
Molecular Markers of Ovarian Germ Cells of Banana Prawn (Fenneropenaeus merguiensis)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue-Specific Distribution of Gene Expression
2.3. Sequence Analysis
2.4. Cloning of Partial FmCyclinB, FmNanos, and FmNASP Genes
2.5. In Situ Hybridization (ISH)
2.6. Quantitative Reversed Transcript PCR (qRT-PCR)
2.7. Statistical Analysis
3. Results
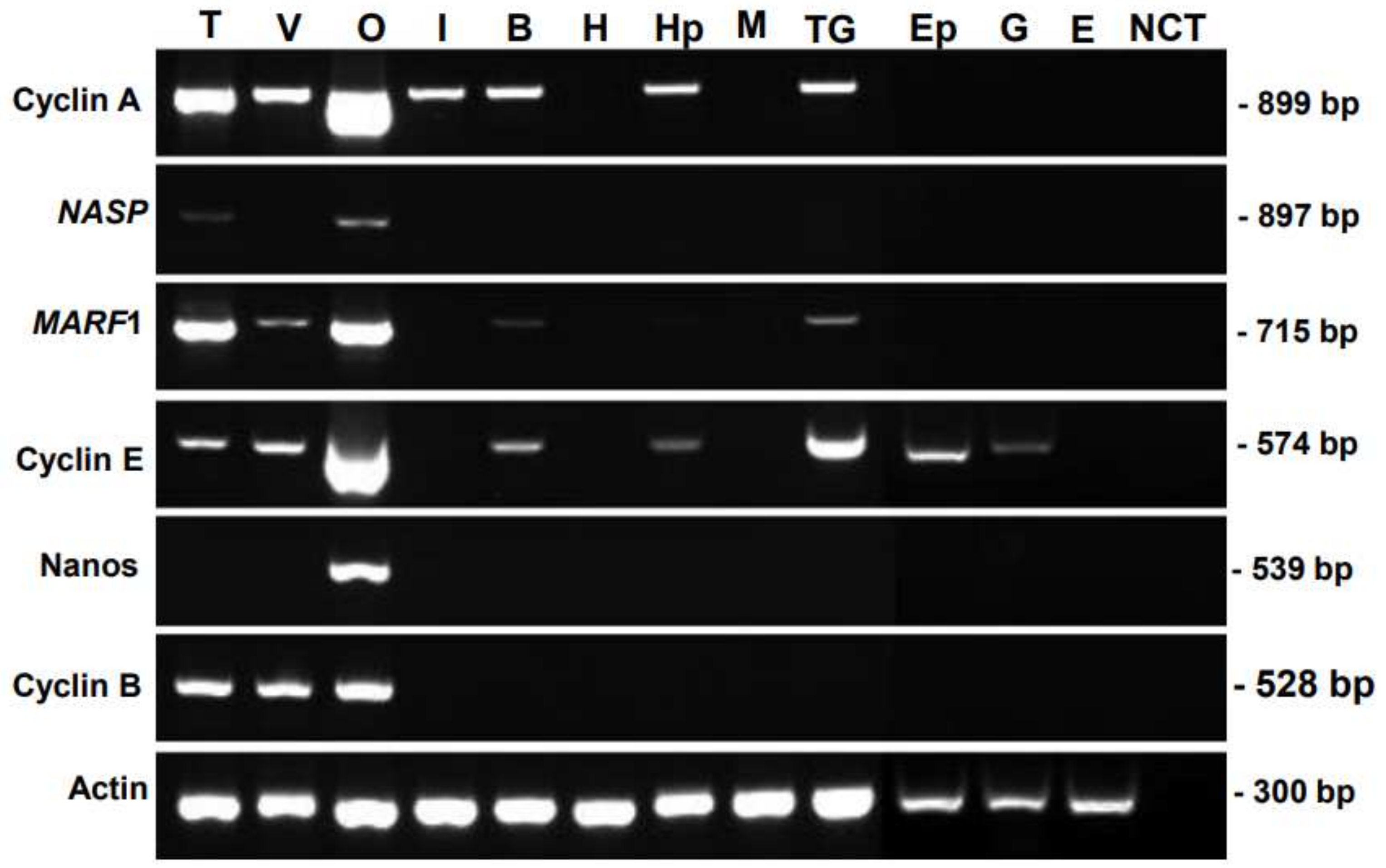
