

Whole Transcriptome Analysis of Intervention Effect of Sophora subprostrate Polysaccharide on Inflammation in PCV2 Infected Murine Splenic Lymphocytes

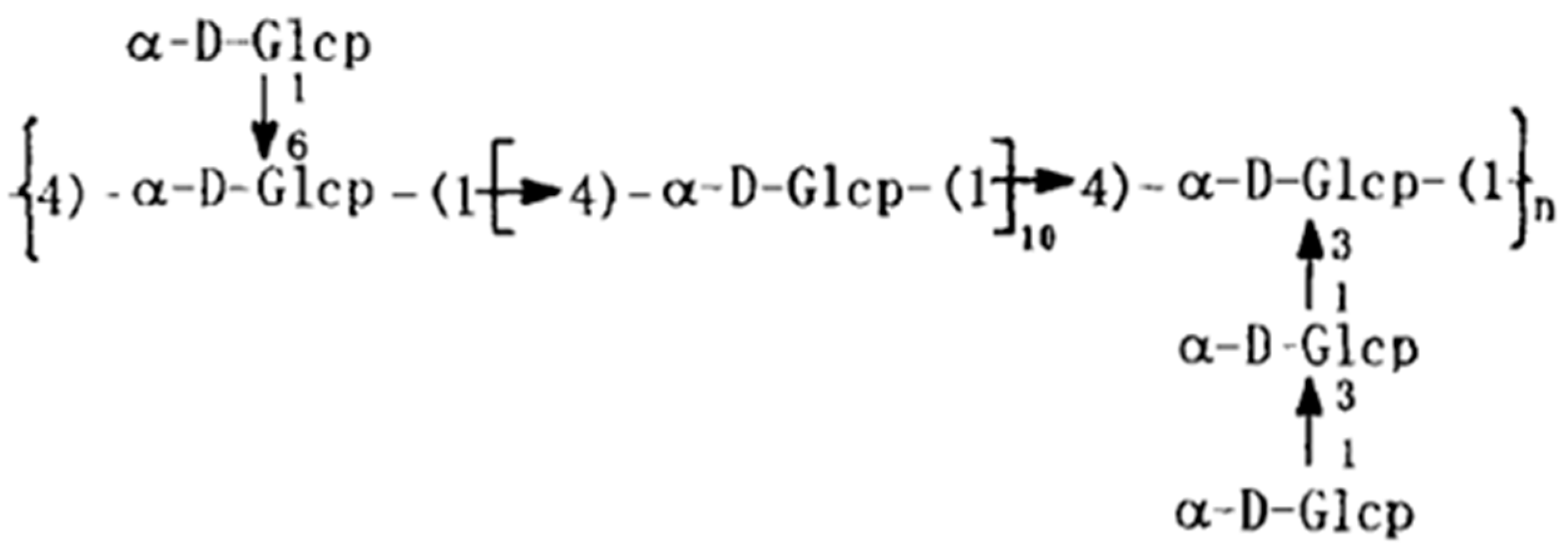{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. SSP1 and Virus
2.2. Isolation and Treatment of Murine Splenic Lymphocytes
2.3. Reagents
2.4. Cell Grouping and Treatment
2.5. RNA Sequencing and Bioinformatics Analysis
2.6. Differential Expression Analysis
2.7. Trend Analysis of mRNAs and Transcription Factor Targeting Analysis
2.8. RT- qPCR to Detect Differentially Expressed RNA
2.9. Western Blot
2.10. Target Gene Prediction and ceRNA Network Construction
2.11. The Interaction between LncRNAs, miRNAs, and mRNAs in V Group and S Group
2.12. Statistical Analysis
3. Results
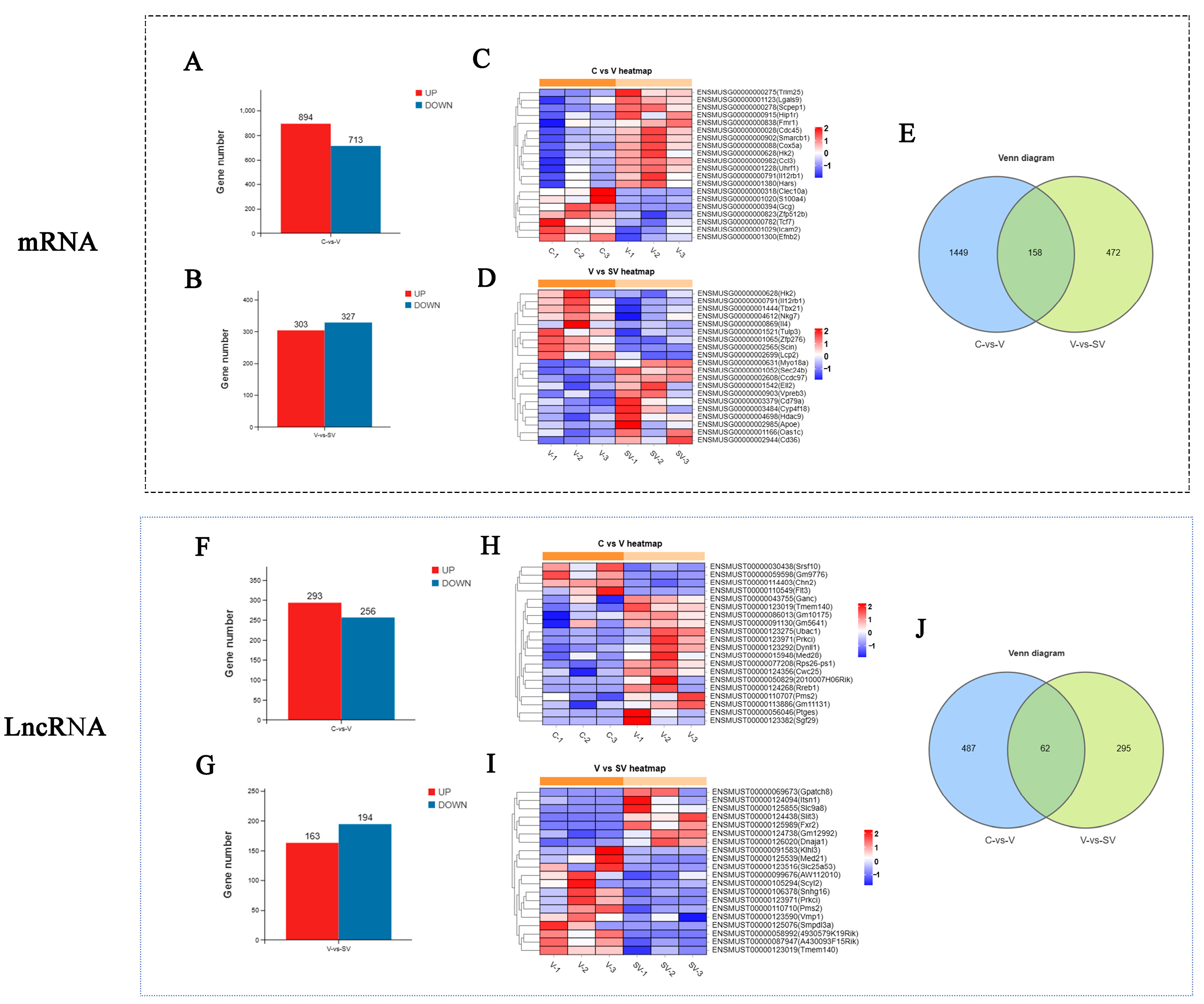
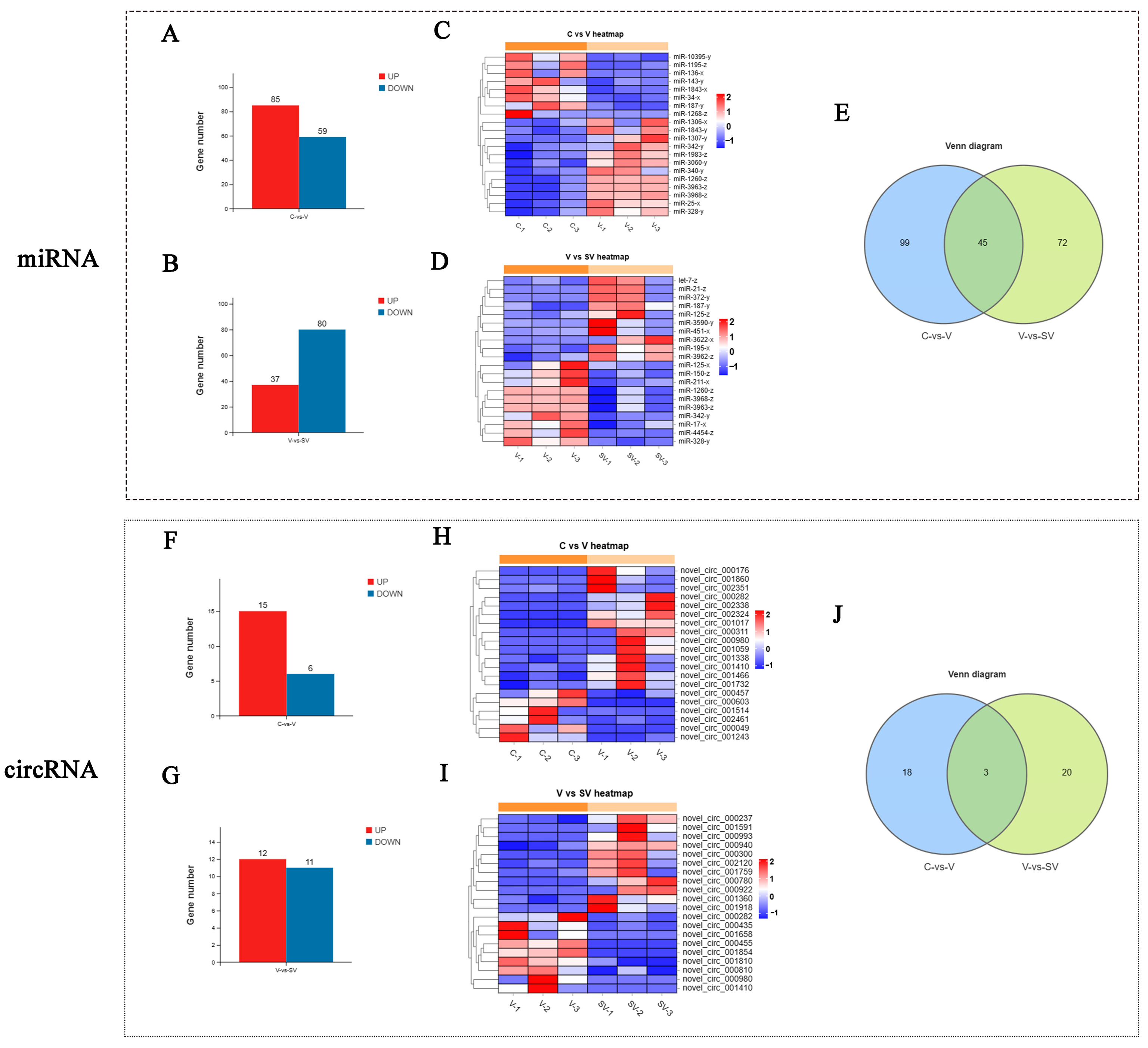
3.1. Effect of SSP1 on the mRNA, lncRNA, and miRNA Expression of the PCV2-Infected Murine Splenic Lymphocytes
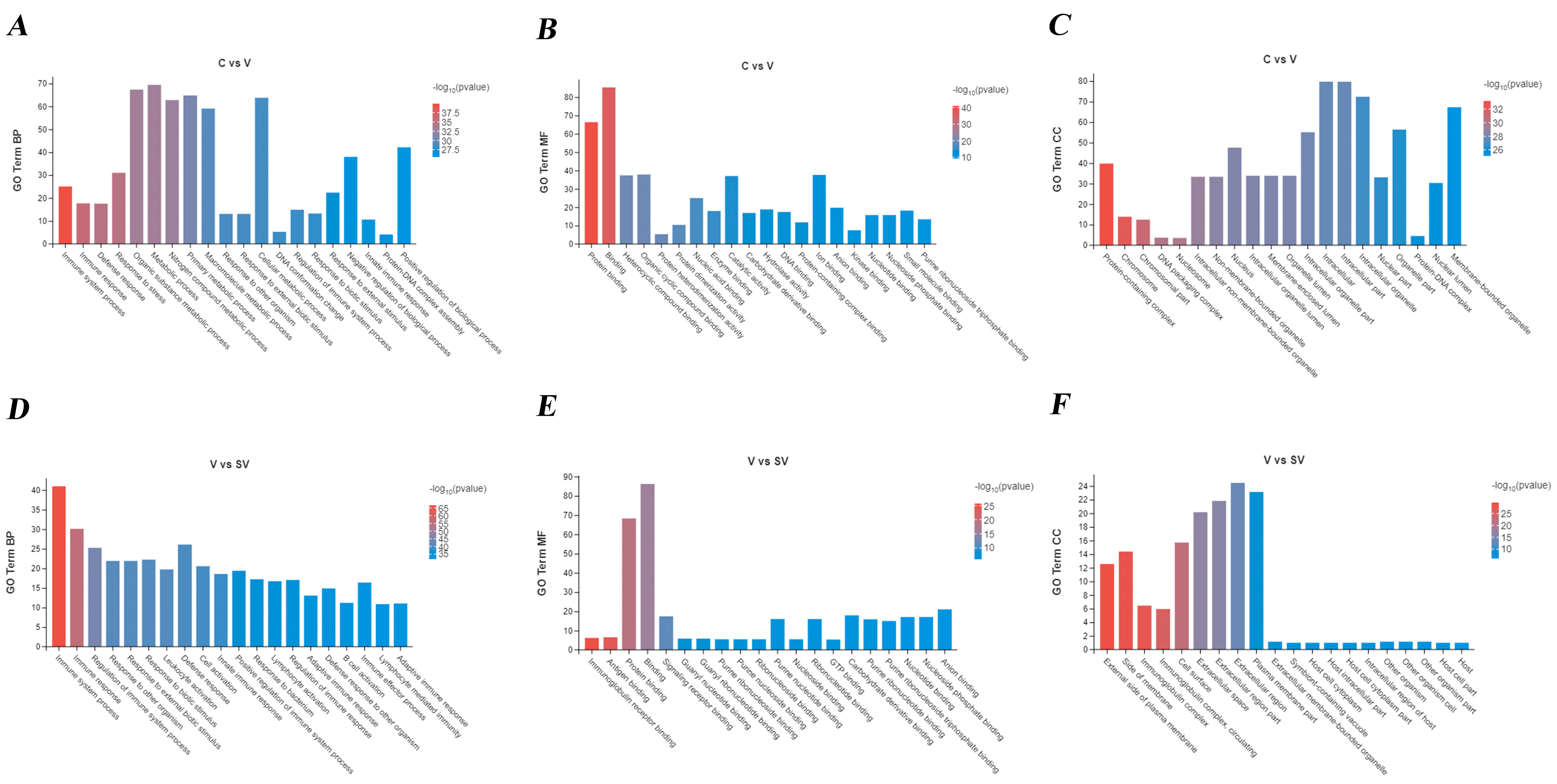
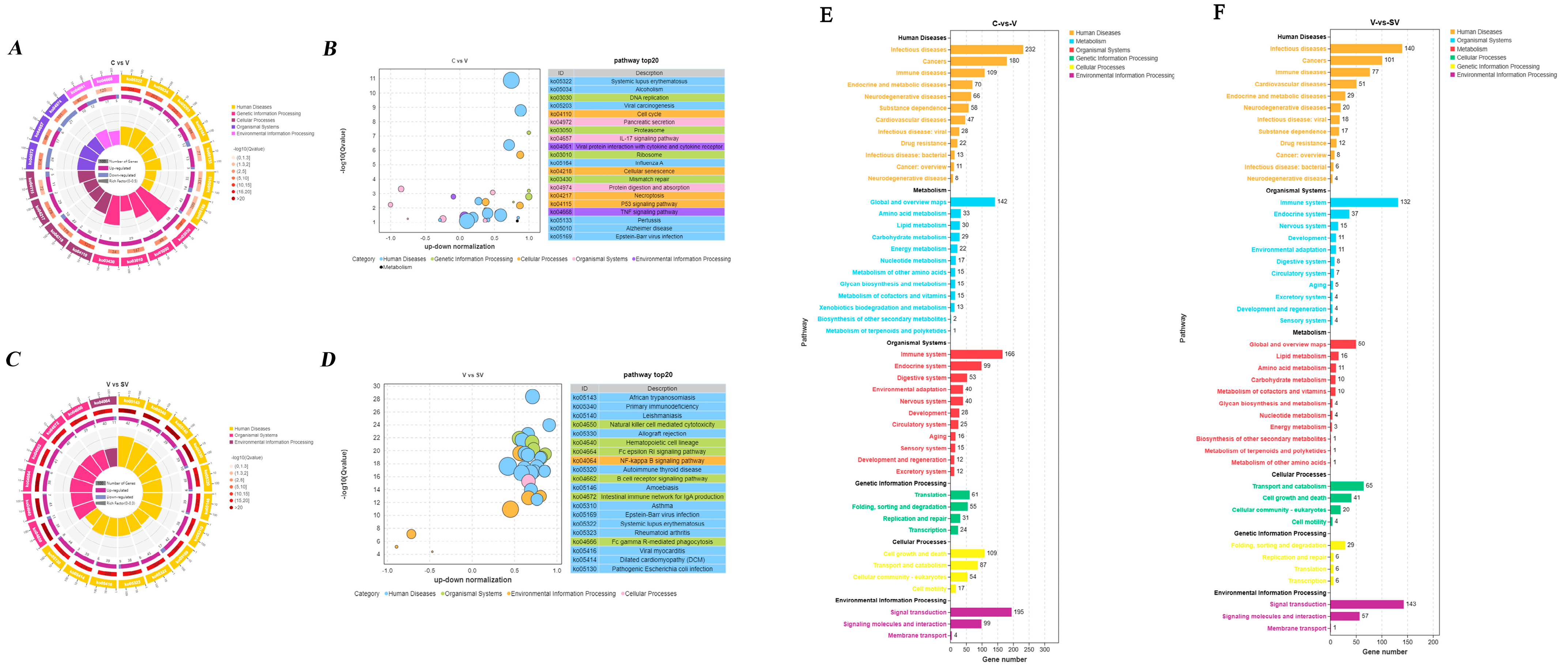
3.2. GO, KEGG Analysis of DEGs
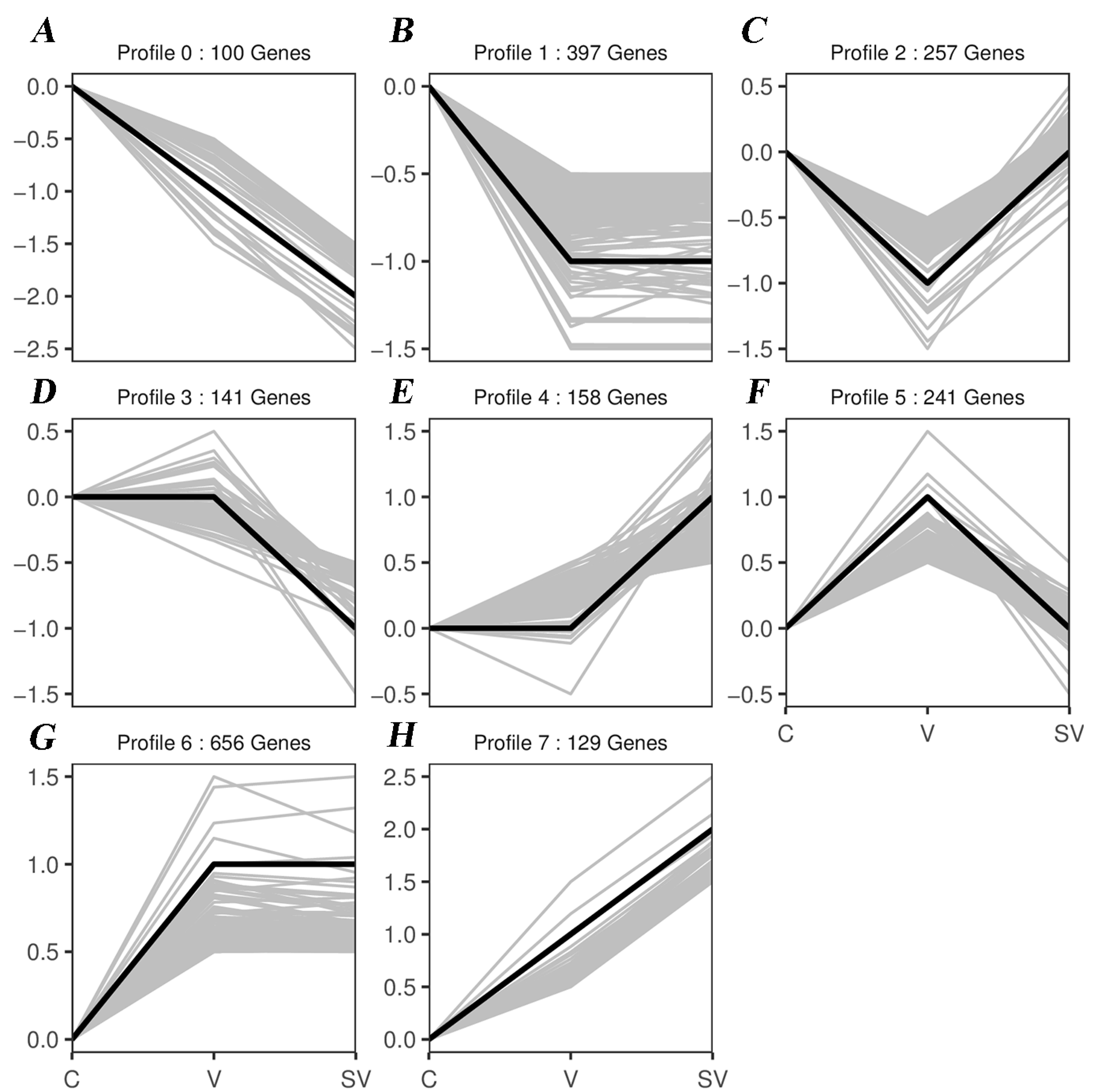
3.3. Series Test of Cluster and Transcription Factor Targeting Analysis
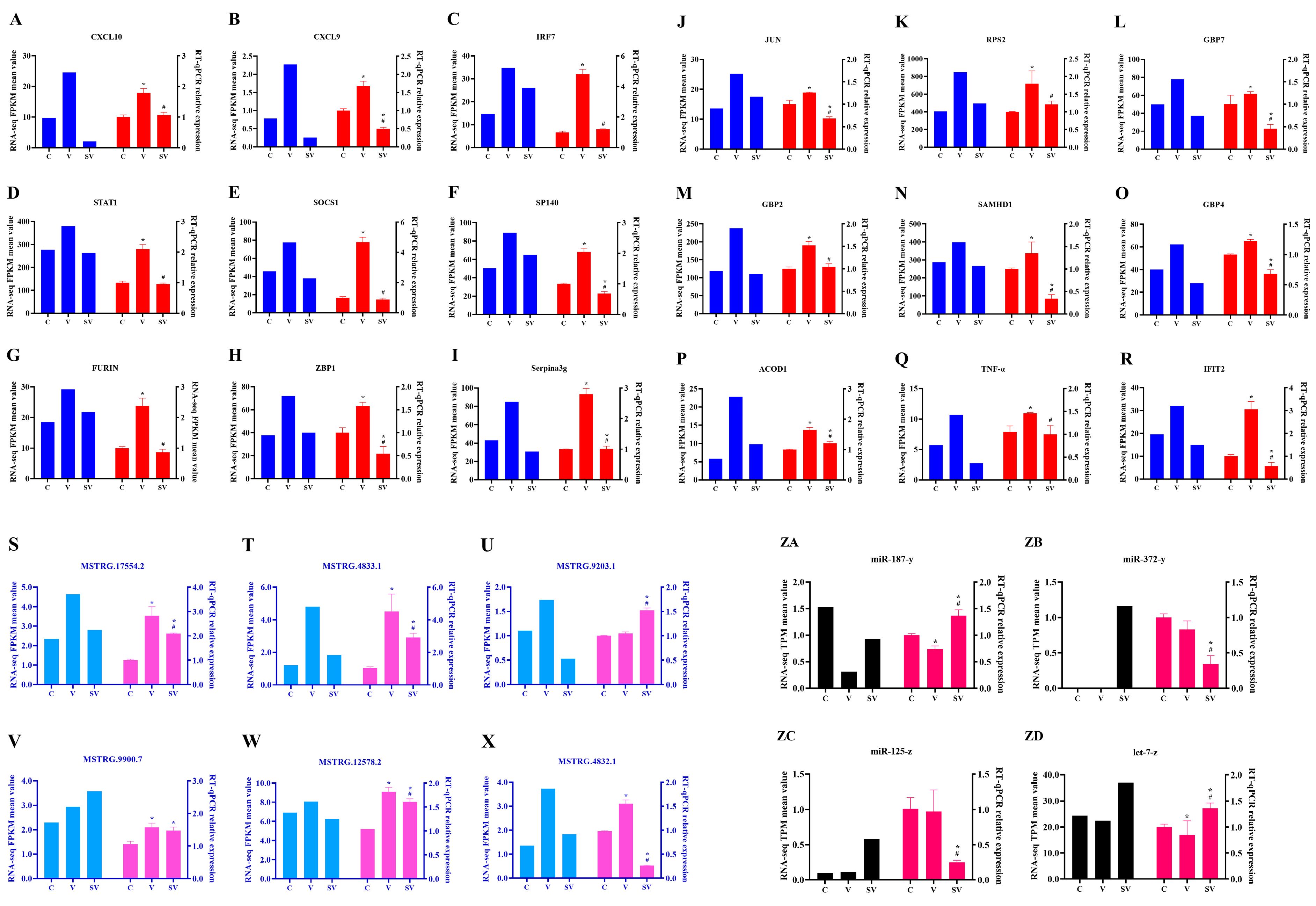
3.4. Validation of lncRNA, mRNA and miRNA
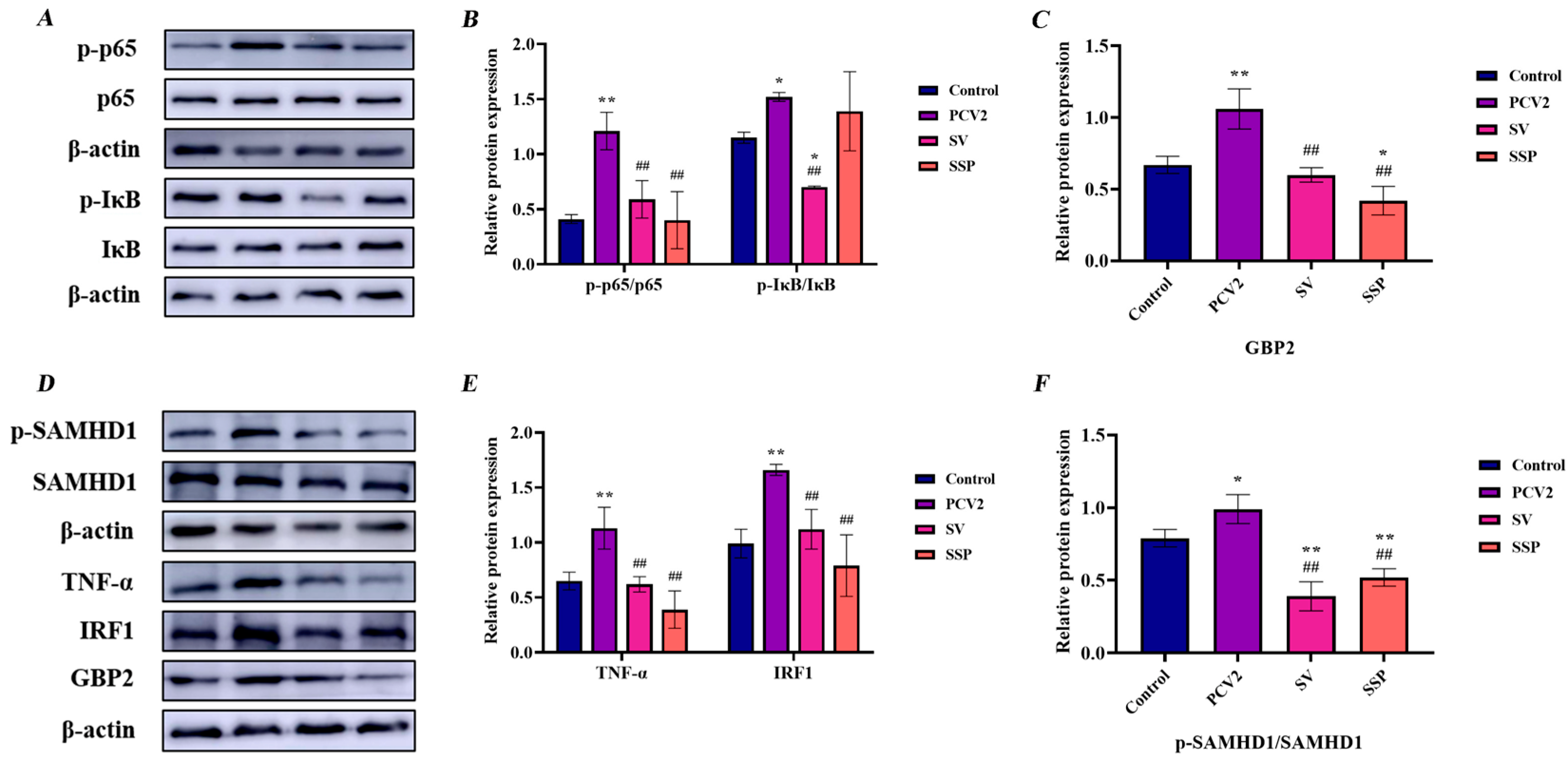
3.5. Intervention Effect of SSP1 on Inflammation-Related Protein Expression in PCV2-Infected Splenic Lymphocytes
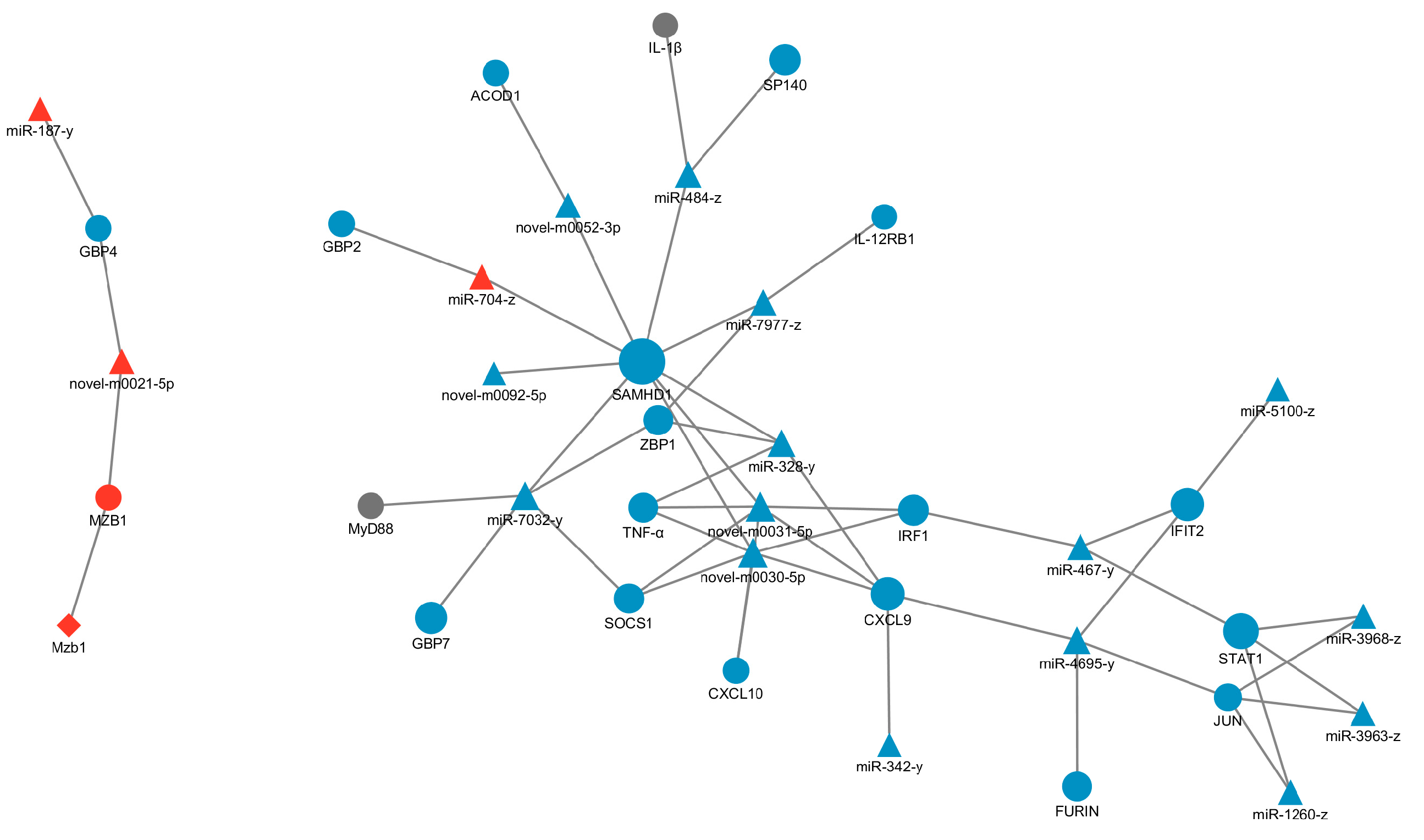
3.6. Target Gene Prediction and ceRNA Network Construction
3.7. The Interaction between LncRNAs, mRNAs and miRNAs in the V and SV Group
4. Discussion
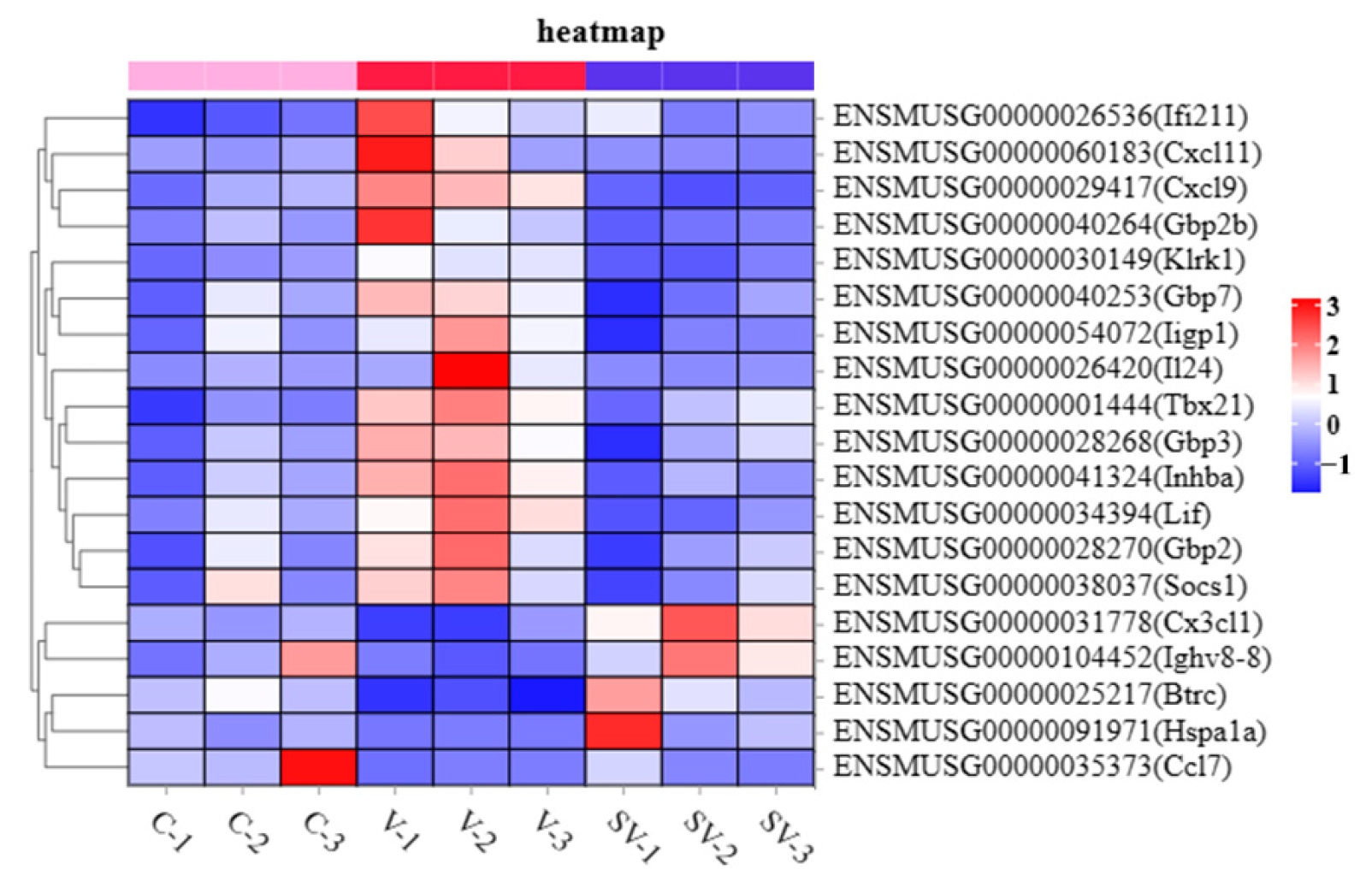
4.1. SSP1 Alleviates PCV2-Induced Inflammation in Splenic Lymphocytes by Modulating Inflammatory Gene Responses
4.2. SSP1 May Attenuate the Activation of the TNF/NF-κB Signalling Pathway through the IL12 Signalling Pathway, Thereby Reducing the Inflammatory Response
