
Synergistic Chemopreventive Effects of a Novel Combined Plant Extract Comprising Gallic Acid and Hesperidin on Colorectal Cancer


 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. HPT Extraction
2.3. Measurement of Phytochemicals
2.4. HPLC Analysis of Gallic Acid and Hesperidin
2.5. Cell Proliferation Assay
2.6. Cell-Cycle Analysis
2.7. Immunoblotting
2.8. Spheroid Formation in Soft Agar
2.9. Tissue Processing and Immunohistochemical Staining
2.10. Statistical Analysis
3. Results
3.1. EA Extract of HPT Contained the Most Abundant Polyphenols
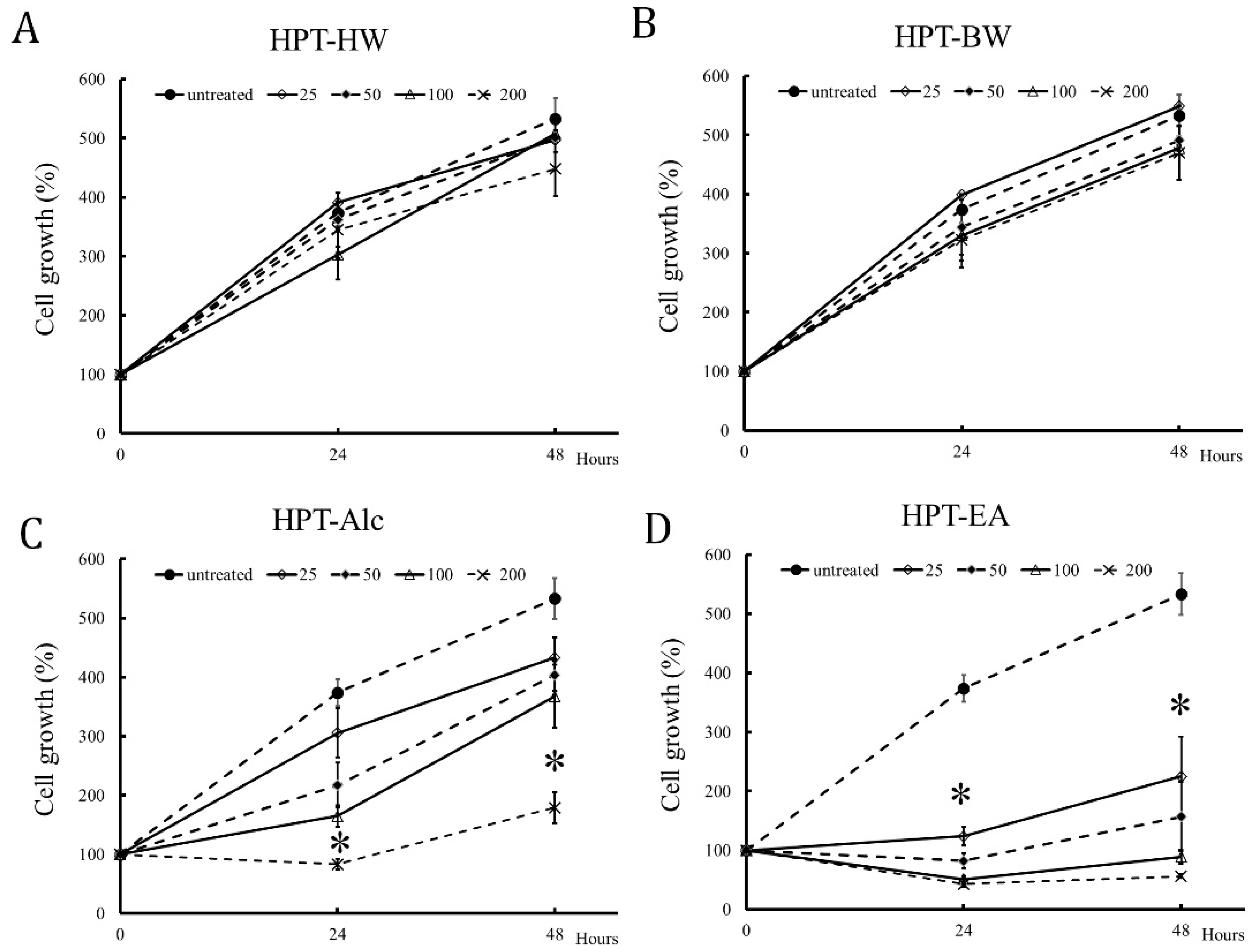
3.2. HPT-Alc and HPT-EA Treatment Inhibited HT-29 Cell Growth in a Dose-Dependent Manner
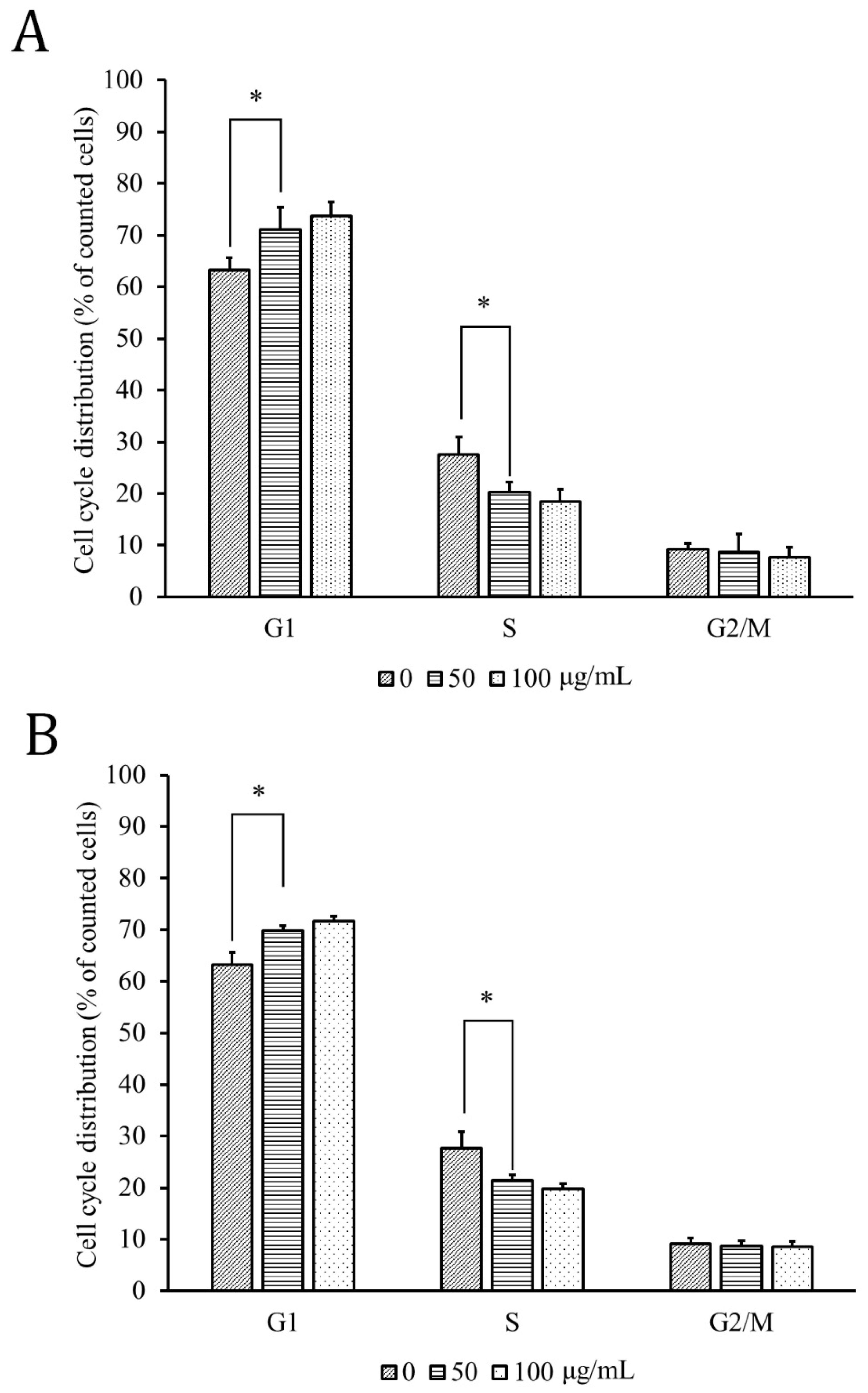
3.3. HPT Retarded the G1 Phase of the Cell Cycle
3.4. HPT Treatment Attenuates Cyclins D1 and E and Accentuates Cip1/p21 Expression via G1-Phase-Controlling Protein Mechanism
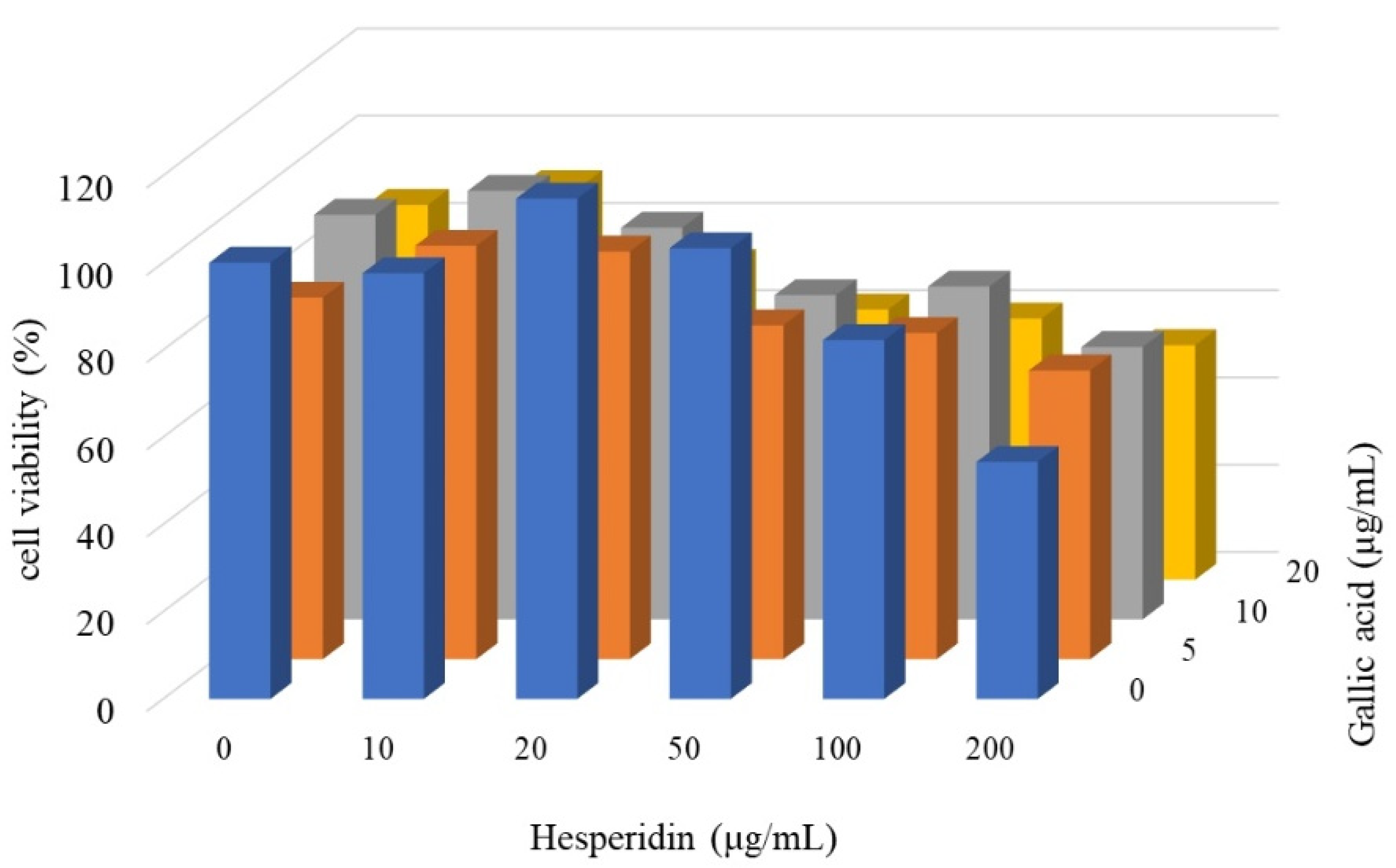
3.5. Synergistic Inhibition Effect of Gallic Acid and Hesperidin on CRC Cell Proliferation
3.6. Inhibition of In Vitro Tumor Growth by Combined Gallic Acid and Hesperidin Treatment
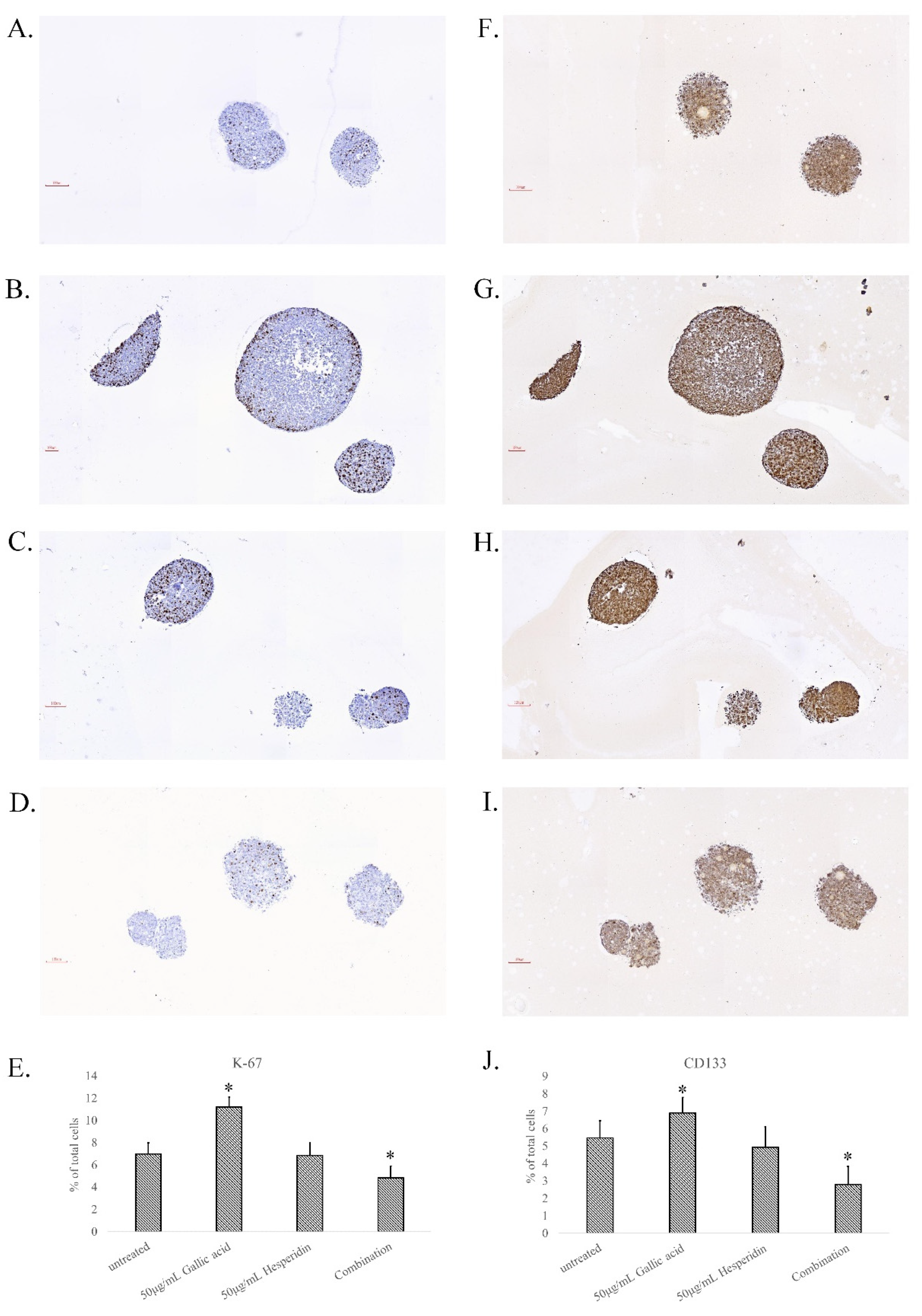
3.7. Cell Division and Stemness in Gallic Acid and Hesperidin Combination-Treated Spheroids
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tamang, L.D.; Kim, B.W. Deep Learning Approaches to Colorectal Cancer Diagnosis: A Review. Appl. Sci. 2021, 11, 10982. [Google Scholar] [CrossRef]
