
The Role of Amino Acids in the Diagnosis, Risk Assessment, and Treatment of Breast Cancer: A Review



Abstract
:1. Introduction
2. Diagnostic Value of Determination of Amino Acids in Biological Fluids
2.1. Amino Acid Composition of Serum and Blood Plasma in Breast Cancer
2.2. Features of the Amino Acid Composition of Serum/Plasma in Different Molecular Biological Subtypes of Breast Cancer
2.3. Racial Characteristics of the Serum/Plasma Amino Acid Profile in Breast Cancer
3. Amino Acid Metabolism in Breast Cancer
3.1. Metabolic Features of Breast Cancer
3.2. Amino Acid Metabolism as a Target for Breast Cancer Imaging
4. Amino Acids in Potential Strategies for the Treatment of Breast Cancer
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zeng, H.; Fan, J.; Wang, F.; Xu, C.; Li, Y.; Tu, J.; Nephew, K.P.; Long, X. Glutamine metabolism in breast cancer and possible therapeutic targets. Biochem. Pharmacol. 2023, 210, 115464. [Google Scholar] [CrossRef]
- Jiao, Z.; Pan, Y.; Chen, F. The Metabolic Landscape of Breast Cancer and Its Therapeutic Implications. Mol. Diagn. Ther. 2023, 27, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Jové, M.; Collado, R.; Quiles, J.L.; Ramírez-Tortosa, M.C.; Sol, J.; Ruiz-Sanjuan, M.; Fernandez, M.; de la Torre Cabrera, C.; Ramírez-Tortosa, C.; Granados-Principal, S.; et al. A plasma metabolomic signature discloses human breast cancer. Oncotarget 2017, 8, 19522–19533. [Google Scholar] [CrossRef]
- Jasbi, P.; Wang, D.; Cheng, S.L.; Fei, Q.; Cui, J.Y.; Liu, L.; Wei, Y.; Raftery, D.; Gu, H. Breast cancer detection using targeted plasma metabolomics. J. Chromatogr. B 2019, 1105, 26–37. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X.; Chou, J.; Yu, J.; Yang, T.; Liu, L.; Zhang, F. Taurine, glutamic acid and ethylmalonic acid as important metabolites for detecting human breast cancer based on the targeted metabolomics. Cancer Biomarkers 2018, 23, 255–268. [Google Scholar] [CrossRef]
- Shen, J.; Yan, L.; Liu, S.; Ambrosone, C.B.; Zhao, H. Plasma metabolomic profiles in breast cancer patients and healthy controls: By race and tumor receptor subtypes. Transl. Oncol. 2013, 6, 757–765. [Google Scholar] [CrossRef]
- Da Cunha, P.A.; Nitusca, D.; Canto, L.M.D.; Varghese, R.S.; Ressom, H.W.; Willey, S.; Marian, C.; Haddad, B.R. Metabolomic Analysis of Plasma from Breast Cancer Patients Using Ultra-High Performance Liquid Chromatography Coupled with Mass Spectrometry: An Untargeted Study. Metabolites 2022, 12, 447. [Google Scholar] [CrossRef]
- Neagu, A.-N.; Whitham, D.; Bruno, P.; Morrissiey, H.; Darie, C.A.; Darie, C.C. Omics-Based Investigations of Breast Cancer. Molecules 2023, 28, 4768. [Google Scholar] [CrossRef]
- Wei, Y.; Jasbi, P.; Shi, X.; Turner, C.; Hrovat, J.; Liu, L.; Rabena, Y.; Porter, P.; Gu, H. Early Breast Cancer Detection Using Untargeted and Targeted Metabolomics. J. Proteome Res. 2021, 20, 3124–3133. [Google Scholar] [CrossRef]
- Dougan, M.M.; Li, Y.; Chu, L.W.; Haile, R.W.; Whittemore, A.S.; Han, S.S.; Moore, S.C.; Sampson, J.N.; Andrulis, I.L.; John, E.M.; et al. Metabolomic profiles in breast cancer: A pilot case-control study in the breast cancer family registry. BMC Cancer 2018, 18, 532. [Google Scholar] [CrossRef]
- Budhu, A.; Terunuma, A.; Zhang, G.; Hussain, S.P.; Ambs, S.; Wang, X.W. Metabolic profiles are principally different between cancers of the liver, pancreas and breast. Int. J. Biol. Sci. 2014, 10, 966–972. [Google Scholar] [CrossRef]
- Jobard, E.; Pontoizeau, C.; Blaise, B.J.; Bachelot, T.; Elena-Herrmann, B.; Trédan, O. A serum nuclear magnetic resonance-based metabolomic signature of advanced metastatic human breast cancer. Cancer Lett. 2014, 343, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Ambs, S. Metabolic Signatures of Human Breast Cancer. Mol. Cell. Oncol. 2015, 2, e992217. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zhou, X.; Xia, T.S.; Chen, Z.; Li, J.; Liu, Q.; Alolga, R.N.; Chen, Y.; Lai, M.D.; Li, P.; et al. Human plasma metabolomics for identifying differential metabolites and predicting molecular subtypes of breast cancer. Oncotarget 2016, 7, 9925–9938. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Wang, Q.; Chen, G. Serum metabolomics analysis reveals changes in signaling lipids in breast cancer patients. Biomed. Chromatogr. 2016, 30, 42–47. [Google Scholar] [CrossRef]
- Gu, H.; Pan, Z.; Xi, B.; Asiago, V.; Musselman, B.; Raftery, D. Principal component directed partial least squares analysis for combining nuclear magnetic resonance and mass spectrometry data in metabolomics: Application to the detection of breast cancer. Anal. Chim. Acta 2011, 686, 57–63. [Google Scholar] [CrossRef]
- Oakman, C.; Tenori, L.; Claudino, W.M.; Cappadona, S.; Nepi, S.; Battaglia, A.; Bernini, P.; Zafarana, E.; Saccenti, E.; Fornier, M.; et al. Identification of a serum-detectable metabolomic fingerprint potentially correlated with the presence of micrometastatic disease in early breast cancer patients at varying risks of disease relapse by traditional prognostic methods. Ann. Oncol. 2011, 22, 1295–1301. [Google Scholar] [CrossRef]
- Tenori, L.; Oakman, C.; Claudino, W.M.; Bernini, P.; Cappadona, S.; Nepi, S.; Biganzoli, L.; Arbushites, M.C.; Luchinat, C.; Bertini, I.; et al. Exploration of serum metabolomic profiles and outcomes in women with metastatic breast cancer: A pilot study. Mol. Oncol. 2012, 6, 437–444. [Google Scholar] [CrossRef]