4.3. SSP1 Regulates miRNA-mRNA Networks to Modulate PCV2-Induced Inflammatory Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meng, X.J. Porcine circovirus type 2 (PCV2): Pathogenesis and interaction with the immune system. Annu. Rev. Anim. Biosci. 2013, 1, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Correa-Fiz, F.; Franzo, G.; Llorens, A.; Huerta, E.; Sibila, M.; Kekarainen, T.; Segalés, J. Porcine circovirus 2 (PCV2) population study in experimentally infected pigs developing PCV2-systemic disease or a subclinical infection. Sci. Rep. 2020, 10, 17747. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, S.; Zhang, L.Y.; Niu, G.Y.; Zhang, X.W.; Yang, L.; Ji, W.L.; Ren, L.Z. Coinfection of porcine circovirus 2 and pseudorabies virus enhances immunosuppression and inflammation through NF-κB, JAK/STAT, MAPK, and NLRP3 pathways. Int. J. Mol. Sci. 2022, 23, 4469. [Google Scholar] [CrossRef] [PubMed]
- Han, J.Y.; Zhang, S.X.; Zhang, Y.Q.; Chen, M.M.; Lv, Y.J. Porcine circovirus type 2 increases interleukin-1beta and interleukin-10 production via the MyD88-NF-kappa B signaling pathway in porcine alveolar macrophages in vitro. J. Vet. Sci. 2017, 18, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.X.; Gan, F.; Zhang, Z.Q.; Hu, J.F.; Chen, X.X.; Huang, K.H. Astragalus polysaccharides inhibits PCV2 replication by inhibiting oxidative stress and blocking NF-κB pathway. Int. J. Biol. Macromol. 2015, 81, 22–30. [Google Scholar] [CrossRef]
- Wei, L.; Zhu, S.S.; Wang, J.; Liu, J. Activation of the phosphatidylinositol 3-kinase/Akt signaling pathway during porcine circovirus type 2 infection facilitates cell survival and viral replication. J. Virol. 2012, 86, 13589–13597. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Zhang, H.; Sun, P.P.; Khan, A.; Guo, J.H.; Zheng, X.Z.; Sun, Y.G.; Fan, K.H.; Yin, W.; Li, H.Q. Matrine exhibits antiviral activity in a PRRSV/PCV2 co-infected mouse model. Phytomedicine 2020, 77, 153289. [Google Scholar] [CrossRef]