- Goh, J.X.H.; Tan, L.T.-H.; Goh, J.K.; Chan, K.G.; Pusparajah, P.; Lee, L.-H.; Goh, B.-H. Nobiletin and Derivatives: Functional Compounds from Citrus Fruit Peel for Colon Cancer Chemoprevention. Cancers 2019, 11, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekundayo, B.E.; Obafemi, T.O.; Afolabi, B.A.; Adewale, O.B.; Onasanya, A.; Osukoya, O.A.; Falode, J.A.; Akintayo, C.; Adu, I.A. Gallic Acid and Hesperidin Elevate Neurotransmitters Level and Protect against Oxidative Stress, Inflammation and Apoptosis in Aluminum Chloride-Induced Alzheimer’s Disease in Rats. Pharmacol. Res. Mod. Chin. Med. 2022, 5, 100193. [Google Scholar] [CrossRef]
- Ho, S.-C.; Hwang, L.S.; Shen, Y.-J.; Lin, C.-C. Suppressive Effect of a Proanthocyanidin-Rich Extract from Longan (Dimocarpus Longan Lour.) Flowers on Nitric Oxide Production in LPS-Stimulated Macrophage Cells. J. Agric. Food Chem. 2007, 55, 10664–10670. [Google Scholar] [CrossRef]
- Chung, Y.-C.; Lin, C.-C.; Chou, C.-C.; Hsu, C.-P. The Effect of Longan Seed Polyphenols on Colorectal Carcinoma Cells. Eur. J. Clin. Investig. 2010, 40, 713–721. [Google Scholar] [CrossRef]
- Hsu, C.-P.; Lin, C.-C.; Huang, C.-C.; Lin, Y.-H.; Chou, J.-C.; Tsia, Y.-T.; Su, J.-R.; Chung, Y.-C. Induction of Apoptosis and Cell Cycle Arrest in Human Colorectal Carcinoma by Litchi Seed Extract. J. Biomed. Biotechnol. 2012, 2012, 341479. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-P.; Lin, Y.-H.; Zhou, S.-P.; Chung, Y.-C.; Lin, C.-C.; Wang, S.-C. Longan Flower Extract Inhibits the Growth of Colorectal Carcinoma. Nutr. Cancer 2010, 62, 229–236. [Google Scholar] [CrossRef]