- Asiago, V.M.; Alvarado, L.Z.; Shanaiah, N.; Gowda, G.A.N.; Owusu-Sarfo, K.; Ballas, R.A.; Raftery, D. Early detection of recurrent breast cancer using metabolite profiling. Cancer Res. 2010, 70, 8309–8318. [Google Scholar] [CrossRef]
- Tenori, L.; Oakman, C.; Morris, P.G.; Gralka, E.; Turner, N.; Cappadona, S.; Fornier, M.; Hudis, C.; Norton, L.; Luchinat, C.; et al. Serum metabolomic profiles evaluated after surgery may identify patients with oestrogen receptor negative early breast cancer at increased risk of disease recurrence. Results from a retrospective study. Mol. Oncol. 2015, 9, 128–139. [Google Scholar] [CrossRef]
- Wei, S.; Liu, L.; Zhang, J.; Bowers, J.; Gowda, G.A.N.; Seeger, H.; Fehm, T.; Neubauer, H.J.; Vogel, U.; Clare, S.E.; et al. Metabolomics approach for predicting response to neoadjuvant chemotherapy for breast cancer. Mol. Oncol. 2013, 7, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Zapater-Moros, A.; Díaz-Beltrán, L.; Gámez-Pozo, A.; Trilla-Fuertes, L.; Lumbreras-Herrera, M.I.; López-Camacho, E.; González-Olmedo, C.; Espinosa, E.; Zamora, P.; Sánchez-Rovira, P.; et al. Metabolomics unravels subtype-specific characteristics related to neoadjuvant therapy response in breast cancer patients. Metabolomics 2023, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Torata, N.; Kubo, M.; Miura, D.; Ohuchida, K.; Mizuuchi, Y.; Fujimura, Y.; Hayakawa, E.; Kai, M.; Oda, Y.; Mizumoto, K.; et al. Visualizing energy charge in breast carcinoma tissues by MALDI mass-spectrometry imaging profiles of low-molecular-weight metabolites. Anticancer Res. 2018, 38, 4267–4272. [Google Scholar] [CrossRef] [PubMed]
- More, T.H.; RoyChoudhury, S.; Christie, J.; Taunk, K.; Mane, A.; Santra, M.K.; Chaudhury, K.; Rapole, S. Metabolomic alterations in invasive ductal carcinoma of breast: A comprehensive metabolomic study using tissue and serum samples. Oncotarget 2018, 9, 2678–2696. [Google Scholar] [CrossRef]
- Bathen, T.F.; Geurts, B.; Sitter, B.; Fjosne, H.E.; Lundgren, S.; Buydens, L.M.; Gribbestad, I.S.; Postma, G.; Giskeødegård, G.F. Feasibility of MR metabolomics for immediate analysis of resection margins during breast cancer surgery. PLoS ONE 2013, 8, e61578. [Google Scholar] [CrossRef]
- Eniu, D.T.; Romanciuc, F.; Moraru, C.; Goidescu, I.; Eniu, D.; Staicu, A.; Rachieriu, C.; Buiga, R.; Socaciu, C. The decrease of some serum free amino acids can predict breast cancer diagnosis and progression. Scand. J. Clin. Lab. Investig. 2019, 79, 17–24. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, Y.; Zhang, X. Metabolomics studies on serum and urine of patients with breast cancer using 1H-NMR spectroscopy. Oncotarget 2017, 5. [Google Scholar] [CrossRef]
- Huang, S.; Chong, N.; Lewis, N.E.; Jia, W.; Xie, G.; Garmire, L.X. Novel personalized pathway-based metabolomics models reveal key metabolic pathways for breast cancer diagnosis. Genome Med. 2016, 8, 34. [Google Scholar] [CrossRef]
- Yuan, B.W.; Schafferer, S.; Tang, Q.Q.; Scheffler, M.; Nees, J.; Heil, J.; Schott, S.; Golatta, M.; Wallwiener, M.; Sohn, C.; et al. A plasma metabolite panel as biomarkers for early primary breast cancer detection. Int. J. Cancer 2019, 144, 2833–2842. [Google Scholar] [CrossRef]
- Suman, S.; Sharma, R.K.; Kumar, V.; Sinha, N.; Shukla, Y. Metabolic fingerprinting in breast cancer stages through (1)H NMR spectroscopy-based metabolomic analysis of plasma. J. Pharm. Biomed. Anal. 2018, 160, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Cala, M.P.; Aldana, J.; Medina, J.; Sanchez, J.; Guio, J.; Wist, J.; Meesters, R.J.W. Multiplatform plasma metabolic and lipid fingerprinting of breast cancer: A pilot control-case study in Colombian Hispanic women. PLoS ONE 2018, 13, e0190958. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, Y.; Higashiyama, M.; Gochi, A.; Akaike, M.; Ishikawa, T.; Takeshi, M.; Saruki, N.; Bando, E.; Kimura, H.; Imamura, F.; et al. Plasma free amino acid profiling of five types of cancer patients and its application for early detection. PLoS ONE 2011, 6, e24143. [Google Scholar] [CrossRef] [PubMed]
- Murata, T.; Yanagisawa, T.; Kurihara, T.; Kaneko, M.; Ota, S.; Enomoto, A.; Tomita, M.; Sugimoto, M.; Sunamura, M.; Hayashida, T.; et al. Salivary metabolomics with alternative decision tree-based machine learning methods for breast cancer discrimination. Breast Cancer Res. Treat. 2019, 177, 591–601. [Google Scholar] [CrossRef]
- Zhong, L.; Cheng, F.; Lu, X.; Duan, Y.; Wang, X. Untargeted saliva metabonomics study of breast cancer based on ultra-performance liquid chromatography coupled to mass spectrometry with HILIC and RPLC separations. Talanta 2016, 158, 351–360. [Google Scholar] [CrossRef]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef]
- Bel’skaya, L.V.; Sarf, E.A.; Loginova, A.I. Diagnostic Value of Salivary Amino Acid Levels in Cancer. Metabolites 2023, 13, 950. [Google Scholar] [CrossRef]
- Cala, M.P.; Aldana, J.; Sanchez, J.; Guio, J.; Meesters, R.J.W. Urinary metabolite and lipid alterations in Colombian Hispanic women with breast cancer: A pilot study. J. Pharm. Biomed. Anal. 2018, 152, 234–241. [Google Scholar] [CrossRef]