- Xue, H.X.; Gan, F.; Qian, G.; Hu, J.F.; Hao, S.; Xu, J.; Chen, X.X.; Huang, K.H. Astragalus polysaccharides attenuate PCV2 infection by inhibiting endoplasmic reticulum stress in vivo and in vitro. Sci. Rep. 2017, 7, 40440. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Cao, M.X.; Hu, W.Y.; Wei, Y.Y.; Hu, T.J. Sophora subprosrate polysaccharide suppress the inflammatory reaction of RAW264.7 cells infected with PCV2 via regulation NF-κB/MAPKs/c-Jun signal pathway and histone acetylation modification. Int. J. Biol. Macromol. 2020, 159, 957–965. [Google Scholar] [CrossRef]
- Su, Z.J.; Yang, J.; Luo, W.J.; Wei, Y.Y.; Shuai, X.H.; Hu, T.J. Inhibitory effect of Sophora subprosrate polysaccharide on mitochondria oxidative stress induced by PCV-2 infection in RAW264.7 cells. Int. J. Biol. Macromol. 2017, 95, 608–617. [Google Scholar] [CrossRef]
- Shuai, X.H.; Hu, T.J.; Liu, H.L.; Su, Z.J.; Zeng, Y.; Li, Y.H. Immunomodulatory effect of a Sophora subprosrate polysaccharide in mice. Int. J. Biol. Macromol. 2010, 46, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.X.; Yang, J.; Wang, X.R.; Hu, W.Y.; Xie, X.D.; Zhao, Y.; Liu, M.Q.; Wei, Y.Y.; Yu, M.L.; Hu, T.J. Sophora subprostrate polysaccharide regulates histone acetylation to inhibit inflammation in PCV2-infected murine splenic lymphocytes in vitro and in vivo. Int. J. Biol. Macromol. 2021, 191, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: lncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.B.; Tsitsipatis, D.; Gorospe, M. Integrated lncRNA function upon genomic and epigenomic regulation. Mol. Cell 2022, 82, 2252–2266. [Google Scholar] [CrossRef]