- Hsu, C.-P.; Shih, Y.-T.; Lin, B.-R.; Chiu, C.-F.; Lin, C.-C. Inhibitory Effect and Mechanisms of an Anthocyanins- and Anthocyanidins-Rich Extract from Purple-Shoot Tea on Colorectal Carcinoma Cell Proliferation. J. Agric. Food Chem. 2012, 60, 3686–3692. [Google Scholar] [CrossRef]
- Kern, D.H.; Morgan, C.R.; Hildebrand-Zanki, S.U. In Vitro Pharmacodynamics of 1-Beta-D-Arabinofuranosylcytosine: Synergy of Antitumor Activity with Cis-Diamminedichloroplatinum(II). Cancer Res. 1988, 48, 117–121. [Google Scholar]
- Chung, Y.-C.; Ku, Y.-L.; Chiang, H.-C.; Liu, W.-C.; Kao, T.-Y.; Yang, C.-H.; Huang, C.-C.; Hsu, C.-P. Antibody to Interleukin-6 Receptor Inhibits in Vivo Growth of Human Colorectal Carcinoma Cell Xenografts. Anticancer Res. 2021, 41, 4907–4916. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Xu, F.; Han, Y.; Hernández-Ledesma, B.; Xiao, H. Inhibitory Effects of Peptide Lunasin in Colorectal Cancer HCT-116 Cells and Their Tumorsphere-Derived Subpopulation. Int. J. Mol. Sci. 2020, 21, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Z.; Tang, P.; Liu, B.; Ran, C.; Yuan, C.; Zhang, Y.; Lu, Y.; Duan, X.; Yang, Y.; Wu, H. Ferroptosis-Related Genes for Overall Survival Prediction in Patients with Colorectal