
Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) on Gamete Quality Parameters of Male Koi Carp (Cyprinus carpio)


 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals Used
2.2. Experimental Fish
2.3. Exposure Experiment
2.4. GSI and Histopathological Study
2.5. Quantification of DEHP
2.6. Collection of Semen
2.7. Evaluation of Semen Volume and pH
2.8. Evaluation of Sperm Motility
2.9. Evaluation of Spermatocrit Value
2.10. Evaluation of Sperm Density
2.11. Biochemical Analysis of Seminal Fluid
2.12. Determination of Sex Steroids
2.13. RNA Extraction and cDNA Synthesis
2.14. Quantitative Real-Time PCR
2.15. Statistical Analysis
3. Results
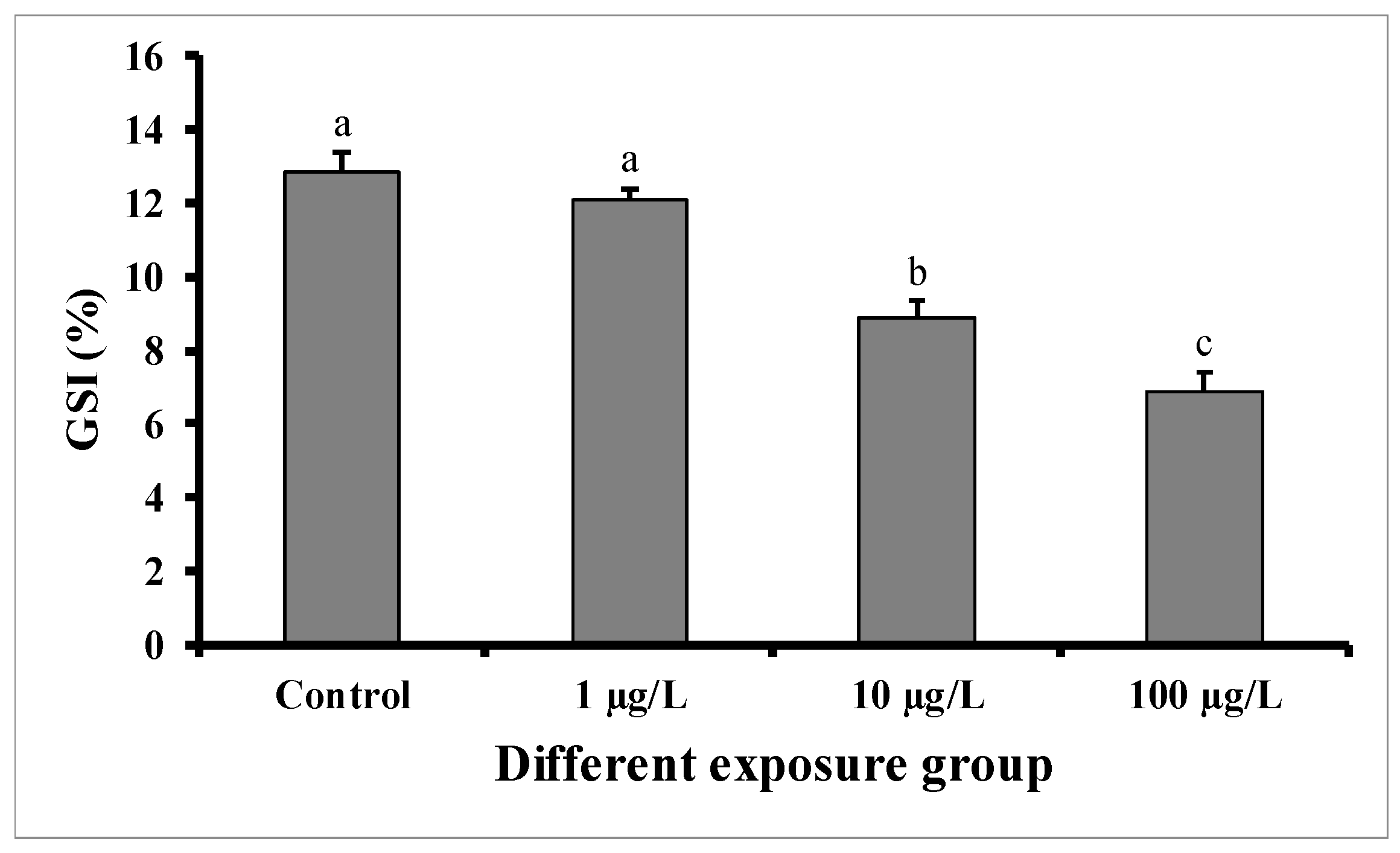
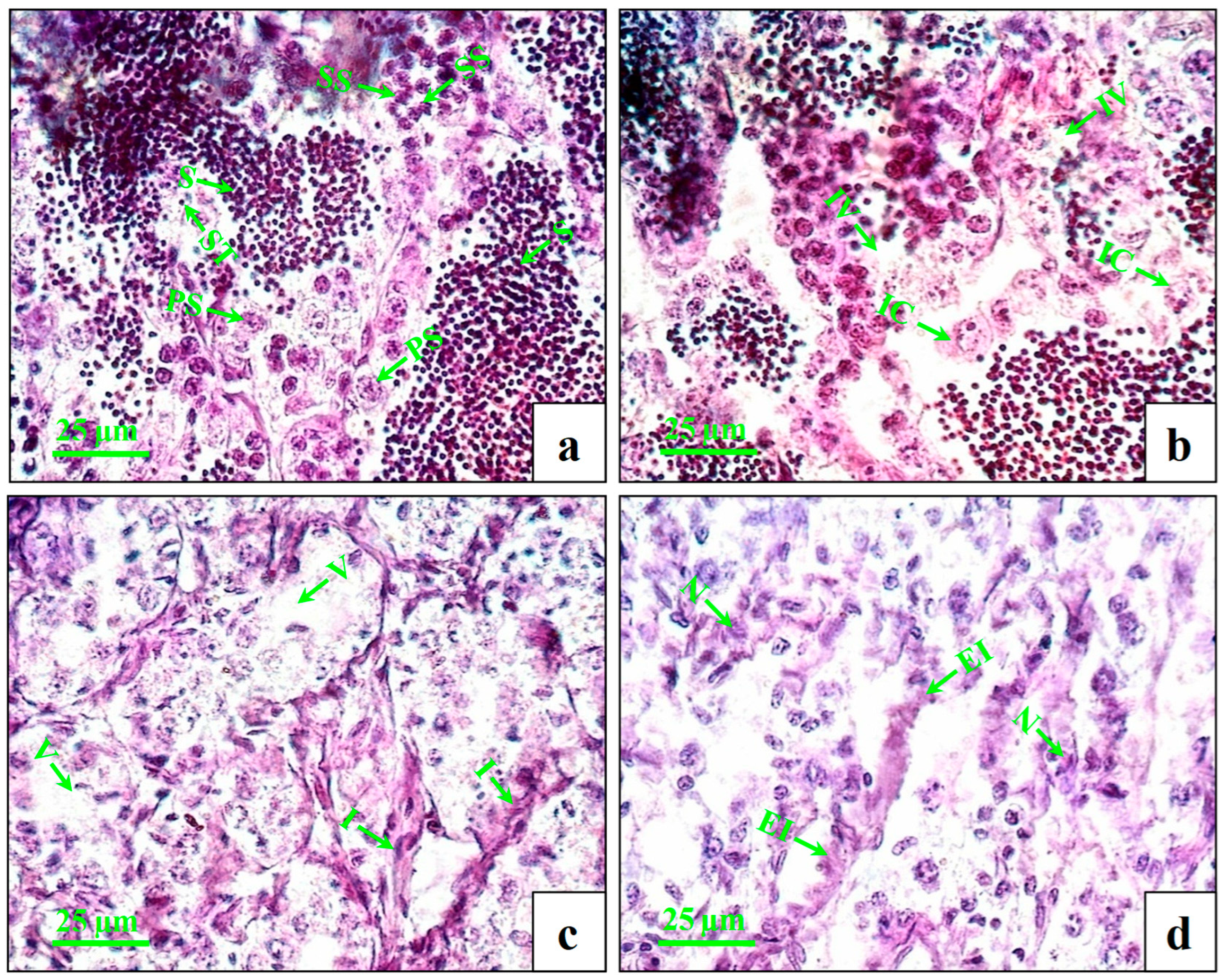
3.1. Changes in GSI and Histopathology
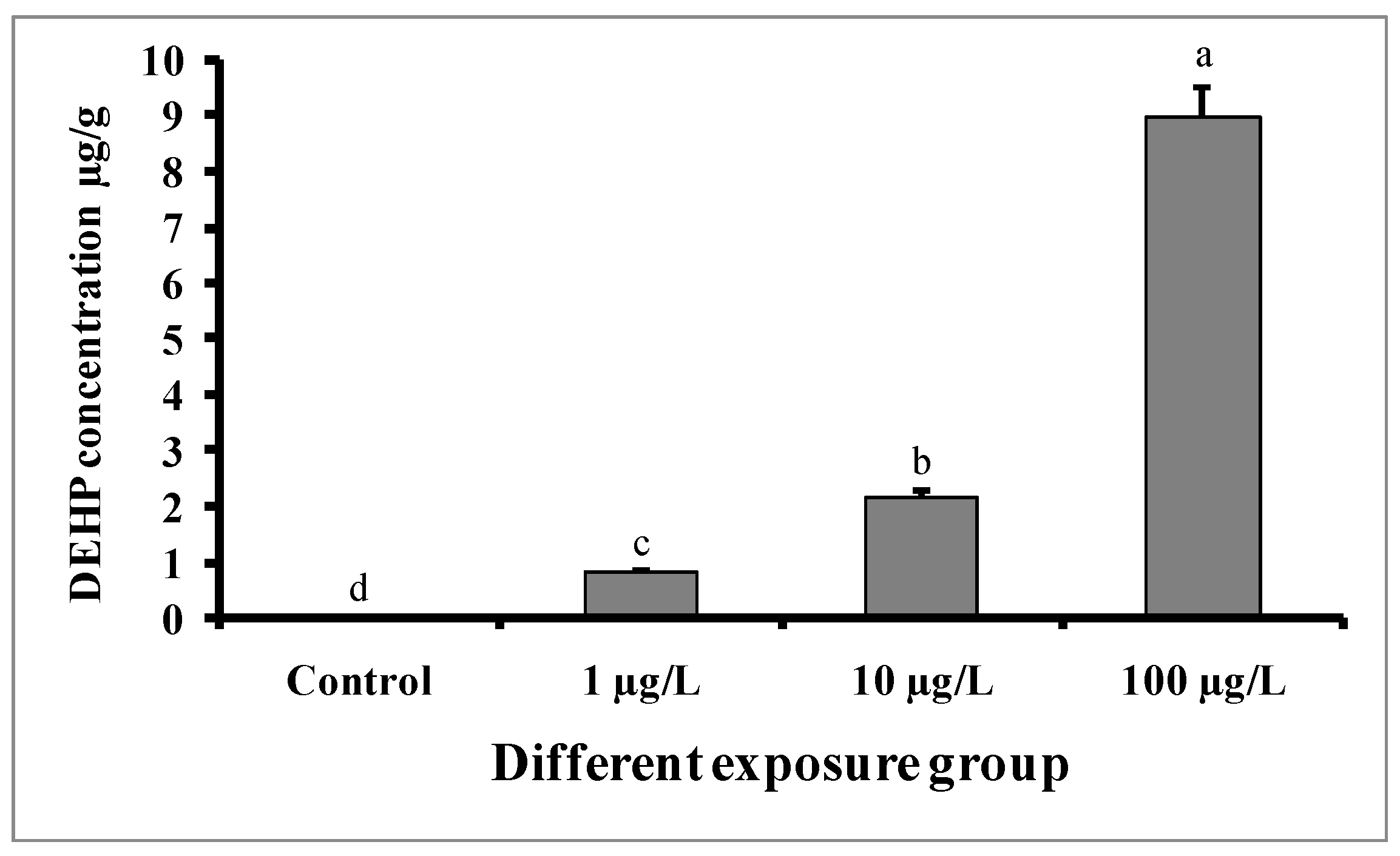
3.2. Assessment of DEHP in Tissue Samples
3.3. Changes in Semen Volume and pH
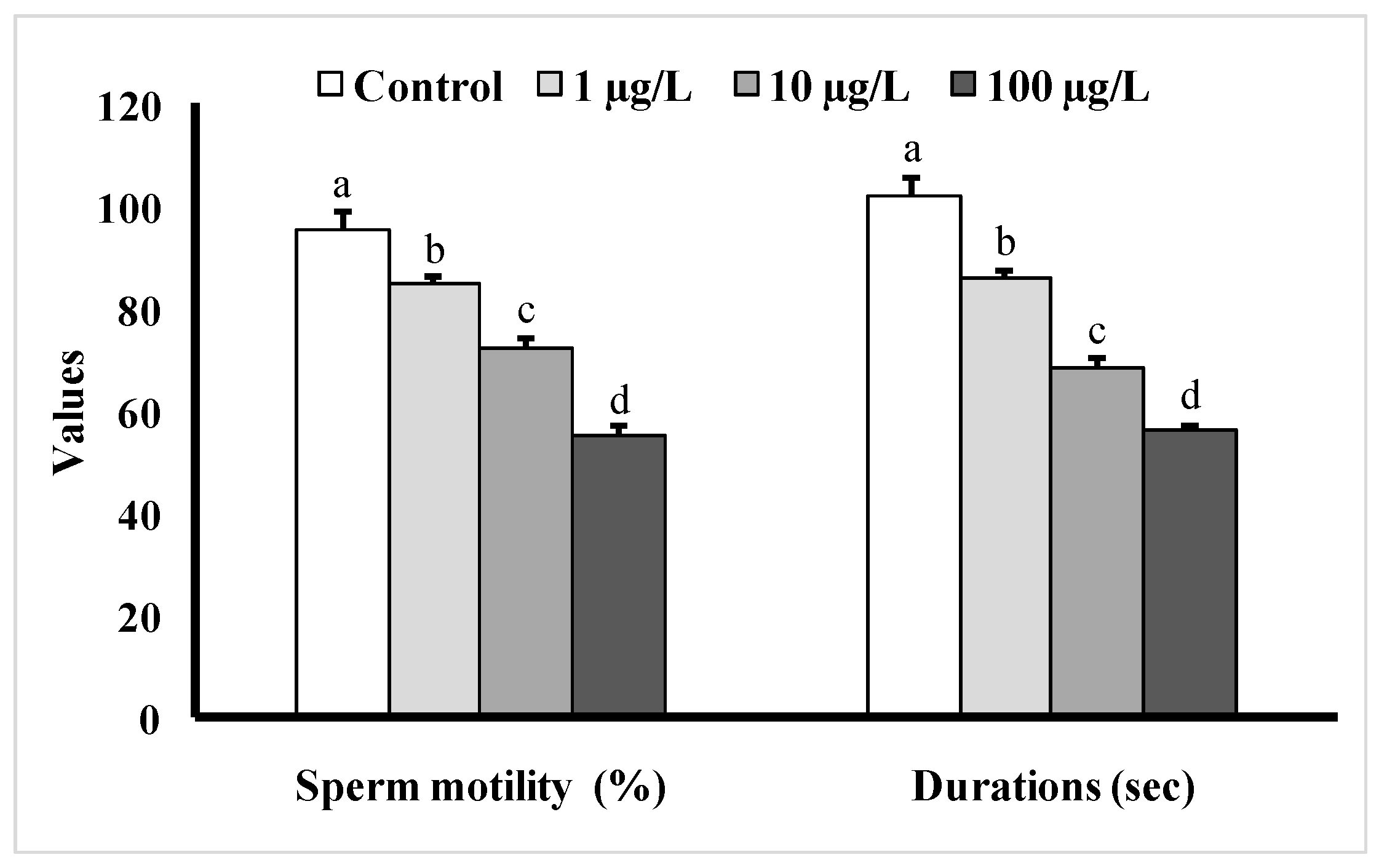
3.4. Changes in Sperm Motility
3.5. Changes in Spermatocrit Value
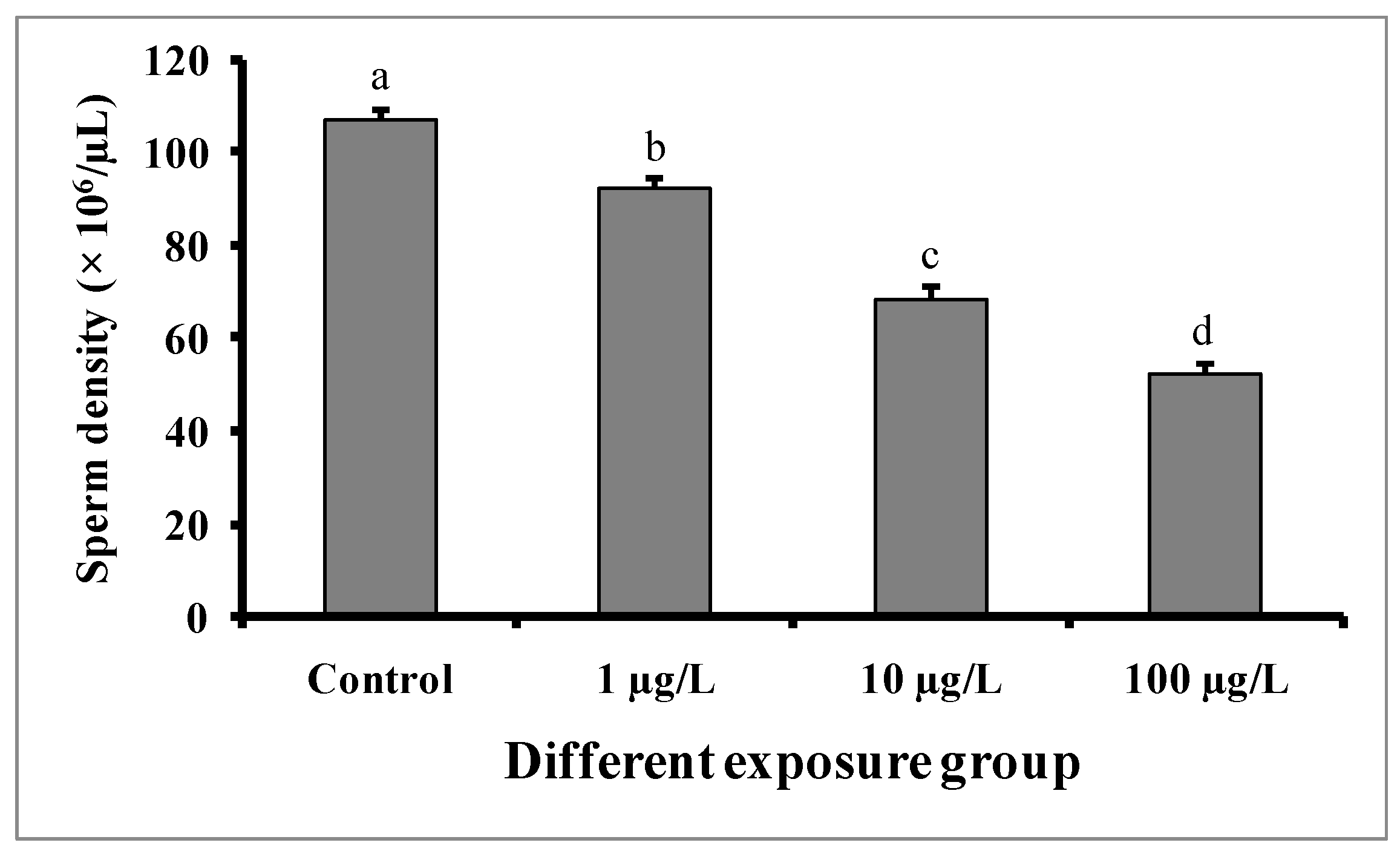
3.6. Changes in Sperm Density
3.7. Changes in Biochemical Parameters of Seminal Fluid
3.8. Sex Steroid Levels
3.9. Gene Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bold, J.; Swinburne, D. Pre-conceptual guidelines for men: A review of male infertility experience, including nutrition and lifestyle factors. Dietetics 2022, 1, 164–181. [Google Scholar] [CrossRef]
- Kumar, N.; Singh, A.K. Impact of environmental factors on human semen quality and male fertility: A narrative review. Environ. Sci. Eur. 2022, 34, 6. [Google Scholar] [CrossRef]