- Li, C.; Xia, J.R.; Yao, W.; Yang, G.; Tian, Y.Y.; Qi, Y.M.; Hao, C.F. Mechanism of LncRNA XIST/ miR-101-3p/ZEB1 axis in EMT associated with silicosis. Toxicol. Lett. 2022, 360, 11–19. [Google Scholar] [CrossRef]
- Xin, L.; Lu, H.; Liu, C.; Zeng, F.; Yuan, Y.W.; Wu, Y.; Wang, J.L.; Wu, D.Z.; Zhou, L.Q. Methionine deficiency promoted mitophagy via lncRNA PVT1-mediated promoter demethylation of BNIP3 in gastric cancer. Int. J. Biochem. Cell Biol. 2021, 141, 106100. [Google Scholar] [CrossRef]
- Navarro-Corcuera, A.; Sehrawat, T.S.; Jalan-Sakrikar, N.; Gibbons, H.R.; Pirius, N.E.; Khanal, S.; Hamdan, F.H.; Aseem, S.O.; Cao, S.; Banales, J.M.; et al. Long non-coding RNA ACTA2-AS1 promotes ductular reaction by interacting with the p300/ELK1 complex. J. Hepatol. 2022, 76, 921–933. [Google Scholar] [CrossRef]
- Li, Y.; Miao, M.; Yin, F.; Shen, N.; Yu, W.Q.; Guo, S.D. The polysaccharide-peptide complex from mushroom Cordyceps militaris ameliorates atherosclerosis by modulating the lncRNA-miRNA-mRNA axis. Food Funct. 2022, 13, 3185–3197. [Google Scholar] [CrossRef]
- Chen, D.; Chen, T.; Guo, Y.; Wang, C.; Dong, L.; Lu, C. Platycodin D (PD) regulates LncRNA-XIST/miR-335 axis to slow down bladder cancer progression in vitro and in vivo. Exp. Cell Res. 2020, 396, 112281. [Google Scholar] [CrossRef]
- Wang, J.K.; Wang, Z.; Li, G. MicroRNA-125 in immunity and cancer. Cancer Lett. 2019, 454, 134–145. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Ferragut Cardoso, A.P.; Banerjee, M.; Nail, A.N.; Lykoudi, A.; States, J.C. miRNA dysregulation is an emerging modulator of genomic instability. Semin. Cancer Biol. 2021, 76, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.C.; Zhao, J.; Yang, Y. Downregulation of lncRNA H19 inhibits the migration and invasion of melanoma cells by inactivating the NF-κB and PI3K/Akt signaling pathways. Mol. Med. Rep. 2018, 17, 7313–7318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.Y.; Pan, J.; Zhang, N.; Wei, W.; Yu, S.S.; Ai, L.M. Knockdown of long non-coding RNA H19 inhibits multiple myeloma cell growth via NF-κB pathway. Sci. Rep. 2017, 7, 18079. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Sun, Y.M.; Li, J.; Jiang, C.S.; Zeng, W.; Zhang, H.; Fan, S.X.; He, Q.G. PCV2 regulates cellular inflammatory responses through dysregulating cellular miRNA-mRNA networks. Viruses 2019, 11, 1055. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Leng, C.L.; Pan, J.Z.; Li, A.Q.; Zhang, H.; Cong, F.; Wang, H.N. Identification of lncRNAs involved in PCV2 infection of PK-15 cells. Pathogens 2020, 9, 479. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.N.; Zhang, M.L.; Ding, Y.Q.; Hu, T.J. Effect of Sophora subprosrate polysaccharide on mRNA expression of inflammatory factor in murine splenic lymphocytes infected with PCV2. Mod. J. Anim. Husb. Vet. Med. 2022, 403, 1–5. [Google Scholar]
- Montuori, E.; Martinez, K.A.; De Luca, D.; Ianora, A.; Lauritano, C. Transcriptome sequencing of the diatom asterionellopsis thurstonii and in silico identification of enzymes potentially involved in the synthesis of bioactive molecules. Mar. Drugs 2023, 21, 126. [Google Scholar] [CrossRef]
- Cai, L.L.; Zou, S.S.; Liang, D.P.; Luan, L.B. Structural characterization, antioxidant and hepatoprotective activities of polysaccharides from Sophorae tonkinensis Radix. Carbohydr. Polym. 2018, 184, 354–365. [Google Scholar] [CrossRef]
- Mariappan, V.; Adikari, S.; Shanmugam, L.; Easow, J.M.; Balakrishna Pillai, A. Differential expression of interferon inducible protein: Guanylate binding protein (GBP1 & GBP2) in severe dengue. Free. Radic. Biol. Med. 2023, 194, 131–146. [Google Scholar]
- Sze, A.; Olagnier, D.; Lin, R.; van Grevenynghe, J.; Hiscott, J. SAMHD1 host restriction factor: A link with innate immune sensing of retrovirus infection. J. Mol. Biol. 2013, 425, 4981–4994. [Google Scholar] [CrossRef] [PubMed]