Cancer Can Be Inhibited by Gallic Acid. Int. J. Biol. Sci. 2021, 17, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Rosman, R.; Saifullah, B.; Maniam, S.; Dorniani, D.; Hussein, M.; Fakurazi, S. Improved Anticancer Effect of Magnetite Nanocomposite Formulation of Gallic Acid (Fe3O4-PEG-GA) against Lung, Breast and Colon Cancer Cells. Nanomaterials 2018, 8, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani, A.; Azarian, M.M.S.; Ibrahim, W.N.; Hussain, S.A. Preparation, Characterization and Therapeutic Properties of Gum Arabic-Stabilized Gallic Acid Nanoparticles. Sci. Rep. 2020, 10, 17808. [Google Scholar] [CrossRef]
- Subramanian, A.P.; Jaganathan, S.K.; Mandal, M.; Supriyanto, E.; Muhamad, I.I. Gallic Acid Induced Apoptotic Events in HCT-15 Colon Cancer Cells. World J. Gastroenterol. 2016, 22, 3952–3961. [Google Scholar] [CrossRef]
- Tanaka, T.; Tanaka, T.; Tanaka, M.; Kuno, T. Cancer Chemoprevention by Citrus Pulp and Juices Containing High Amounts of β-Cryptoxanthin and Hesperidin. J. Biomed. Biotechnol. 2012, 2012, 516981. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Kohno, H.; Murakami, M.; Shimada, R.; Kagami, S.; Sumida, T.; Azuma, Y.; Ogawa, H. Suppression of Azoxymethane-Induced Colon Carcinogenesis in Male F344 Rats by Mandarin Juices Rich in Beta-Cryptoxanthin and Hesperidin. Int. J. Cancer 2000, 88, 146–150. [Google Scholar] [CrossRef]