- Slupsky, C.M.; Steed, H.; Wells, T.H.; Dabbs, K.; Schepansky, A.; Capstick, V.; Faught, W.; Sawyer, M.B. Urine metabolite analysis offers potential early diagnosis of ovarian and breast cancers. Clin. Cancer Res. 2010, 16, 5835–5841. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, R.; Song, Y.; He, J.; Sun, J.; Bai, J.; An, Z.; Dong, L.; Zhan, Q.; Abliz, Z. RRLC-MS/MS-based metabonomics combined with in-depth analysis of metabolic correlation network: Finding potential biomarkers for breast cancer. Analyst 2009, 134, 2003–2011. [Google Scholar] [CrossRef]
- Hussain, A.; Xie, L.; Deng, G.; Kang, X. Common alterations in plasma free amino acid profiles and gut microbiota-derived tryptophan metabolites of five types of cancer patients. Amino Acids 2023. [Google Scholar] [CrossRef]
- Liu, N.; Shi, F.; Yang, L.; Liao, W.; Cao, Y. Oncogenic viral infection and amino acid metabolism in cancer progression: Molecular insights and clinical implications. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2022, 1877, 188724. [Google Scholar] [CrossRef]
- Kubota, A.; Meguid, M.M.; Hitch, D.C. Amino acid profiles correlate diagnostically with organ site in three kinds of malignant tumors. Cancer 1992, 69, 2343–2348. [Google Scholar] [CrossRef]
- Cascino, A.; Muscaritoli, M.; Cangiano, C.; Conversano, L.; Laviano, A.; Ariemma, S.; Meguid, M.M.; Rossi Fanelli, F. Plasma amino acid imbalance in patients with lung and breast cancer. Anticancer Res. 1995, 15, 507–510. [Google Scholar] [PubMed]
- Proenza, A.M.A.; Oliver, J.J.; Palou, A.A.; Roca, P.P. Breast and lung cancer are associated with a decrease in blood cell amino acid content. J. Nutr. Biochem. 2003, 14, 133–138. [Google Scholar] [CrossRef]
- Minet-Quinard, R.; Van Praagh, I.; Kwiatkowski, F.; Beaujon, G.; Feillel, V.; Beaufrère, B.; Bargnoux, P.J.; Cynober, L.; Vasson, M.P. Pre- and postoperative aminoacidemia in breast cancer: A study vs. matched healthy subjects. Cancer Investig. 2004, 22, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Vissers, Y.L.J.; Dejong, C.H.C.; Luiking, Y.C.; Fearon, K.C.H.; von Meyenfeldt, M.F.; Deutz, N.E.P. Plasma arginine concentrations are reduced in cancer patients: Evidence for arginine deficiency? Am. J. Clin. Nutr. 2005, 81, 1142–1146. [Google Scholar] [CrossRef]
- Okamoto, N.; Miyagi, Y.; Chiba, A.; Akaike, M.; Shiozawa, M.; Imaizumi, A.; Yamamoto, H.; Ando, T.; Yamakado, M.; Tochikubo, O. Diagnostic modeling with differences in plasma amino acid profiles between non-cachectic colorectal/breast cancer patients and healthy individuals. Int. J. Med. Med. Sci. 2009, 1, 1–8. [Google Scholar]
- Poschke, I.; Mao, Y.; Kiessling, R.; Boniface, J. Tumor-dependent increase of serum amino acid levels in breast cancer patients has diagnostic potential and correlates with molecular tumor subtypes. J. Transl. Med. 2013, 11, 290. [Google Scholar] [CrossRef]
- Barnes, T.; Bell, K.; DiSebastiano, K.M.; Vance, V.; Hanning, R.; Russell, C.; Dubin, J.A.; Bahl, M.; Califaretti, N.; Campbell, C.; et al. Plasma amino acid profiles of breast cancer patients early in the trajectory of the disease differ from healthy comparison groups. Appl. Physiol. Nutr. Metab. 2014, 39, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Chen, T.; Fu, S.; Sun, X.; Wang, L.; Lu, Y.; Ding, S.; Ruan, G.; Teng, L.; Wang, M. Perioperative dynamics and significance of amino acid profiles in patients with cancer. J. Transl. Med. 2015, 13, 35. [Google Scholar] [CrossRef]
- Park, J.; Shin, Y.; Kim, T.H.; Kim, D.-H.; Lee, A. Plasma metabolites as possible biomarkers for diagnosis of breast cancer. PLoS ONE 2019, 14, e0225129. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, M.; Men, Y.; Wang, W.; Zhang, W. Heavy metals interfere with plasma metabolites, including lipids and amino acids, in patients with breast cancer. Oncol. Lett. 2020, 19, 2925–2933. [Google Scholar] [CrossRef]
- Politi, C.; Fattuoni, C.; Serra, A.; Noto, A.; Loi, S.; Casanova, A.; Faa, G.; Ravarino, A.; Saba, L. Metabolomic analysis of plasma from breast tumour patients. A pilot study. J. Public Health Res. 2021, 10, 2304. [Google Scholar] [CrossRef]
- An, R.; Yu, H.; Wang, Y.; Lu, J.; Gao, Y.; Xie, X.; Zhang, J. Integrative analysis of plasma metabolomics and proteomics reveals the metabolic landscape of breast cancer. Cancer Metab. 2022, 10, 13. [Google Scholar] [CrossRef]
- Baranovicova, E.; Racay, P.; Zubor, P.; Smolar, M.; Kudelova, E.; Halasova, E.; Dvorska, D.; Dankova, Z. Circulating metabolites in the early stage of breast cancer were not related to cancer stage or subtypes but associated with ki67 level. Promising statistical discrimination from controls. Mol. Cell. Probes 2022, 66, 101862. [Google Scholar] [CrossRef]
- Han, X.; Li, D.; Wang, S.; Lin, Y.; Liu, Y.; Lin, L.; Qiao, L. Serum amino acids quantification by plasmonic colloidosome-coupled MALDI-TOF MS for triple-negative breast cancer diagnosis. Mater. Today Bio 2022, 17, 100486. [Google Scholar] [CrossRef] [PubMed]
- Santaliz-Casiano, A.; Mehta, D.; Danciu, O.C.; Patel, H.; Banks, L.; Zaidi, A.; Buckley, J.; Rauscher, G.H.; Schulte, L.; Weller, L.R.; et al. Identification of metabolic pathways contributing to ER+ breast cancer disparities using a machine-learning pipeline. Sci. Rep. 2023, 13, 12136. [Google Scholar] [CrossRef] [PubMed]
- Panigoro, S.S.; Kurniawan, A.; Ramadhan, R.; Sukartini, N.; Herqutanto, H.; Paramita, R.I.; Sandra, F. Amino Acid Profile of Luminal A and B Subtypes Breast Cancer. Indones. Biomed. J. 2023, 15, 194–295. [Google Scholar] [CrossRef]