- Srinivasachar Badarinarayan, S.; Shcherbakova, I.; Langer, S.; Koepke, L.; Preising, A.; Hotter, D.; Kirchhoff, F.; Sparrer, K.M.J.; Schotta, G.; Sauter, D. HIV-1 infection activates endogenous retroviral promoters regulating antiviral gene expression. Nucleic Acids Res. 2020, 48, 10890–10908. [Google Scholar] [CrossRef] [PubMed]
- Tolomeo, M.; Cavalli, A.; Cascio, A. Stat1 and its crucial role in the control of viral infections. Int. J. Mol. Sci. 2022, 23, 4095. [Google Scholar] [CrossRef]
- Wu, M.; Skaug, B.; Bi, X.; Mills, T.; Salazar, G.; Zhou, X.; Reveille, J.; Agarwal, S.K.; Blackburn, M.R.; Mayes, M.D.; et al. Interferon regulatory factor 7 (IRF7) represents a link between inflammation and fibrosis in the pathogenesis of systemic sclerosis. Ann. Rheum. Dis. 2019, 78, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Y.H.; Wang, S.; Zhang, Y.; Huang, T.; Cai, Y.D. Prediction and analysis of essential genes using the enrichments of gene ontology and KEGG pathways. PLoS ONE 2017, 12, e0184129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Xi, S. Bioinformatics analysis of differentially expressed genes and pathways in the development of cervical cancer. BMC Cancer 2021, 21, 733. [Google Scholar] [CrossRef]
- Wu, L.; Du, L.; Ju, Q.; Chen, Z.; Ma, Y.; Bai, T.; Ji, G.; Wu, Y.; Liu, Z.; Shao, Y.; et al. Silencing TLR4/MyD88/NF-κB signaling pathway alleviated inflammation of corneal epithelial cells infected by ISE. Inflammation 2021, 44, 633–644. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, Z.; Fu, T.; Ji, J.; Yang, J.; Gu, Z. TNF-α regulates microglial activation via the NF-κB signaling pathway in systemic lupus erythematosus with depression. Int. J. Biol. Macromol. 2019, 125, 892–900. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of TNF-α and IFN-γ Triggers Inflammatory Cell Death, Tissue Damage, and Mortality in SARS-CoV-2 Infection and Cytokine Shock Syndromes. Cell 2021, 184, 149–168. [Google Scholar] [CrossRef]