- Calogero, A.E.; Fiore, M.; Giacone, F.; Altomare, M.; Asero, P.; Ledda, C.; Romeo, G.; Mongioì, L.M.; Copat, C.; Giuffrida, M.; et al. Exposure to multiple metals/metalloids and human semen quality: A cross-sectional study. Ecotoxicol. Environ. Saf. 2021, 215, 112165. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, H.E.; Jørgensen, N.; Toppari, J. Semen quality in the 21st century. Nat. Rev. Urol. 2017, 14, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Montano, L.; Giorgini, E.; Notarstefano, V.; Notari, T.; Ricciardi, M.; Piscopo, M.; Motta, O. Raman Microspectroscopy evidence of microplastics in human semen. Sci. Total Environ. 2023, 901, 165922. [Google Scholar] [CrossRef]
- Yang, W.K.; Chiang, L.F.; Tan, S.W.; Chen, P.J. Environmentally relevant concentrations of di-(2-ethylhexyl) phthalate exposure alter larval growth and locomotion in medaka fish via multiple pathways. Sci. Total Environ. 2018, 640, 512–522. [Google Scholar] [CrossRef]
- Kastner, J.; Cooper, D.G.; Marić, M.; Dodd, P.; Yargeau, V. Aqueous leaching of di-(2-ethylhexyl) phthalate and “green” plasticizers from poly (vinyl chloride). Sci. Total Environ. 2012, 432, 357–364. [Google Scholar] [CrossRef]
- Magdouli, S.; Daghrir, R.; Brar, S.K.; Drogui, P.; Tyagi, R.D. Di-(2-ethylhexyl) phtalate in the aquatic and terrestrial environment: A critical review. J. Environ. Manag. 2013, 127, 36–49. [Google Scholar] [CrossRef]
- Bergé, A.; Cladière, M.; Gasperi, J.; Coursimault, A.; Tassin, B.; Moilleron, R. Meta-analysis of environmental contamination by phthalates. Environ. Sci. Pollut. Res. 2013, 20, 8057–8076. [Google Scholar] [CrossRef]
- Martine, B.; Cendrine, D.; Fabrice, A.; Marc, C. Assessment of adult human exposure to phthalate esters in the urban centre of Paris (France). Bull. Environ. Contam. Toxicol. 2013, 90, 91–96. [Google Scholar] [CrossRef]