- Morad, H.M.; Abou-Elzahab, M.M.; Aref, S.; EL-Sokkary, A.M.A. Diagnostic Value of 1H NMR-Based Metabolomics in Acute Lymphoblastic Leukemia, Acute Myeloid Leukemia, and Breast Cancer. ACS Omega 2022, 7, 8128–8140. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.S.; Lee, J.C.; Lee, P.H.; Wang, S.T.; Chen, W.J. Plasma free amino acid profile in cancer patients. Semin. Cancer Biol. 2005, 15, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y.; Cai, H.; Wang, S.; Shen, Y.; Ke, C. Application of metabolomics in the diagnosis of breast cancer: A systematic review. J. Cancer 2020, 11, 2540–2551. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Gao, Y.; Cao, Y.; Zhang, Y.; Xu, M.; Wang, Y.; Jing, Y.; Guo, S.; Jing, F.; Hu, X.; et al. Identification of arginine and its “Downstream” molecules as potential markers of breast cancer. IUBMB Life 2016, 68, 817–822. [Google Scholar] [CrossRef]
- Budczies, J.; Denkert, C.; Muller, B.M.; Brockmoller, S.F.; Klauschen, F.; Gyorffy, B.; Dietel, M.; Richter-Ehrenstein, C.; Marten, U.; Salek, R.M.; et al. Remodeling of central metabolism in invasive breast cancer compared to normal breast tissue—A GC-TOFMS based metabolomics study. BMC Genom. 2012, 13, 334. [Google Scholar] [CrossRef]
- Budczies, J.; Brockmöller, S.F.; Müller, B.M.; Barupal, D.K.; Richter-Ehrenstein, C.; Kleine-Tebbe, A.; Griffin, J.L.; Orešič, M.; Dietel, M.; Denkert, C.; et al. Comparative metabolomics of estrogen receptor positive and estrogen receptor negative breast cancer: Alterations in glutamine and beta-alanine metabolism. J. Proteom. 2013, 94, 279–288. [Google Scholar] [CrossRef]
- Togashi, Y.; Arao, T.; Kato, H.; Matsumoto, K.; Terashima, M.; Hayashi, H.; de Velasco, M.A.; Fujita, Y.; Kimura, H.; Yasuda, T. Frequent amplification of ORAOV1 gene in esophageal squamous cell cancer promotes an aggressive phenotype via proline metabolism and ROS production. Oncotarget 2014, 5, 2962. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Beltrán, L.; González-Olmedo, C.; Luque-Caro, N.; Díaz, C.; Martín-Blázquez, A.; Fernández-Navarro, M.; Ortega-Granados, A.L.; Gálvez-Montosa, F.; Vicente, F.; Pérez del Palacio, J.; et al. Human Plasma Metabolomics for Biomarker Discovery: Targeting the Molecular Subtypes in Breast Cancer. Cancers 2021, 13, 147. [Google Scholar] [CrossRef] [PubMed]
- Tayyari, F.; Gowda, G.A.N.; Olopade, O.F.; Berg, R.; Yang, H.H.; Lee, M.P.; Ngwa, W.F.; Mittal, S.K.; Raftery, D.; Mohammed, S.I. Metabolic profiles of triple-negative and luminal A breast cancer subtypes in African-American identify key metabolic differences. Oncotarget 2018, 9, 11677–11690. [Google Scholar] [CrossRef]
- Chlebowski, R.T.; Chen, Z.; Anderson, G.L.; Rohan, T.; Aragaki, A.; Lane, D.; Dolan, N.C.; Paskett, E.D.; McTiernan, A.; Hubbell, F.A.; et al. Ethnicity and breast cancer: Factors influencing differences in incidence and outcome. J. Natl. Cancer Inst. 2005, 97, 439–448. [Google Scholar] [CrossRef]
- Kwan, M.L.; Kushi, L.H.; Weltzien, E.; Maring, B.; Kutner, S.E.; Fulton, R.S.; Lee, M.M.; Ambrosone, C.B.; Caan, B.J. Epidemiology of breast cancer subtypes in two prospective cohort studies of breast cancer survivors. Breast Cancer Res. 2009, 11, R31. [Google Scholar] [CrossRef] [PubMed]
- Amirikia, K.C.; Mills, P.; Bush, J.; Newman, L.A. Higher population-based incidence rates of triple-negative breast cancer among young African-American women: Implications for breast cancer screening recommendations. Cancer 2011, 117, 2747–2753. [Google Scholar] [CrossRef] [PubMed]
- Mavaddat, N.; Antoniou, A.C.; Easton, D.F.; Garcia-Closas, M. Genetic susceptibility to breast cancer. Mol. Oncol. 2010, 4, 174–191. [Google Scholar] [CrossRef]
- Pharoah, P.D.P.; Antoniou, A.C.; Easton, D.F.; Ponder, B.A.J. Polygenes, risk prediction, and targeted prevention of breast cancer. N. Engl. J. Med. 2008, 358, 2796–2803. [Google Scholar] [CrossRef] [PubMed]
- Gieger, C.; Geistlinger, L.; Altmaier, E.; Hrabé de Angelis, M.; Kronenberg, F.; Meitinger, T.; Mewes, H.W.; Wichmann, H.E.; Weinberger, K.M.; Adamski, J.; et al. Genetics meets metabolomics: A genome-wide association study of metabolite profiles in human serum. PLoS Genet. 2008, 4, e1000282. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Shen, J.; Moore, S.C.; Ye, Y.; Wu, X.; Esteva, F.J.; Tripathy, D.; Chow, W.H. Breast cancer risk in relation to plasma metabolites among Hispanic and African American women. Breast Cancer Res. Treat. 2019, 176, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Huo, M.; Zhang, J.; Huang, W.; Wang, Y. Interplay among metabolism, epigenetic modifcations, and gene expression in cancer. Front. Cell Dev. Biol. 2021, 9, 793428. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Herman, J.G.; Baylin, S.B. Gene silencing in cancer in association with promoter hypermethylation. N. Engl. J. Med. 2003, 349, 2042–2054. [Google Scholar] [CrossRef]
- Garcia-Martinez, L.; Zhang, Y.; Nakata, Y.; Chan, H.L.; Morey, L. Epigenetic mechanisms in breast cancer therapy and resistance. Nat. Commun. 2021, 12, 1786. [Google Scholar] [CrossRef]
- Mogol, A.N.; Zuo, Q.; Yoo, J.Y.; Kaminsky, A.Z.; Imir, O.B.; Landesman, Y.; Walker, C.J.; Erdogan, Z.M. NAD+ metabolism generates a metabolic vulnerability in endocrine-resistant metastatic breast tumors in females. Endocrinology 2023, 164, bqad073. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.C.; Kadlubar, S.; Su, L.J.; Acheampong, D.; Rogers, L.J.; Runnells, G.; McElfish, P.A.; Schootman, M. County poverty levels infuence genome-wide DNA methylation profles in African American and European American women. Transl. Cancer Res. 2019, 8, 683–692. [Google Scholar] [CrossRef]