- Brown, D.G.; Borresen, E.C.; Brown, R.J.; Ryan, E.P. Heat-Stabilised Rice Bran Consumption by Colorectal Cancer Survivors Modulates Stool Metabolite Profiles and Metabolic Networks: A Randomised Controlled Trial. Br. J. Nutr. 2017, 117, 1244–1256. [Google Scholar] [CrossRef] [Green Version]
- Hosseinimehr, S.J.; Jalayer, Z.; Naghshvar, F.; Mahmoudzadeh, A. Hesperidin Inhibits Cyclophosphamide-Induced Tumor Growth Delay in Mice. Integr. Cancer Ther. 2012, 11, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Tao, L. Weichang’an and 5-Fluorouracil Suppresses Colorectal Cancer in a Mouse Model. World J. Gastroenterol. 2015, 21, 1125. [Google Scholar] [CrossRef]
- Chung, Y.-C.; Lu, L.-C.; Tsai, M.-H.; Chen, Y.-J.; Chen, Y.-Y.; Yao, S.-P.; Hsu, C.-P. The Inhibitory Effect of Ellagic Acid on Cell Growth of Ovarian Carcinoma Cells. Evid.-Based Complement. Altern. Med. Ecam 2013, 2013, 306705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.-C.; Chen, C.-H.; Tsai, Y.-T.; Lin, C.-C.; Chou, J.-C.; Kao, T.-Y.; Huang, C.-C.; Cheng, C.-H.; Hsu, C.-P. Litchi Seed Extract Inhibits Epidermal Growth Factor Receptor Signaling and Growth of Two Non-Small Cell Lung Carcinoma Cells. BMC Complement. Altern. Med. 2017, 17, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-J.; Chung, Y.-C.; Chang, H.-L.; Chang, H.-P.; Chou, J.-L.; Lin, C.-C.; Chen, C.-H.; Hsu, C.