- Schettler, T.E.D. Human exposure to phthalates via consumer products. Int. J. Androl. 2006, 29, 134–139. [Google Scholar] [CrossRef]
- Rowdhwal, S.S.S.; Chen, J. Toxic effects of di-(2-ethylhexyl) phthalate: An overview. Biomed Res. Int. 2018, 2018, 1750368. [Google Scholar] [CrossRef] [PubMed]
- Yuwatini, E.; Hata, N.; Taguchi, S. Behavior of di-(2-ethylhexyl) phthalate discharged from domestic waste water into aquatic environment. J. Environ. Monit. 2006, 8, 191–196. [Google Scholar] [CrossRef]
- Khan, M.H.; Jung, J.Y. Ozonation catalyzed by homogeneous and heterogeneous catalysts for degradation of DEHP in aqueous phase. Chemosphere 2008, 72, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Uren-Webster, T.M.; Lewis, C.; Filby, A.L.; Paull, G.C.; Santos, E.M. Mechanisms of toxicity of di-(2-ethylhexyl) phthalate on the reproductive health of male zebrafish. Aquat. Toxicol. 2010, 99, 360–369. [Google Scholar] [CrossRef] [PubMed]
- Kalo, D.; Roth, Z. Low level of mono-(2-ethylhexyl) phthalate reduces oocyte developmental competence in association with impaired gene expression. Toxicology 2017, 377, 38–48. [Google Scholar] [CrossRef]
- Bhattacharya, N.; Dufour, J.M.; Vo, M.N.; Okita, J.; Okita, R.; Kim, K.H. Differential effects of phthalates on the testes and the liver. Biol. Reprod. 2005, 72, 745–754. [Google Scholar] [CrossRef]
- Corton, J.C.; Lapinskas, P.J. Peroxisome proliferator-activated receptors: Mediators of phthalate ester-induced effects in the male reproductive tract. Toxicol. Sci. 2005, 83, 4–17. [Google Scholar] [CrossRef]