3.1. Tissue-Specific Distribution of Genes
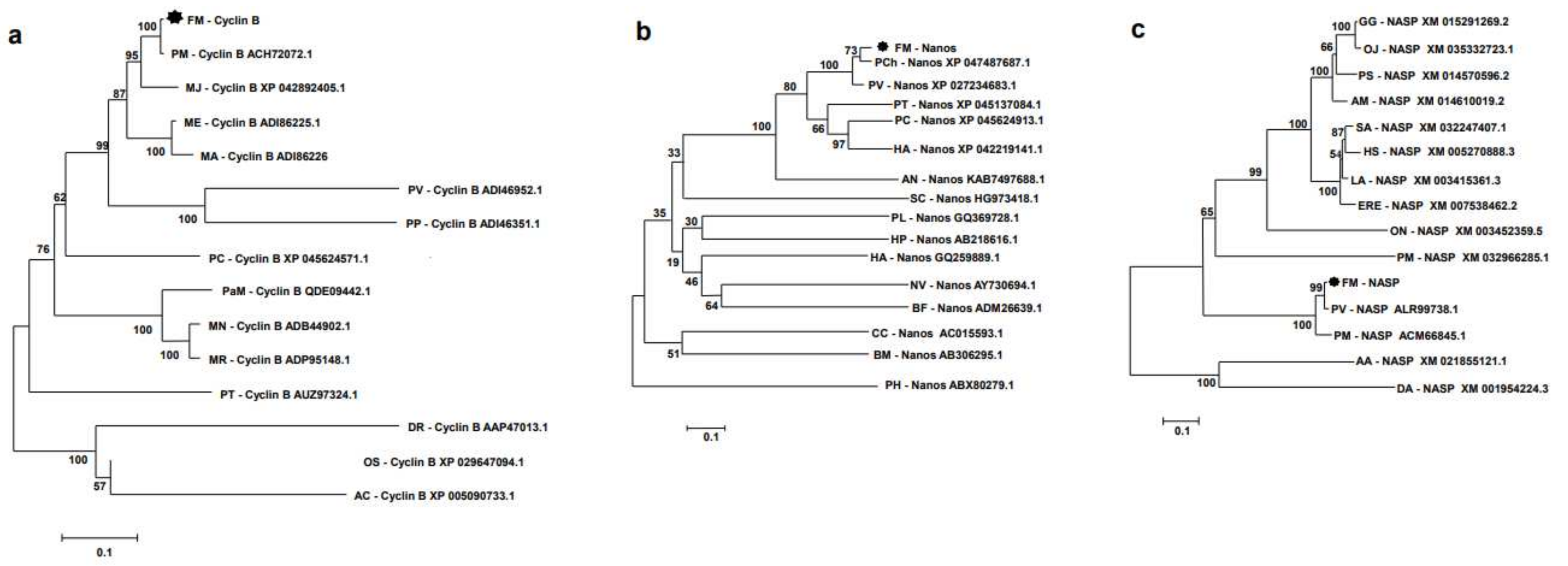
3.2. Sequence Analysis
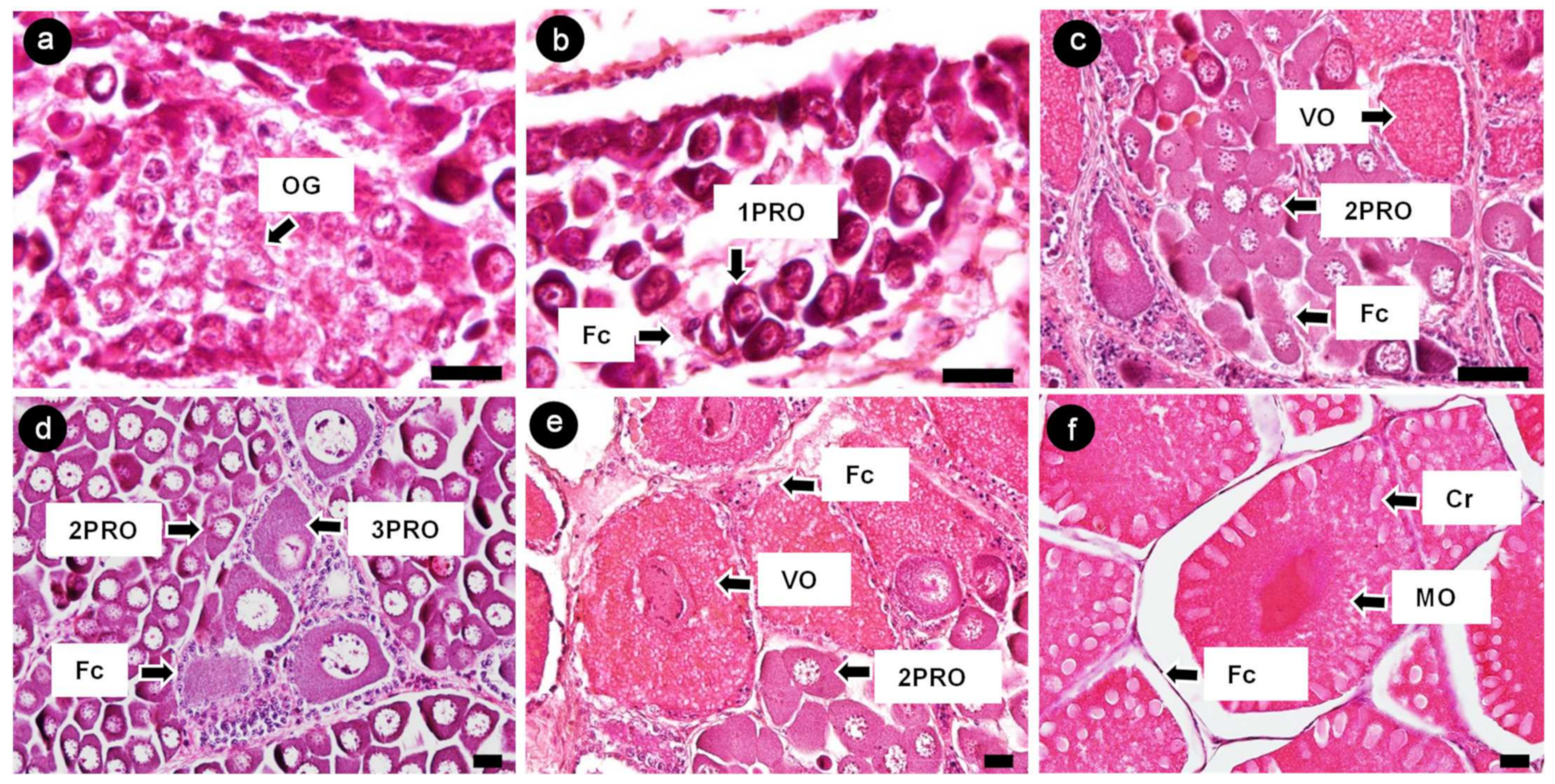
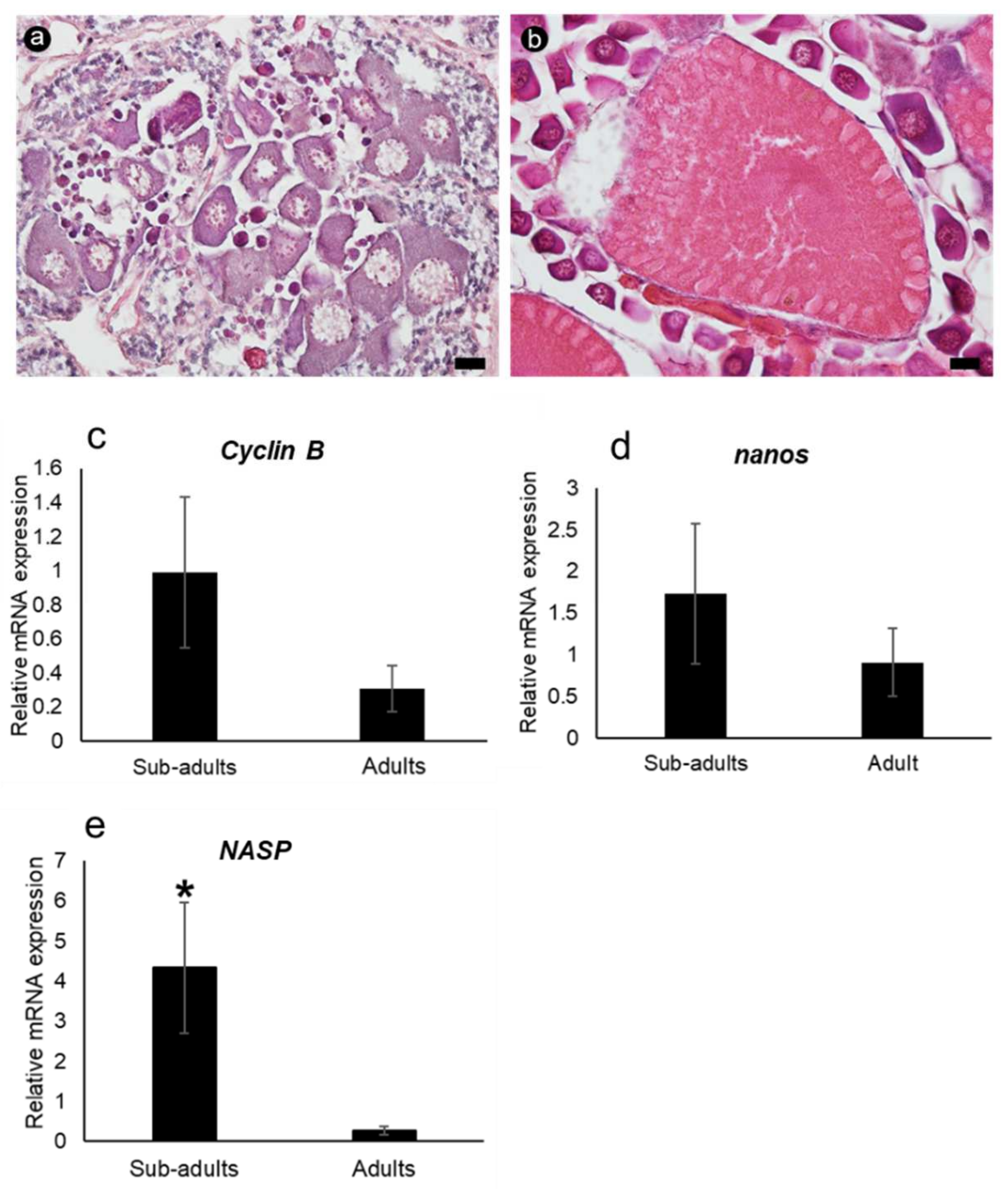
3.3. Characterization of Banana Prawn Ovarian Cells