- Moore, S.C.; Playdon, M.C.; Sampson, J.N.; Hoover, R.N.; Trabert, B.; Matthews, C.E.; Ziegler, R.G. A metabolomics analysis of body mass index and postmenopausal breast cancer risk. J. Natl. Cancer Inst. 2018, 110, 588–597. [Google Scholar] [CrossRef]
- Medina, V.; Cricco, G.; Nunez, M.; Martin, G.; Mohamad, N.; Correa-Fiz, F.; Sanchez-Jimenez, F.; Bergoc, R.; Rivera, E.S. Histamine-mediated signaling processes in human malignant mammary cells. Cancer Biol. Ther. 2006, 11, 1462–1471. [Google Scholar] [CrossRef]
- Wolfe, R.R. Branched-chain amino acids and muscle protein synthesis in humans: Myth or reality? J. Int. Soc. Sports Nutr. 2017, 14, 30. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Zeleznik, O.A.; Balasubramanian, R.; Ren, Y.; Tobias, D.K.; Rosner, B.A.; Peng, C.; Bever, A.M.; Frueh, L.; Jeanfavre, S.; Avila-Pacheco, J.; et al. Branched-Chain Amino Acids and Risk of Breast Cancer. JNCI Cancer Spectr. 2021, 5, pkab059. [Google Scholar] [CrossRef] [PubMed]
- Nagata, C.; Wada, K.; Tsuji, M.; Hayashi, M.; Takeda, N.; Yasuda, K. Plasma amino acid profiles are associated with biomarkers of breast cancer risk in premenopausal Japanese women. Cancer Causes Control 2014, 25, 143–149. [Google Scholar] [CrossRef]
- Lécuyer, L.; Victor Bala, A.; Deschasaux, M.; Bouchemal, N.; Nawfal Triba, M.; Vasson, M.P.; Rossary, A.; Demidem, A.; Galan, P.; Hercberg, S.; et al. NMR metabolomic signatures reveal predictive plasma metabolites associated with long-term risk of developing breast cancer. Int. J. Epidemiol. 2018, 47, 484–494. [Google Scholar] [CrossRef]
- His, M.; Viallon, V.; Dossus, L.; Gicquiau, A.; Achaintre, D.; Scalbert, A.; Ferrari, P.; Romieu, I.; Onland-Moret, N.C.; Weiderpass, E.; et al. Prospective analysis of circulating metabolites and breast cancer in EPIC. BMC Med. 2019, 17, 178. [Google Scholar] [CrossRef]
- Zhang, J.; Tao, B.; Chong, Y.; Ma, S.; Wu, G.; Zhu, H.; Zhao, Y.; Zhao, S.; Niu, M.; Zhang, S.; et al. Ornithine and breast cancer: A matched case-control study. Sci. Rep. 2020, 10, 15502. [Google Scholar] [CrossRef] [PubMed]
- Jobard, E.; Dossus, L.; Baglietto, L.; Fornili, M.; Lécuyer, L.; Mancini, F.R.; Gunter, M.J.; Trédan, O.; Boutron-Ruault, M.C.; Elena-Herrmann, B.; et al. Investigation of circulating metabolites associated with breast cancer risk by untargeted metabolomics: A case-control study nested within the French E3N cohort. Br. J. Cancer 2021, 124, 1734–1743. [Google Scholar] [CrossRef] [PubMed]
- Stevens, V.L.; Carter, B.D.; Jacobs, E.J.; McCullough, M.L.; Teras, L.R.; Wang, Y. A prospective case–cohort analysis of plasma metabolites and breast cancer risk. Breast Cancer Res. 2023, 25, 5. [Google Scholar] [CrossRef] [PubMed]
- Kühn, T.; Floegel, A.; Sookthai, D.; Johnson, T.; Rolle-Kampczyk, U.; Otto, W.; von Bergen, M.; Boeing, H.; Kaaks, R. Higher plasma levels of lysophosphatidylcholine 18:0 are related to a lower risk of common cancers in a prospective metabolomics study. BMC Med. 2016, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Lécuyer, L.; Dalle, C.; Lyan, B.; Demidem, A.; Rossary, A.; Vasson, M.P.; Petera, M.; Lagree, M.; Ferreira, T.; Centeno, D.; et al. Plasma metabolomic signatures associated with long-term breast cancer risk in the SU. VI. MAX prospective cohort. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1300–1307. [Google Scholar] [CrossRef]
- Jung, M.K.; Okekunle, A.P.; Lee, J.E.; Sung, M.K.; Lim, Y.J. Role of Branched-chain Amino Acid Metabolism in Tumor Development and Progression. J. Cancer Prev. 2021, 26, 237–243. [Google Scholar] [CrossRef]
- Chi, R.; Yao, C.; Chen, S.; Liu, Y.; He, Y.; Zhang, J.; Ellies, L.G.; Wu, X.; Zhao, Q.; Zhou, C.; et al. Elevated BCAA suppresses the development and metastasis of breast cancer. Front. Oncol. 2022, 12, 887257. [Google Scholar] [CrossRef]
- Tobias, D.K.; Chai, B.; Tamimi, R.M.; Manson, J.E.; Hu, F.B.; Willett, W.C.; Eliassen, A.H. Dietary Intake of Branched Chain Amino Acids and Breast Cancer Risk in the NHS and NHS II Prospective Cohorts. JNCI Cancer Spectr. 2021, 5, pkab032. [Google Scholar] [CrossRef]
- Nouri-Majd, S.; Salari-Moghaddam, A.; Benisi-Kohansal, S.; Azadbakht, L.; Esmaillzadeh, A. Dietary intake of branched-chain amino acids in relation to the risk of breast cancer. Breast Cancer 2022, 29, 993–1000. [Google Scholar] [CrossRef]
- Mrowiec, K.; Kurczyk, A.; Jelonek, K.; Debik, J.; Giskeødegård, G.F.; Bathen, T.F.; Widłak, P. Association of serum metabolome profile with the risk of breast cancer in participants of the HUNT2 study. Front. Oncol. 2023, 13, 1116806. [Google Scholar] [CrossRef]
- Risi, E.; Lisanti, C.; Vignoli, A.; Biagioni, C.; Paderi, A.; Cappadona, S.; Monte, F.D.; Moretti, E.; Sanna, G.; Livraghi, L.; et al. Risk assessment of disease recurrence in early breast cancer: A serum metabolomic study focused on elderly patients. Transl. Oncol. 2023, 27, 101585. [Google Scholar] [CrossRef]
- Hart, C.D.; Vignoli, A.; Tenori, L.; Uy, G.L.; Van To, T.; Adebamowo, C.; Hossain, S.M.; Biganzoli, L.; Risi, E.; Love, R.R.; et al. Serum Metabolomic Profiles Identify ER-Positive Early Breast Cancer Patients at Increased Risk of Disease Recurrence in a Multicenter Population. Clin. Cancer Res. 2017, 23, 1422–1431. [Google Scholar] [CrossRef]