- Grande, S.W.; Andrade, A.J.; Talsness, C.E.; Grote, K.; Chahoud, I. A dose-response study following in utero and lactational exposure to di-(2-ethylhexyl) phthalate: Effects on female rat reproductive development. Toxicol. Sci. 2006, 91, 247–254. [Google Scholar] [CrossRef]
- Howdeshell, K.L.; Wilson, V.S.; Furr, J.; Lambright, C.R.; Rider, C.V.; Blystone, C.R.; Hotchkiss, A.K.; Gray, L.E., Jr. A mixture of five phthalate esters inhibits fetal testicular testosterone production in the sprague-dawley rat in a cumulative, dose-additive manner. Toxicol. Sci. 2008, 105, 153–165. [Google Scholar] [CrossRef]
- Lee, Y.J.; Lee, E.; Kim, T.H.; Choi, J.S.; Lee, J.; Jung, K.K.; Kwack, S.J.; Kim, K.B.; Kang, T.S.; Han, S.Y.; et al. Effects of di-(2-ethylhexyl) phthalate on regulation of steroidogenesis or spermatogenesis in testes of Sprague-Dawley rats. J. Health Sci. 2009, 55, 380–388. [Google Scholar] [CrossRef]
- Herr, C.; Nieden, A.Z.; Koch, H.M.; Schuppe, H.C.; Fieber, C.; Angerer, J.; Eikmann, T.; Stilianakis, N.I. Urinary di-(2-ethylhexyl) phthalate (DEHP)—Metabolites and male human markers of reproductive function. Int. J. Hyg. Environ. Health 2009, 212, 648–653. [Google Scholar] [CrossRef]
- Mendiola, J.; Jørgensen, N.; Andersson, A.M.; Calafat, A.M.; Silva, M.J.; Redmon, J.B.; Sparks, A.; Drobnis, E.Z.; Wang, C.; Liu, F.; et al. Associations between urinary metabolites of di-(2-ethylhexyl) phthalate and reproductive hormones in fertile men. Int. J. Androl. 2011, 34, 369–378. [Google Scholar] [CrossRef]
- Mendiola, J.; Meeker, J.D.; Jørgensen, N.; Andersson, A.M.; Liu, F.; Calafat, A.M.; Redmon, J.B.; Drobnis, E.Z.; Sparks, A.E.; Wang, C.; et al. Urinary concentrations of di-(2-ethylhexyl) phthalate metabolites and serum reproductive hormones: Pooled analysis of fertile and infertile men. J. Androl. 2012, 33, 488–498. [Google Scholar] [CrossRef]