- Chu, Y.B.; Li, J.; Jia, P.; Cui, J.; Zhang, R.; Kang, X.; Lv, M.; Zhang, S. Irf1- and Egr1-activated transcription plays a key role in macrophage polarization: A multiomics sequencing study with partial validation. Int. Immunopharmacol. 2021, 99, 108072. [Google Scholar] [CrossRef]
- Issaranggun Na Ayuthaya, B.; Everts, V.; Pavasant, P. Interleukin-12 induces receptor activator of nuclear factor-kappa B ligand expression by human periodontal ligament cells. J. Periodontol. 2017, 88, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Ito, T.; Oyama, T.; Kitagawa, T.; Yamori, T.; Fujiwara, H.; Matsuda, H. The direct effect of IL-12 on tumor cells: IL-12 acts directly on tumor cells to activate NF-kappaB and enhance IFN-gamma-mediated STAT1 phosphorylation. Biochem. Biophys. Res. Commun. 2001, 280, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.Z.; Liu, J.; Wang, G.Q.; Song, Z.F. miR-484: A Potential Biomarker in Health and Disease. Front. Oncol. 2022, 12, 830420. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.M. MiR-484 suppressed proliferation, migration, invasion and induced apoptosis of gastric cancer via targeting CCL-18. Int. J. Exp. Pathol. 2020, 101, 203–214. [Google Scholar] [CrossRef]
- Xu, M.; Li, X.Y.; Song, L.C.; Tao, C.; Fang, J.H.; Tao, L. miR-484 targeting of Yap1-induced LPS-inhibited proliferation, and promoted apoptosis and inflammation in cardiomyocyte. Biosci. Biotechnol. Biochem. 2021, 85, 378–385. [Google Scholar] [CrossRef]
- Yang, J.S.; Zhang, M.; Yang, D.W.; Ma, Y.F.; Tang, Y.T.; Xing, M.Y.; Li, L.Y.; Chen, L.; Jin, Y.C.; Ma, C.Y. m6A-mediated upregulation of AC008 promotes osteoarthritis progression through the miR-328-3p–AQP1/ANKH axis. Exp. Mol. Med. 2021, 53, 1723–1734. [Google Scholar] [CrossRef]