- Yatabe, J.; Yatabe, M.S.; Ishibashi, K.; Nozawa, Y.; Sanada, H. Early detection of colon cancer by amino acid profiling using AminoIndex Technology: A case report. Diagn. Pathol. 2013, 8, 203. [Google Scholar] [CrossRef]
- Maeda, J.; Higashiyama, M.; Imaizumi, A.; Nakayama, T.; Yamamoto, H.; Daimon, T.; Yamakado, M.; Imamura, F.; Kodama, K. Possibility of multivariate function composed of plasma amino acid profiles as a novel screening index for non-small cell lung cancer: A case control study. BMC Cancer 2010, 10, 690. [Google Scholar] [CrossRef]
- Okamoto, N. Use of ‘AminoIndex Technology’ for cancer screening. Ningen Dock 2012, 26, 911–922. [Google Scholar]
- Jikuzono, T.; Ishibashi, O.; Kure, S.; Ohmae, Y.; Ohmae, T. Associations of AminoIndex Cancer Screening (Breast) Grade with Clinical and Laboratory Variables. J. Nippon Med. Sch. 2022, 89, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, N.; Miyagi, Y.; Chiba, A.; Shiozawa, M.; Akaike, M.; Imaizumi, A.; Ando, T.; Tochikubo, O. Multivariate discrimination functions composed with amino acid profiles (Amino Index®) as a novel diagnostic marker for breast and colon cancer. Cancer Prev. 2008, 6, 47–48. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, X.; Cheng, C.; Yu, W.; Yi, P. Metabolism of Amino Acids in Cancer. Front. Cell Dev. Biol. 2020, 8, 603837. [Google Scholar] [CrossRef] [PubMed]
- Geck, R.C.; Toker, A. Nonessential amino acid metabolism in breast cancer. Adv. Biol. Regul. 2016, 62, 11–17. [Google Scholar] [CrossRef]
- Subramani, R.; Poudel, S.; Smith, K.D.; Estrada, A.; Lakshmanaswamy, R. Metabolomics of Breast Cancer: A Review. Metabolites 2022, 12, 643. [Google Scholar] [CrossRef]
- Willmann, L.; Schlimpert, M.; Halbach, S.; Erbes, T.; Stickeler, E.; Kammerer, B. Metabolic profiling of breast cancer: Differences in central metabolism between subtypes of breast cancer cell lines. J. Chromatogr B Anal. Technol. Biomed. Life Sci. 2015, 1000, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Wang, Y.; Alatrash, N.; Weatherly, C.A.; Roy, D.; MacDonnell, F.M.; Armstrong, D.W. Altered profiles and metabolism of land d-amino acids in cultured human breast cancer cells vs. non-tumorigenic human breast epithelial cells. J. Pharm. Biomed. Anal. 2019, 164, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, T.; Cao, Y.; Gao, P.; Dong, J.; Fang, Y.; Fang, Z.; Sun, X.; Zhu, Z. A dried blood spot mass spectrometry metabolomic approach for rapid breast cancer detection. Onco Targets Ther. 2016, 9, 1389–1398. [Google Scholar] [PubMed]
- Jain, M.; Nilsson, R.; Sharma, S.; Madhusudhan, N.; Kitami, T.; Souza, A.L.; Kafri, R.; Kirschner, M.W.; Clish, C.B.; Mootha, V.K. Metabolite profiling identifies a key role for glycine in rapid cancer cell proliferation. Science 2012, 336, 1040–1044. [Google Scholar] [CrossRef]
- Craze, M.L.; Cheung, H.; Jewa, N.; Coimbra, N.D.M.; Soria, D.; El-Ansari, R.; Aleskandarany, M.A.; Wai Cheng, K.; Diez-Rodriguez, M.; Nolan, C.C.; et al. MYC regulation of glutamine-proline regulatory axis is key in luminal B breast cancer. Br. J. Cancer 2018, 118, 258–265. [Google Scholar] [CrossRef]
- Cha, Y.J.; Kim, E.S.; Koo, J.S. Amino Acid Transporters and Glutamine Metabolism in Breast Cancer. Int. J. Mol. Sci. 2018, 19, 907. [Google Scholar] [CrossRef]
- Coloff, J.L.; Murphy, J.P.; Braun, C.R.; Harris, I.S.; Shelton, L.M.; Kami, K.; Gygi, S.P.; Selfors, L.M.; Brugge, J.S. Differential glutamate metabolism in proliferating and quiescent mammary epithelial cells. Cell Metab. 2016, 23, 867–880. [Google Scholar] [CrossRef]
- El Ansari, R.; McIntyre, A.; Craze, M.L.; Ellis, I.O.; Rakha, E.A.; Green, A.R. Altered glutamine metabolism in breast cancer; subtype dependencies and alternative adaptations. Histopathology 2018, 72, 183–190. [Google Scholar] [CrossRef]
- Dowling, P.; Henry, M.; Meleady, P.; Clarke, C.; Gately, K.; O’Byrne, K.; Connolly, E.; Lynch, V.; Ballot, J.; Gullo, G.; et al. Metabolomic and proteomic analysis of breast cancer patient samples suggests that glutamate and 12-HETE in combination with CA15-3 may be useful biomarkers reflecting tumour burden. Metabolomics 2015, 11, 620–635. [Google Scholar] [CrossRef]
- Kandasamy, P.; Gyimesi, G.; Kanai, Y.; Hediger, M.A. Amino acid transporters revisited: New views in health and disease. Trends Biochem. Sci. 2018, 43, 752–789. [Google Scholar] [CrossRef]
- Bhutia, Y.D.; Babu, E.; Ramachandran, S.; Ganapathy, V. Amino Acid transporters in cancer and their relevance to “glutamine addiction”: Novel targets for the design of a new class of anticancer drugs. Cancer Res. 2015, 75, 1782–1788. [Google Scholar] [CrossRef] [PubMed]
- van Geldermalsen, M.; Wang, Q.; Nagarajah, R.; Marshall, A.D.; Thoeng, A.; Gao, D.; Ritchie, W.; Feng, Y.; Bailey, C.G.; Deng, N.; et al. ASCT2/SLC1A5 controls glutamine uptake and tumour growth in triple-negative basal-like breast cancer. Oncogene 2016, 35, 3201–3208. [Google Scholar] [CrossRef] [PubMed]
