Biology 45, no. 6: 4875-4890. https://doi.org/10.3390/cimb45060310
APA StyleLee, H. J., Lee, S. J., Lee, S. K., Choi, B. K., Lee, D. R., Park, J. -H., & Oh, J. S. (2023). Magnolia kobus Extract Suppresses Porphyromonas gingivalis LPS-Induced Proinflammatory Cytokine and MMP Expression in HGF-1 Cells and Regulates Osteoclastogenesis in RANKL-Stimulated RAW264.7 Cells. Current Issues in Molecular Biology, 45(6), 4875-4890. https://doi.org/10.3390/cimb45060310