- Specht, I.O.; Toft, G.; Hougaard, K.S.; Lindh, C.H.; Lenters, V.; Jönsson, B.A.; Heederik, D.; Giwercman, A.; Bonde, J.P.E. Associations between serum phthalates and biomarkers of reproductive function in 589 adult men. Environ. Int. 2014, 66, 146–156. [Google Scholar] [CrossRef]
- Wang, Y.X.; You, L.; Zeng, Q.; Sun, Y.; Huang, Y.H.; Wang, C.; Wang, P.; Cao, W.C.; Yang, P.; Li, Y.F.; et al. Phthalate exposure and human semen quality: Results from an infertility clinic in China. Environ. Res. 2015, 142, 1–9. [Google Scholar] [CrossRef]
- Corradetti, B.; Stronati, A.; Tosti, L.; Manicardi, G.; Carnevali, O.; Bizzaro, D. Bis-(2-ethylexhyl) phthalate impairs spermatogenesis in zebrafish (Danio rerio). Reprod. Biol. 2013, 13, 195–202. [Google Scholar] [CrossRef]
- Ye, T.; Kang, M.; Huang, Q.; Fang, C.; Chen, Y.; Shen, H.; Dong, S. Exposure to DEHP and MEHP from hatching to adulthood causes reproductive dysfunction and endocrine disruption in marine medaka (Oryzias melastigma). Aquat. Toxicol. 2014, 146, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Golshan, M.; Hatef, A.; Socha, M.; Milla, S.; Butts, I.A.; Carnevali, O.; Rodina, M.; Sokołowska-Mikołajczyk, M.; Fontaine, P.; Linhart, O.; et al. Di-(2-ethylhexyl)-phthalate disrupts pituitary and testicular hormonal functions to reduce sperm quality in mature goldfish. Aquat. Toxicol. 2015, 163, 16–26. [Google Scholar] [CrossRef]
- Ma, Y.B.; Jia, P.P.; Junaid, M.; Yang, L.; Lu, C.J.; Pei, D.S. Reproductive effects linked to DNA methylation in male zebrafish chronically exposed to environmentally relevant concentrations of di-(2-ethylhexyl) phthalate. Environ. Pollut. 2018, 237, 1050–1061. [Google Scholar] [CrossRef] [PubMed]