-P. Synergistic Effect of Combined Treatment with Longan Flower Extract and 5-Fluorouracil on Colorectal Cancer Cells. Nutr. Cancer 2019, 72, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Sogawa, C.; Eguchi, T.; Namba, Y.; Okusha, Y.; Aoyama, E.; Ohyama, K.; Okamoto, K. Gel-Free 3D Tumoroids with Stem Cell Properties Modeling Drug Resistance to Cisplatin and Imatinib in Metastatic Colorectal Cancer. Cells 2021, 10, 344. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yamashita, K. ASO Author Reflections: ASCL2, a Marker of Dynamic Colon Stem Cell, Involved in Wnt Pathway Activation Reflects Anti-Cancer Drug Resistance in Primary Colorectal Cancer. Ann. Surg. Oncol. 2019, 26, 634. [Google Scholar] [CrossRef]
- Lazer, L.M.; Kesavan, Y.; Gor, R.; Ramachandran, I.; Pathak, S.; Narayan, S.; Anbalagan, M.; Ramalingam, S. Targeting Colon Cancer Stem Cells Using Novel Doublecortin like Kinase 1 Antibody Functionalized Folic Acid Conjugated Hesperetin Encapsulated Chitosan Nanoparticles. Colloids Surf. B Biointerfaces 2022, 217, 112612. [Google Scholar] [CrossRef]
- Hermawan, A.; Khumaira, A.; Ikawati, M.; Putri, H.; Jenie, R.I.; Angraini, S.M.; Muflikhasari, H.A. Identification of Key Genes of Hesperidin in Inhibition of Breast Cancer Stem Cells by Functional Network Analysis. Comput. Biol. Chem. 2021, 90, 107427. [Google Scholar] [CrossRef]
- Aggarwal, V.; Tuli, H.S.; Thakral, F.; Singhal, P.; Aggarwal, D.; Srivastava, S.; Pandey, A.; Sak, K.; Varol, M.; Khan, A.; et al. Molecular Mechanisms of Action of Hesperidin in Cancer: Recent Trends and Advancements. Exp. Biol. Med. 2020, 245, 486–497. [Google Scholar] [CrossRef] [Green Version]