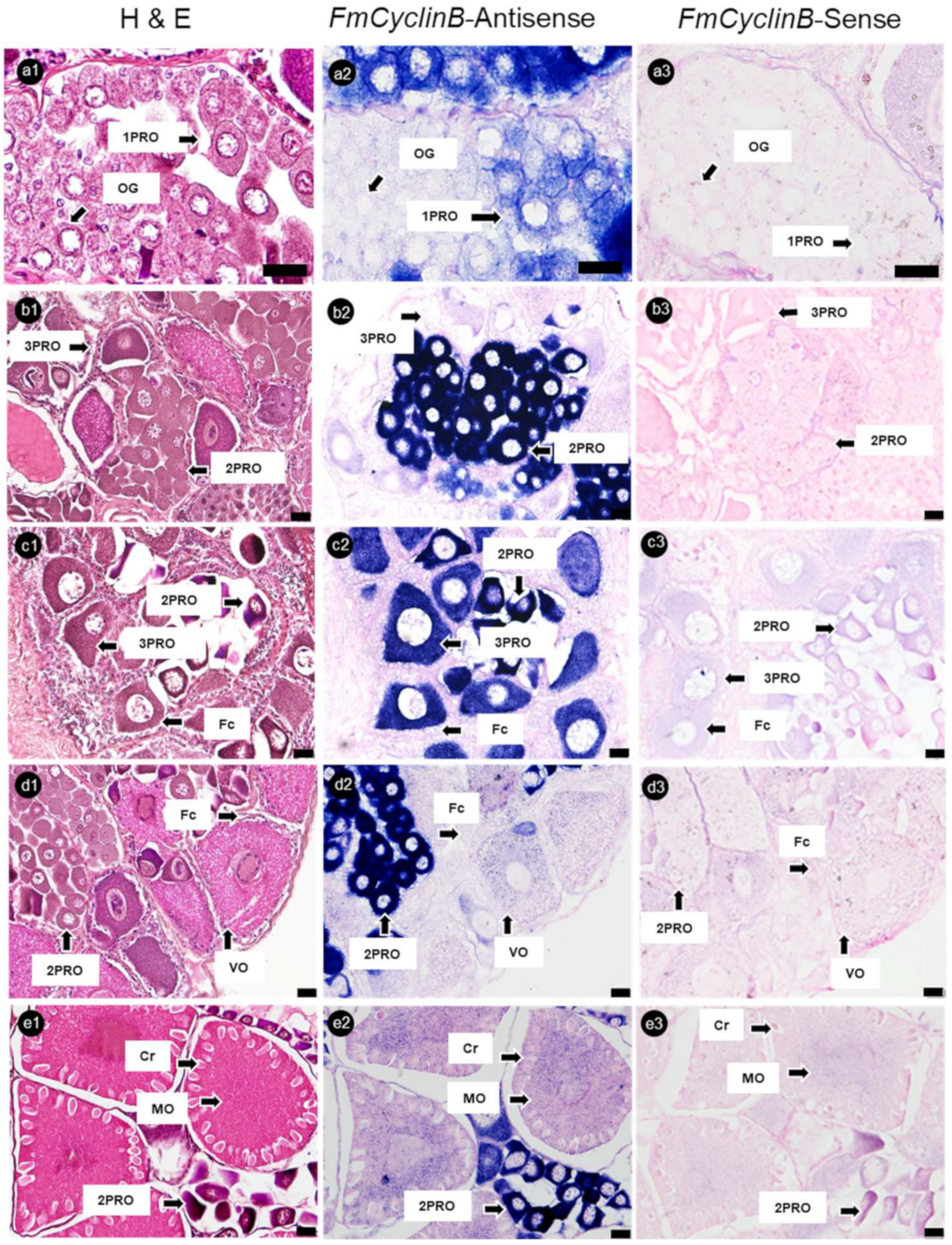
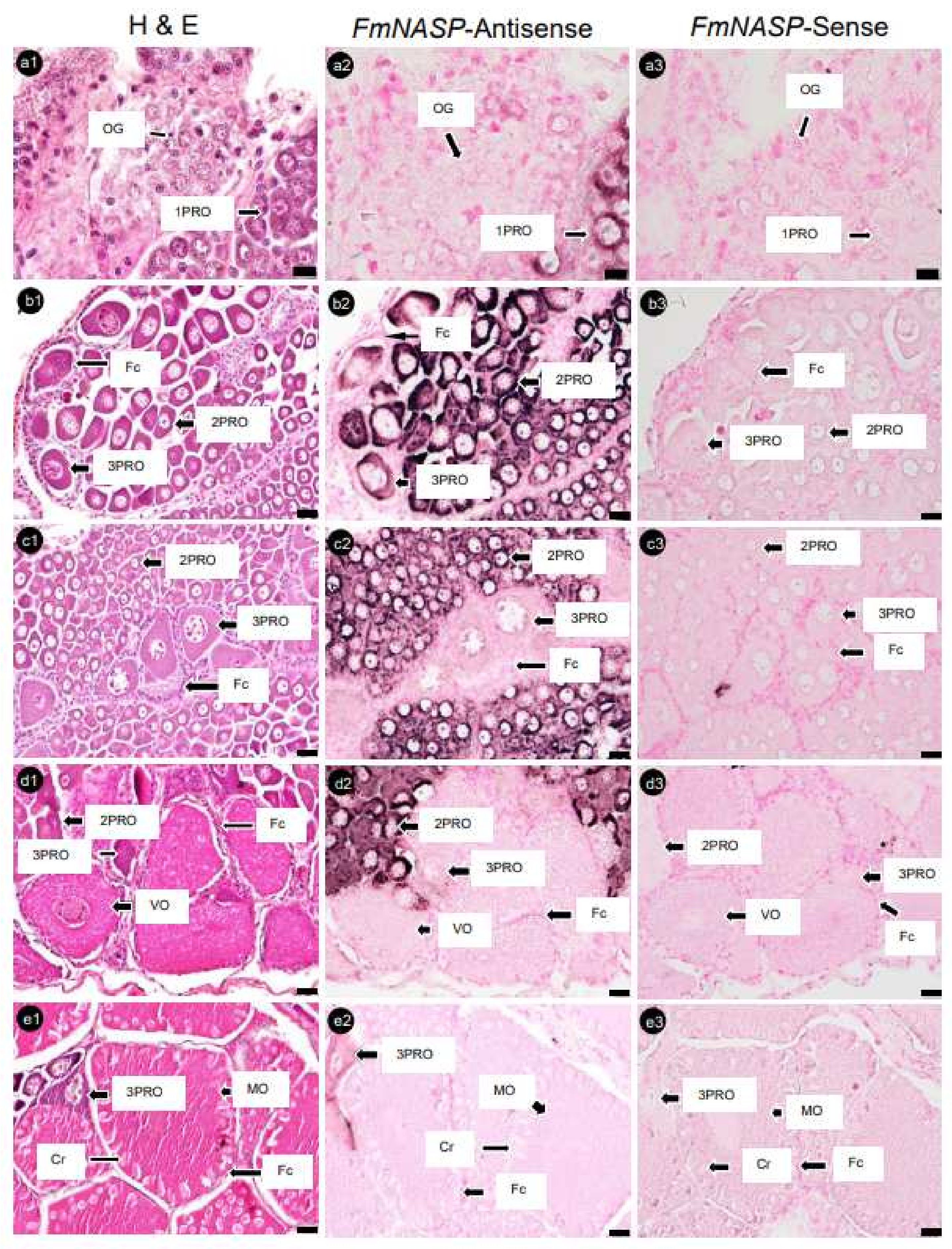
3.4. Localization of mRNA-Positive Cells in Ovaries of Banana Prawn via ISH Analysis
3.5. mRNA Expression Determined via qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Penaeus merguiensis (De Man, 1888). Fisheries and Aquaculture Department. 2023. Available online: https://www.fao.org/fishery/en/aqspecies/pba (accessed on 29 June 2023).