- Adeogun, A.O.; Ibor, O.R.; Imiuwa, M.E.; Omogbemi, E.D.; Chukwuka, A.V.; Omiwole, R.A.; Arukwe, A. Endocrine disruptor responses in African sharptooth catfish (Clarias gariepinus) exposed to di-(2-ethylhexyl)-phthalate. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2018, 213, 7–18. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Cosson, J. Sperm motility in fishes. (II) Effects of ions and osmolality: A review. Cell Biol. Int. 2006, 30, 1–14. [Google Scholar] [CrossRef]
- Linhart, O.; Alavi, S.M.H.; Rodina, M.; Gela, D.; Cosson, J. Comparison of sperm velocity, motility and fertilizing ability between firstly and secondly activated spermatozoa of common carp (Cyprinus carpio). J. Appl. Ichthyol. 2008, 24, 386–392. [Google Scholar] [CrossRef]
- Alavi, S.M.H.; Cosson, J.; Kazemi, R. Semen characteristics in Acipenser persicus in relation to sequential stripping. J. Appl. Ichthyol. 2006, 22, 400–405. [Google Scholar] [CrossRef]
- Rosengrave, P.; Taylor, H.; Montgomerie, R.; Metcalf, V.; Mc-Bride, K.; Gemmell, N.J. Chemical composition of seminal and ovarian fluids of chinook salmon (Oncorhynchus tshawytscha) and their effects on sperm motility traits. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2009, 152, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.K.; Yu, J.H.; Xu, P.; Li, J.L.; Li, H.X.; Ren, H.T. Identification of housekeeping genes suitable for gene expression analysis in Jian carp (Cyprinus carpio var. jian). Fish Shellfish Immunol. 2012, 33, 775–779. [Google Scholar] [CrossRef]
- Muhammad, S.; Zhang, Z.; Pavase, T.R.; Guo, H. Long-term exposure of two plasticizers di-(2-ethylhexyl)-phthalate (DEHP) and acetyl tributyl citrate (ATBC): Toxic effects on gonadal development and reproduction of zebrafish (Danio rerio). Indian J. Geo-Mar. Sci. 2018, 47, 789–797. [Google Scholar]