- Abate, G.; Zhang, L.; Pucci, M.; Morbini, G.; Mac Sweeney, E.; Maccarinelli, G.; Ribaudo, G.; Gianoncelli, A.; Uberti, D.; Memo, M.; et al. Phytochemical Analysis and Anti-Inflammatory Activity of Different Ethanolic Phyto-Extracts of Artemisia annua L. Biomolecules 2021, 11, 975. [Google Scholar] [CrossRef]
- Gupta, A.K.; Dhua, S.; Sahu, P.P.; Abate, G.; Mishra, P.; Mastinu, A. Variation in Phytochemical, Antioxidant and Volatile Composition of Pomelo Fruit (Citrus grandis (L.) Osbeck) during Seasonal Growth and Development. Plants 2021, 10, 1941. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPT-BW | HPT-HW | HPT-Alc | HPT-EA | |
|---|---|---|---|---|
| Total phenols (mg/g) | 62.65 ± 4.13 | 58.92 ± 2.88 | 197.85 ± 13.41 * | 325.16 ± 23.73 * |
| Total flavonoids (mg/g) | 67.42 ± 5.06 | 88.48 ± 9.41 | 89.46 ± 2.68 | 84.39 ± 7.66 |
| Condensed tannin (mg/g) | 55.58 ± 9.77 | 97.78 ± 5.74 * | 127.54 ± 6.97 * | 107.33 ± 6.38 * |
| HPT-EA | HPT-Alc | |
|---|---|---|
| Gallic acid (mg/g) | 204.83 ± 18.93 * | 103.94 ± 9.32 |
| Hesperidin (mg/g) | 41.35 ± 5.23 * | 25.79 ± 2.38 |
| Hesperidin (μg/mL) | Gallic Acid (μg/mL) | |||
|---|---|---|---|---|
| 10 | 20 | 50 | 100 | |
| 10 | 0.85 | 0.92 | 0.93 | 1.07 |
| 20 | 1.02 | 1.19 | 1.37 | 1.18 |
| 50 | 1.12 | 1.29 | 1.43 | 1.09 |
| 100 | 0.91 | 1.00 | 1.18 | 0.87 |