- Metallo, C.M.; Gameiro, P.A.; Bell, E.L.; Mattaini, K.R.; Yang, J.; Hiller, K.; Jewell, C.M.; Johnson, Z.R.; Irvine, D.J.; Guarente, L.; et al. Reductive glutamine metabolism by IDH1 mediates lipogenesis under hypoxia. Nature 2011, 481, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Le, A.; Lane, A.N.; Hamaker, M.; Bose, S.; Gouw, A.; Barbi, J.; Tsukamoto, T.; Rojas, C.J.; Slusher, B.S.; Zhang, H.; et al. Glucoseindependent glutamine metabolism via TCA cycling for proliferation and survival in B cells. Cell Metab. 2012, 15, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Meadows, A.L.; Kong, B.; Berdichevsky, M.; Roy, S.; Rosiva, R.; Blanch, H.W.; Clark, D.S. Metabolic and morphological differences between rapidly proliferating cancerous and normal breast epithelial cells. Biotechnol. Prog. 2008, 24, 334–341. [Google Scholar] [CrossRef]
- Budczies, J.; Pfitzner, B.M.; Györffy, B.; Winzer, K.-J.; Radke, C.; Dietel, M.; Fiehn, O.; Denkert, C. Glutamate enrichment as new diagnostic opportunity in breast cancer. Int. J. Cancer 2015, 136, 1619–1628. [Google Scholar] [CrossRef]
- Zhou, W.-X.; Chen, C.; Liu, X.-Q.; Li, Y.; Lin, Y.-L.; Wu, X.-T.; Kong, L.-Y.; Luo, J.-G. Discovery and optimization of withangulatin A derivatives as novel glutaminase 1 inhibitors for the treatment of triple-negative breast cancer. Eur. J. Med. Chem. 2021, 210, 112980. [Google Scholar] [CrossRef]
- Garcia-Caballero, M.; Neugebauer, E.; Campos, R.; Nunez de Castro, I.; Vara-Thorbeck, C. Increased histidine decarboxylase (HDC) activity in human colorectal cancer: Results of a study on ten patients. Agents Actions 1988, 23, 357–360. [Google Scholar] [CrossRef]
- Xie, G.; Zhou, B.; Zhao, A.; Qiu, Y.; Zhao, X.; Garmire, L.; Shvetsov, Y.B.; Yu, H.; Yen, Y.; Jia, W. Lowered circulating aspartate is a metabolic feature of human breast cancer. Oncotarget 2015, 6, 33369–33381. [Google Scholar] [CrossRef]
- Knott, S.R.V.; Wagenblast, E.; Khan, S.; Kim, S.Y.; Soto, M.; Wagner, M.; Turgeon, M.-O.; Fish, L.; Erard, N.; Gable, A.L.; et al. Asparagine bioavailability governs metastasis in a model of breast cancer. Nature 2018, 554, 378–381. [Google Scholar] [CrossRef]
- Sivanand, S.; Vander Heiden, M.G. Emerging roles for branched-chain amino acid metabolism in cancer. Cancer Cell 2020, 37, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Harvie, M.N.; Campbell, I.T.; Howell, A.; Thatcher, N. Acceptability and tolerance of a low tyrosine and phenylalanine diet in patients with advanced cancer—A pilot study. J. Hum. Nutr. Diet. 2002, 15, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Atoum, M.; Abdel-Fattah, M.; Nimer, N.; Abdel-Rahman, S.; Abdeldayem, S.A. Association of alanine-valine manganese superoxide dismutase gene polymorphism and microheterogeneity manganese superoxide dismutase activity in breast cancer and benign breast tissue. J. Breast Cancer 2012, 15, 157–161. [Google Scholar] [CrossRef]
- Nittoli, A.C.; Costantini, S.; Sorice, A.; Capone, F.; Ciarcia, R.; Marzocco, S.; Budillon, A.; Severino, L. Effects of α-zearalenol on the metabolome of two breast cancer cell lines by 1H-NMR approach. Metabolomics 2018, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Feng, Y.; Zhang, Y.; Zhu, X.; Jin, F. L-Arginine supplementation inhibits the growth of breast cancer by enhancing innate and adaptive immune responses mediated by suppression of MDSCs in vivo. BMC Cancer 2016, 16, 343. [Google Scholar] [CrossRef] [PubMed]
- Opitz, C.A.; Litzenburger, U.M.; Sahm, F.; Ott, M.; Tritschler, I.; Trump, S.; Schumacher, T.; Jestaedt, L.; Schrenk, D.; Weller, M.; et al. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature 2011, 478, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Yue, L.; Shi, J.; Shao, M.; Wu, T. Role of IDO and TDO in Cancers and Related Diseases and the Therapeutic Implications. J. Cancer 2019, 10, 2771–2782. [Google Scholar] [CrossRef]
- Cheong, J.E.; Sun, L. Targeting the IDO1/TDO2–KYN–AhR Pathway for Cancer Immunotherapy—Challenges and Opportunities. Trends Pharmacol. Sci. 2018, 39, 307–325. [Google Scholar] [CrossRef]
- Platten, M.; Wick, W.; Van de Eynde, B.J. Tryptophan Catabolism in Cancer: Beyond IDO and Tryptophan Depletion. Cancer Res. 2012, 72, 5435–5440. [Google Scholar] [CrossRef]
- Pollari, S.; Käkönen, S.M.; Edgren, H.; Wolf, M.; Kohonen, P.; Sara, H.; Guise, T.; Nees, M.; Kallioniemi, O. Enhanced serine production by bone metastatic breast cancer cells stimulates osteoclastogenesis. Breast Cancer Res. Treat. 2011, 125, 421–430. [Google Scholar] [CrossRef]
- Maddocks, O.D.K.; Athineos, D.; Cheung, E.C.; Lee, P.; Zhang, T.; van den Broek, N.J.F.; Mackay, G.M.; Labuschagne, C.F.; Gay, D.; Kruiswijk, F.; et al. Modulating the therapeutic response of tumours to dietary serine and glycine starvation. Nature 2017, 544, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Vettore, L.; Westbrook, R.L.; Tennant, D.A. New aspects of amino acid metabolism in cancer. Br. J. Cancer 2020, 122, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Lee, I.M.; Song, Y.; Cook, N.R.; Selhub, J.; Manson, J.E.; Buring, J.E.; Zhang, S.M. Plasma homocysteine and cysteine and risk of breast cancer in women. Cancer Res. 2010, 70, 2397–2405. [Google Scholar] [CrossRef]
- Bonifácio, V.D.B.; Pereira, S.A.; Serpa, J.; Vicente, J.B. Cysteine metabolic circuitries: Druggable targets in cancer. Br. J. Cancer 2021, 124, 862–879. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.J.; Chang, W.A.; Chuang, C.H.; Wu, K.L.; Cheng, C.H.; Sheu, C.C.; Hsu, Y.L.; Hung, J.Y. Cysteinyl leukotriene pathway and cancer. Int. J. Mol. Sci. 2021, 23, 120. [Google Scholar] [CrossRef] [PubMed]