- Rurangwa, E.; Kime, D.E.; Ollevier, F.; Nash, J.P. The measurement of sperm motility and factors affecting sperm quality in cultured fish. Aquaculture 2004, 234, 1–28. [Google Scholar]
- Miura, T.; Miura, C.I. Molecular control mechanisms of fish spermatogenesis. Fish Physiol. Biochem. 2003, 28, 181–186. [Google Scholar] [CrossRef]
- Ismail, M.F.S.; Siraj, S.S.; Daud, S.K.; Harmin, S.A. Association of annual hormonal profile with gonad maturity of mahseer (Tor tambroides) in captivity. Gen. Comp. Endocrinol. 2011, 170, 125–130. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Oligo Sequence | Annealing Temperature | Product Size | Source |
|---|---|---|---|---|
| Fshr | F: 5′-CACACCAGATGCATTCAACC-3′ R: 5′-GTCTGCAAATGCCAGGTGGC-3′ | 58 °C | 180 bp | This study |
| Lhr | F: 5′-GCCATGCCTTCAATGGAAC-3′ R: 5′-CAGCATCAGCACAGACTCCAG-3′ | 56 °C | 172 bp | This study |
| Ar | F: 5′-GGGCCAAAGGACTTCCAGG-3′ R: 5′-GCTTCATCTGAACACAGTGC-3′ | 56 °C | 212 bp | This study |
| Erα | F: 5′-CTTTGCTCAGGATCTCATCC-3′ R: 5′-CACCATGAAGCTGTCCATC-3′ | 55 °C | 180 bp | This study |
| Erβ | F: 5′-GGAGTGCTGCTGGTTAGAGG-3′ R: 5′-CTTCAGTTCTCTGAACCTGG-3′ | 54 °C | 216 bp | This study |
| 18S rRNA | F: 5′-GAGTATGGTTGCAAAGCTGAAAC-3′ R: 5′-AATCTGTCAATCCTTTCCGTGTCC-3′ | 56 °C | 128 bp | [36] |
| Components | Control | 1 µg/L DEHP | 10 µg/L DEHP | 100 µg/L DEHP |
|---|---|---|---|---|
| Glucose (mg/dL) | 6.4 ± 0.06 d | 8.6 ± 0.08 c | 10.45 ± 0.04 b | 12.8 ± 0.08 a |
| Cholesterol (mg/dL) | 4.2 ± 0.04 c | 4.85 ± 0.08 c | 6.6 ± 0.06 b | 8.45 ± 0.06 a |
| Total protein (g/dL) | 0.86 ± 0.02 a | 0.78 ± 0.02 ab | 0.56 ± 0.01 b | 0.38 ± 0.01 c |