| 200 | 0.68 | 0.81 | 0.87 | 0.71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-J.; Lu, J.-H.; Lin, C.-C.; Zeng, S.-W.; Chang, J.-F.; Chung, Y.-C.; Chang, H.; Hsu, C.-P. Synergistic Chemopreventive Effects of a Novel Combined Plant Extract Comprising Gallic Acid and Hesperidin on Colorectal Cancer. Curr. Issues Mol. Biol. 2023, 45, 4908-4922. https://doi.org/10.3390/cimb45060312
Chen S-J, Lu J-H, Lin C-C, Zeng S-W, Chang J-F, Chung Y-C, Chang H, Hsu C-P. Synergistic Chemopreventive Effects of a Novel Combined Plant Extract Comprising Gallic Acid and Hesperidin on Colorectal Cancer. Current Issues in Molecular Biology. 2023; 45(6):4908-4922. https://doi.org/10.3390/cimb45060312
Chicago/Turabian StyleChen, Szu-Jung, Jui-Hua Lu, Chih-Cheng Lin, Shao-Wei Zeng, Jia-Feng Chang, Yuan-Chiang Chung, Hsiang Chang, and Chih-Ping Hsu. 2023. "Synergistic Chemopreventive Effects of a Novel Combined Plant Extract Comprising Gallic Acid and Hesperidin on Colorectal Cancer" Current Issues in Molecular Biology 45, no. 6: 4908-4922. https://doi.org/10.3390/cimb45060312
APA StyleChen, S. -J., Lu, J. -H., Lin, C. -C., Zeng, S. -W., Chang, J. -F., Chung, Y. -C., Chang, H., & Hsu, C. -P. (2023). Synergistic Chemopreventive Effects of a Novel Combined Plant Extract Comprising Gallic Acid and Hesperidin on Colorectal Cancer. Current Issues in Molecular Biology, 45(6), 4908-4922. https://doi.org/10.3390/cimb45060312