- Boonyaratpalin, M. Nutrition of Penaeus merguiensis and Penaeus idicus. Rev. Fish. Sci. 1998, 6, 69–78. [Google Scholar] [CrossRef]
- Farhadi, A.; Shi, X.; Zhang, Y.; Zhang, Y.; Li, S.; Zheng, H.; Ikhwanuddin, M.; Ma, H. A Novel Imprinted Gene (Sp-Pol) With Sex-Specific SNP Locus and Sex-Biased Expression Pattern Provides Insights Into the Gonad Development of Mud Crab (Scylla paramamosain). Front. Mar. Sci. 2021, 8, 727607. [Google Scholar] [CrossRef]
- Farhadi, A.; Cui, W.; Zheng, H.; Li, S.; Zhang, Y.; Ikhwanuddin, M.; Ma, H. The Regulatory Mechanism of Sexual Development in Decapod Crustaceans. Front. Mar. Sci. 2021, 8, 679687. [Google Scholar] [CrossRef]
- Farhadi, A.; Fang, S.; Zhang, Y.; Cui, W.; Fang, H.; Ikhwanuddin, M.; Ma, H. The significant sex-biased expression pattern of Sp-Wnt4 provides novel insights into the ovarian development of mud crab (Scylla paramamosain). Int. J. Biol. Macromol. 2021, 183, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, A. Sex determination and developmental mechanism of crustaceans and shellfish, volume II. Front. Endocrinol. 2023, 14, 453. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Hua, S.; Li, Y.; Zhang, S.; Liu, X.; Shi, T.; Wang, P.; Yan, B.; Li, J.; Gao, H. Expression of the cyclin-dependent kinase 2 gene (cdk2) influences ovarian development in the ridgetail white prawn, Exopalaemon carinicauda. Aquac. Rep. 2022, 25, 101265. [Google Scholar] [CrossRef]
- Zhou, Z.; Fu, H.; Jin, S.; Qiao, H.; Zhang, W.; Jiang, S.; Xiong, Y.; Gong, Y.; Hu, Y.; Gu, X.; et al. Function analysis and molecular characterization of cyclin A in ovary development of oriental river prawn, Macrobrachium nipponense. Gene 2021, 788, 145583. [Google Scholar] [CrossRef]
- Talakhun, W.; Phaonakrop, N.; Roytrakul, S.; Klinbunga, S.; Menasveta, P.; Khamnamtong, B. Proteomic analysis of ovarian proteins and characterization of thymosin-β and RAC-GTPase activating protein 1 of the giant tiger shrimp Penaeus monodon. Comp. Biochem. Physiol. Part D Genom. Proteom. 2014, 11, 9–19. [Google Scholar] [CrossRef]
- Visudtiphole, V.; Klinbunga, S.; Kirtikara, K. Molecular characterization and expression profiles of cyclin A and cyclin B during ovarian development of the giant tiger shrimp Penaeus monodon. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 152, 535–543. [Google Scholar] [CrossRef]
- Zhao, C.; Fu, M.; Qiu, L. Molecular cloning and functional characterization of cyclin E and CDK2 from Penaeus monodon. Genet. Mol. Res. 2016, 15, 8716. [Google Scholar] [CrossRef]
- Krausz, C.; Escamilla, A.R.; Chianese, C. Genetics of male infertility: From research to clinic. Reproduction 2015, 150, R159–R174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, F.; Zhou, Y.; Luo, X.; Fox, J.; King, M.L. Nanos1 functions as a translational repressor in the Xenopus germline. Mech. Dev. 2011, 128, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, T.; Fakim, H.; Brandmann, T.; Youn, J.-Y.; Gingras, A.-C.; Jinek, M.; Fabian, M.R. Human MARF1 is an endoribonuclease that interacts with the DCP1:2 decapping complex and degrades target mRNAs. Nucleic Acids Res. 2018, 46, 12008–12021. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, S.; Ueki, M.; Kai, T. Drosophila MARF1 ensures proper oocyte maturation by regulating nanos expression. PLoS ONE 2020, 15, e0231114. [Google Scholar] [CrossRef] [Green Version]
- Karoonuthaisiri, N.; Sittikankeaw, K.; Preechaphol, R.; Kalachikov, S.; Wongsurawat, T.; Uawisetwathana, U.; Russo, J.J.; Ju, J.; Klinbunga, S.; Kirtikara, K. ReproArrayGTS: A cDNA microarray for identification of reproduction-related genes in the giant tiger shrimp Penaeus monodon and characterization of a novel nuclear autoantigenic sperm protein (NASP) gene. Comp. Biochem. Physiol. Part D Genom. Proteom. 2009, 4, 90–99. [Google Scholar] [CrossRef]
- Yan, J.; Liu, H.; Fang, Y.; Cai, S. Molecular cloning and expression analysis of the gene encoding nuclear autoantigenic sperm protein from White Prawn (Litopenaeus vannamei). J. Fish. Sci. China 2016, 5, 1073–1079. [Google Scholar]