| Sodium (mmol/L) | 122.2 ± 2.86 a | 116.6 ± 1.88 a | 92.45 ± 1.40 b | 72.6 ± 1.98 c |
| Potassium (mmol/L) | 38.8 ± 1.24 a | 34.65 ± 0.88 ab | 30.2 ± 1.08 b | 22.4 ± 0.96 c |
| Calcium (mmol/L) | 0.66 ± 0.01 a | 0.65 ± 0.02 a | 0.64 ± 0.02 a | 0.62 ± 0.04 a |
| Magnesium (mmol/L) | 1.18 ± 0.12 a | 1.16 ± 0.08 a | 1.15 ± 0.06 a | 1.14 ± 0.04 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bisai, K.; Kumar, V.; Roy, A.; Parida, S.N.; Dhar, S.; Das, B.K.; Behera, B.K.; Pati, M.K. Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) on Gamete Quality Parameters of Male Koi Carp (Cyprinus carpio). Curr. Issues Mol. Biol. 2023, 45, 7388-7403. https://doi.org/10.3390/cimb45090467
Bisai K, Kumar V, Roy A, Parida SN, Dhar S, Das BK, Behera BK, Pati MK. Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) on Gamete Quality Parameters of Male Koi Carp (Cyprinus carpio). Current Issues in Molecular Biology. 2023; 45(9):7388-7403. https://doi.org/10.3390/cimb45090467
Chicago/Turabian StyleBisai, Kampan, Vikash Kumar, Arpita Roy, Satya Narayan Parida, Souvik Dhar, Basanta Kumar Das, Bijay Kumar Behera, and Manoj Kumar Pati. 2023. "Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) on Gamete Quality Parameters of Male Koi Carp (Cyprinus carpio)" Current Issues in Molecular Biology 45, no. 9: 7388-7403. https://doi.org/10.3390/cimb45090467
APA StyleBisai, K., Kumar, V., Roy, A., Parida, S. N., Dhar, S., Das, B. K., Behera, B. K., & Pati, M. K. (2023). Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) on Gamete Quality Parameters of Male Koi Carp (Cyprinus carpio). Current Issues in Molecular Biology, 45(9), 7388-7403. https://doi.org/10.3390/cimb45090467