- Pan, S.; Ren, F.; Li, L.; Liu, D.H.; Li, Y.; Wang, A.M.; Li, W.H.; Dong, Y.Y.; Guo, W.J. MiR-328-3p inhibits cell proliferation and metastasis in colorectal cancer by targeting Girdin and inhibiting the PI3K/Akt signaling pathway. Exp. Cell Res. 2020, 390, 111939. [Google Scholar] [CrossRef]
- Mendonça, L.S.O.; Santos, J.M.; Kaneto, C.M.; De Carvalho, L.D.; Lima-Santos, J.; Augusto, D.G.; Carvalho, S.M.S.; Soares-Martins, J.A.P.; Silva-Jardim, I. Characterization of serum cytokines and circulating microRNAs that are predicted to regulate inflammasome genes in cutaneous leishmaniasis patients. Exp. Parasitol. 2020, 210, 107846. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Jia, N.; Xie, X.; Chen, Q.; Hu, T. Whole Transcriptome Analysis of Intervention Effect of Sophora subprostrate Polysaccharide on Inflammation in PCV2 Infected Murine Splenic Lymphocytes. Curr. Issues Mol. Biol. 2023, 45, 6067-6084. https://doi.org/10.3390/cimb45070383
Zhao Y, Jia N, Xie X, Chen Q, Hu T. Whole Transcriptome Analysis of Intervention Effect of Sophora subprostrate Polysaccharide on Inflammation in PCV2 Infected Murine Splenic Lymphocytes. Current Issues in Molecular Biology. 2023; 45(7):6067-6084. https://doi.org/10.3390/cimb45070383
Chicago/Turabian StyleZhao, Yi, Nina Jia, Xiaodong Xie, Qi Chen, and Tingjun Hu. 2023. "Whole Transcriptome Analysis of Intervention Effect of Sophora subprostrate Polysaccharide on Inflammation in PCV2 Infected Murine Splenic Lymphocytes" Current Issues in Molecular Biology 45, no. 7: 6067-6084. https://doi.org/10.3390/cimb45070383
APA StyleZhao, Y., Jia, N., Xie, X., Chen, Q., & Hu, T. (2023). Whole Transcriptome Analysis of Intervention Effect of Sophora subprostrate Polysaccharide on Inflammation in PCV2 Infected Murine Splenic Lymphocytes. Current Issues in Molecular Biology, 45(7), 6067-6084. https://doi.org/10.3390/cimb45070383