- Palasin, K.; Makkapan, W.; Wonglapsuwan, M.; Chotigeat, W. Effect of a recombinant ribosomal protein L10a (Rrpl10a) on Mouse Spermatogenesis. Songklanakarin J. Sci. Technol. 2019, 41, 959–965. [Google Scholar]
- Nagasawa, K.; Takeuchi, Y.; Miwa, M.; Higuchi, K.; Morita, T.; Mitsuboshi, T.; Miyaki, K.; Kadomura, K.; Yoshizaki, G. cDNA cloning and expression analysis of a vasa-like gene in Pacific bluefin tuna Thunnus orientalis. Fish. Sci. 2009, 75, 71–79. [Google Scholar] [CrossRef]
- Okutsu, T.; Kang, B.J.; Miwa, M.; Yoshizaki, G.; Maeno, Y.; Wilder, M.N. Molecular cloning and characterization of Dmc1, a gene involved in gametogenesis, from the whiteleg shrimp Litopenaeus vannamei. Fish. Sci. 2010, 76, 961–969. [Google Scholar] [CrossRef]
- Hashimoto, H.; Hara, K.; Hishiki, A.; Kawaguchi, S.; Shichijo, N.; Nakamura, K.; Unzai, S.; Tamaru, Y.; Shimizu, T.; Sato, M. Crystal structure of zinc-finger domain of Nanos and its functional implications. EMBO Rep. 2010, 11, 848–853. [Google Scholar] [CrossRef] [Green Version]
- Yano, I. Oocyte development in the kuruma prawn Penaeus japonicus. Mar. Biol. 1988, 99, 547–553. [Google Scholar] [CrossRef]
- Wang, L.; Guo, Q.; Levy, T.; Chen, T.; Wu, X. Ovarian development pattern and vitellogenesis of ridgetail white prawn, Exopalaemon carinicauda. Cell Tissue Res. 2020, 382, 367–379. [Google Scholar] [CrossRef]
- Glotzer, M.; Murray, A.W.; Kirschner, M.W. Cyclin is degraded by the ubiquitin pathway. Nature 1991, 349, 132–138. [Google Scholar] [CrossRef]
- Ito, M. Factors controlling cyclin B expression. Plant Mol. Biol. 2000, 43, 677–690. [Google Scholar] [CrossRef]
- Masui, Y.; Markert, C.L. Cytoplasmic control of nuclear behavior during meiotic maturation of frog oocytes. J. Exp. Zool. 1971, 177, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Qiu, G.-F.; Yamano, K. Three forms of cyclin B transcripts in the ovary of the kuruma prawn Marsupenaeus japonicus: Their molecular characterizations and expression profiles during oogenesis. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 141, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Kadyrova, L.Y.; Habara, Y.; Lee, T.H.; Wharton, R.P. Translational control of Maternal Cyclin B mRNA by Nanos in the Drosophila Germline. Development 2007, 134, 1519–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakahata, S.; Katsu, Y.; Mita, K.; Inoue, K.; Nagahama, Y.; Yamashita, M. Biochemical identification of Xenopus Pumilio as a sequence-specific Cyclin B1 MRNA-Binding protein that physically tnteracts with a Nanos Homolog, Xcat-2, and a Cytoplasmic Polyadenylation Element-Binding protein. J. Biol. Chem. 2001, 276, 20945–20953. [Google Scholar] [CrossRef] [Green Version]
- Qiu, G.-F.; Ramachandra, R.K.; Rexroad, C.E.; Yao, J. Molecular characterization and expression profiles of cyclin B1, B2 and Cdc2 kinase during oogenesis and spermatogenesis in rainbow trout (Oncorhynchus mykiss). Anim. Reprod. Sci. 2008, 105, 209–225. [Google Scholar] [CrossRef]
- Nishimura, Y.; Endo, T.; Kano, K.; Naito, K. Porcine Aurora A accelerates Cyclin B and Mos synthesis and promotes meiotic resumption of porcine oocytes. Anim. Reprod. Sci. 2009, 113, 114–124. [Google Scholar] [CrossRef]
- Paynton, B.V.; Bachvarova, R. Polyadenylation and deadenylation of maternal mRNAS during oocyte growth and maturation in the mouse. Mol. Reprod. Dev. 1994, 37, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, E.T.; Tansey, T.R.; Ruderman, J.V.; Gottesman, M. Sequence-specific adenylations and deadenylations accompany changes in the translation of maternal messenger RNA after fertilization of Spisula oocytes. J. Mol. Biol. 1983, 166, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Jiang, S.; Zhou, F.; Huang, J.; Guo, Y. Molecular cloning and characterization of a cyclin B gene on the ovarian maturation stage of black tiger shrimp (Penaeus monodon). Mol. Biol. Rep. 2007, 35, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Curtis, D.; Treiber, D.K.; Tao, F.; Zamore, P.D.; Williamson, J.R.; Lehmann, R. A CCHC metal-binding domain in Nanos is essential for translational regulation. EMBO J. 1997, 16, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Chen, K.; Yao, Q.; Wang, W.; Wang, Y.; Mu, R.; Chen, H.; Yang, H.; Zhou, H. The nanos gene of Bombyx mori and its expression patterns in developmental embryos and larvae tissues. Gene Expr. Patterns 2008, 8, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, R.; Nüsslein-Volhard, C. The maternal gene nanos has a central role in posterior pattern formation of the Drosophila embryo. Development 1991, 112, 679–691. [Google Scholar] [CrossRef]
- Dearden, P.K. Germ Cell Development in the Honeybee (Apis mellifera); Vasa and Nanos expression. BMC Dev. Biol. 2006, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Parisi, M.; Lin, H. Translational repression: A duet of Nanos and Pumilio. Curr. Biol. 2000, 10, R81–R83. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Name | Primer Sequence | Annealing Temp. and Time | Elongation Temp. and Time |
|---|---|---|---|---|
| Cyclin A | FmCyclinA-F | 5′-GCTGCGAAATATGAGGAGAT-3′ | 56 °C, 45 s | 72 °C, 45 s |
| FmCyclinA-R | 5′-TATGGCAGTTCAAACGCTA-3′ | |||
| Cyclin B | FmCyclinB-F | 5′-GATGTGGAGGAGGAAGC-3′ | 60 °C, 30 s | 72 °C, 30 s |
| FmCyclinB-R | 5′-AGGAAGTGCAAGGGAAGG-3′ | |||
| Cyclin E | FmCyclinE-F | 5′-CTGTGGCTATGCTCACTCCA-3′ | 60 °C, 30 s | 72 °C, 30 s |
| FmCyclinE-R | 5′-GGAGGAACCTGAATGACAAT-3′ | |||
| Nanos | FmNanos-F | 5′-ATTGCCAGAAATCACCAG-3′ | 59 °C, 30 s | 72 °C, 30 s |
| FmNanos-R | 5′-ATTCTGCCGTGTCAACAT-3′ | |||
| NASP | FmNASP-F | 5′-AAGGTTGAGGCTAAGGAG-3′ | 62 °C, 45 s | 72 °C, 45 s |
| FmNASP-R | 5′-CACAGGTTCAAGAGGCAG-3′ | |||
| MARF1 | FmMARF1-F | 5′-TGAGGCTTTTCCAGTTGCTT-3′ | 64 °C, 45 s | 72 °C, 45 s |
| FmMARF1-R | 5′-TAGGGGATAGGTGCAGTTGG-3′ | |||
| Actin | Actin-F | 5′-GCTACAGCTTCACCACCACCG-3′ | 58 °C, 30 s | 72 °C, 30 s |
| Actin-R | 5′-GATGTCCACGTCRCACTTCAT-3′ |
| Gene Name | Primer Name | Primer Sequence | Annealing Temp. and Time |
|---|---|---|---|
| Cyclin B | qFmCyclinB-F | 5′-GGAAGTGGTAGAGCATGTGGAGCA-3′ | 67 °C, 30 s |
| qFmCyclinB-R | 5′-TTGAAGCAGGGTGAAGCGGAGG-3′ | ||
| Nanos | qFmNanos-F | 5′-GGCAGTCCTCAGGATACATTTCAGC-3′ | 67 °C, 45 s |
| qFmNanos-R | 5′-GCACAGAGGGCAGACATACATTC-3′ | ||
| NASP | qFmNASP-F | 5′-TGCCGAGACCAGTGCCAGCC-3′ | 67 °C, 30 s |
| qFmNASP-R | 5′-CTCCGTTCTCCATGCGTGCCA-3′ | ||
| Actin | Actin-F | 5′-GCTACAGCTTCACCACCACCG-3′ | 54 °C, 30 s |
| Actin-R | 5′-GATGTCCACGTCRCACTTCAT-3′ |
| Gene | Nucleotide (bp) | Amino Acid (Residue) | Phosphorylation Residue | Kinase Motif |
|---|---|---|---|---|
| FmCyclinB | 2420 | 401 | S = 23, T = 7, Y = 7 | PKA, PKC, CKII, ATM, DNAPK, RSK, INSR, EGFR, cdc2 |
| FmNanos | 1486 | 426 | S = 36, T = 14, Y = 3 | PKA, PKC, PKG, CKI, CKII, ATM, DNAPK, SRC, INSR, p38MAPK, cdc2, cdk5 |
| FmNASP | 2700 | 686 | S = 44, T = 32, Y = 6 | PKA, PKC, PKG, CKI, CKII, ATM, DNAPK, EGFR, p38MAPK, cdc2, cdk5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sengseng, T.; Okutsu, T.; Songnui, A.; Boonchuay, J.; Sakunrang, C.; Wonglapsuwan, M. Molecular Markers of Ovarian Germ Cells of Banana Prawn (Fenneropenaeus merguiensis). Curr. Issues Mol. Biol. 2023, 45, 5708-5724. https://doi.org/10.3390/cimb45070360
Sengseng T, Okutsu T, Songnui A, Boonchuay J, Sakunrang C, Wonglapsuwan M. Molecular Markers of Ovarian Germ Cells of Banana Prawn (Fenneropenaeus merguiensis). Current Issues in Molecular Biology. 2023; 45(7):5708-5724. https://doi.org/10.3390/cimb45070360
Chicago/Turabian StyleSengseng, Tatiyavadee, Tomoyuki Okutsu, Anida Songnui, Jaruwan Boonchuay, Chanida Sakunrang, and Monwadee Wonglapsuwan. 2023. "Molecular Markers of Ovarian Germ Cells of Banana Prawn (Fenneropenaeus merguiensis)" Current Issues in Molecular Biology 45, no. 7: 5708-5724. https://doi.org/10.3390/cimb45070360
APA StyleSengseng, T., Okutsu, T., Songnui, A., Boonchuay, J., Sakunrang, C., & Wonglapsuwan, M. (2023). Molecular Markers of Ovarian Germ Cells of Banana Prawn (Fenneropenaeus merguiensis). Current Issues in Molecular Biology, 45(7), 5708-5724. https://doi.org/10.3390/cimb45070360