- Haukaas, T.H.; Euceda, L.R.; Giskeødegård, G.F.; Bathen, T.F. Metabolic Portraits of Breast Cancer by HR MAS MR Spectroscopy of Intact Tissue Samples. Metabolites 2017, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Cho, H.T.; Williams, L.; Zhu, A.; Liang, K.; Huang, K.; Wu, H.; Jiang, C.; Hong, S.; Crowe, R.; et al. Potential biomarker of L-type amino acid transporter 1 in breast cancer progression. Nucl. Med. Mol. Imaging 2011, 45, 93–102. [Google Scholar] [CrossRef]
- Ulaner, G.A.; Schuster, D.M. Amino Acid Metabolism as a Target for Breast Cancer Imaging. PET Clin. 2018, 13, 437–444. [Google Scholar] [CrossRef]
- Hayashi, K.; Anzai, N. L-type amino acid transporter 1 as a target for inflammatory disease and cancer immunotherapy. J. Pharmacol. Sci. 2022, 148, 31–40. [Google Scholar] [CrossRef]
- Bhat, H.K.; Vadgama, J.V. Role of estrogen receptor in the regulation of estrogen induced amino acid transport of System A in breast cancer and other receptor positive tumor cells. Int. J. Mol. Med. 2002, 9, 271–279. [Google Scholar] [CrossRef]
- Shennan, D.B.; Thomson, J. Inhibition of system L (LAT1/CD98hc) reduces the growth of cultured human breast cancer cells. Oncol. Rep. 2008, 20, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.; Choi, C.M.; Ahn, S.H.; Lee, J.W.; Gong, G.; Ryu, J.S.; Oh, S.J.; Bacher-Stier, C.; Fels, L.; Koglin, N.; et al. Exploratory clinical trial of (4s)-4-(3-[18f]fluoropropyl)-l-glutamate for imaging xc-transporter using positron emission tomography in patients with non-small cell lung or breast cancer. Clin. Cancer Res. 2012, 18, 5427–5437. [Google Scholar] [CrossRef]
- Saito, Y.; Soga, T. Amino acid transporters as emerging therapeutic targets in cancer. Cancer Sci. 2021, 112, 2958–2965. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, P.; Lapela, M.; Någren, K.; Lehikoinen, P.; Minn, H.; Jyrkkiö, S. Preliminary study of carbon-11 methionine PET in the evaluation of early response to therapy in advanced breast cancer. Nucl. Med. Commun. 2009, 30, 30–36. [Google Scholar] [CrossRef]
- Mehravi, B.; Ardestani, M.S.; Damercheli, M.; Soltanghoraee, H.; Ghanaldarlaki, N.; Alizadeh, A.M.; Oghabian, M.A.; Shirazi, M.S.; Mahernia, S.; Amanlou, M. Breast Cancer Cells Imaging by Targeting Methionine Transporters with Gadolinium-Based Nanoprobe. Mol. Imaging Biol. 2014, 16, 519–528. [Google Scholar] [CrossRef]
- Taylor, K.M.; Kim, J.S.; Rieter, W.J.; An, H.; Lin, W.; Lin, W. Mesoporous silica nanospheres as highly efficient MRI contrast agents. J. Am. Chem. Soc. 2008, 130, 2154–2155. [Google Scholar] [CrossRef]
- Okudaira, H.; Nakanishi, T.; Oka, S.; Kobayashi, M.; Tamagami, H.; Schuster, D.M.; Goodman, M.M.; Shirakami, Y.; Tamai, I.; Kawai, K. Kinetic analyses of trans-1-amino-3-[18F]fluorocyclobutanecarboxylic acid transport in Xenopus laevis oocytes expressing human ASCT2 and SNAT2. Nucl. Med. Biol. 2013, 40, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Okudaira, H.; Ono, M.; Schuster, D.M.; Goodman, M.M.; Kawai, K.; Shirakami, Y. Differences in transport mechanisms of trans-1-amino-3-[18F] fluorocyclobutanecarboxylic acid in inflammation, prostate cancer, and glioma cells: Comparison with L-[methyl-11C]methionine and 2-deoxy-2-[18F]fluoro-D-glucose. Mol. Imaging Biol. 2014, 16, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Tade, F.I.; Cohen, M.A.; Styblo, T.M.; Odewole, O.A.; Holbrook, A.I.; Newell, M.S.; Savir-Baruch, B.; Li, X.; Goodman, M.M.; Nye, J.A.; et al. Anti-3-18FFACBC (18F-Fluciclovine) PET/CT of breast cancer: An exploratory study. J. Nucl. Med. 2016, 57, 1357–1363. [Google Scholar] [CrossRef]
- Ulaner, G.A.; Goldman, D.A.; Corben, A.; Lyashchenko, S.K.; Gönen, M.; Lewis, J.S.; Dickler, M. Prospective clinical trial of 18F-fluciclovine PET/CT for determining the response to neoadjuvant therapy in invasive ductal and invasive lobular breast cancers. J. Nucl. Med. 2017, 58, 1037–1042. [Google Scholar] [CrossRef]
- Ulaner, G.A.; Goldman, D.A.; Gonen, M.; Pham, H.; Castillo, R.; Lyashchenko, S.K.; Lewis, J.S.; Dang, C. Initial results of a prospective clinical trial of 18f-fluciclovine pet/ct in newly diagnosed invasive ductal and invasive lobular breast cancers. J. Nucl. Med. 2016, 57, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Kanagawa, M.; Doi, Y.; Schuster, D.M.; Goodman, M.M.; Yoshimura, H. Fasting enhances the contrast of bone metastatic lesions in 18F-fluciclovine-PET: Preclinical study using a rat model of mixed osteolytic/osteoblastic bone metastases. Int. J. Mol. Sci. 2017, 18, 934. [Google Scholar] [CrossRef] [PubMed]
- Koglin, N.; Mueller, A.; Berndt, M.; Schmitt-Willich, H.; Toschi, L.; Stephens, A.W.; Gekeler, V.; Friebe, M.; Dinkelborg, L.M. Specific PET imaging of xC-transporter activity using a (1)(8)Flabeled glutamate derivative reveals a dominant pathway in tumor metabolism. Clin. Cancer Res. 2011, 17, 6000–6011. [Google Scholar] [CrossRef]
- Yang, H.; Jenni, S.; Colovic, M.; Merkens, H.; Poleschuk, C.; Rodrigo, I.; Miao, Q.; Johnson, B.F.; Rishel, M.J.; Sossi, V.; et al. (18)f-5-fluoroaminosuberic acid as a potential tracer to gauge oxidative stress in breast cancer models. J. Nucl. Med. 2017, 58, 367–373. [Google Scholar] [CrossRef]
- Kole, A.C.; Nieweg, O.E.; Pruim, J.; Paans, A.M.; Plukker, J.T.; Hoekstra, H.J.; Schraffordt Koops, H.; Vaalburg, W. Standardized uptake value and quantification of metabolism for breast cancer imaging with FDG and L-[1-11C]tyrosine PET. J. Nucl. Med. 1997, 38, 692–696. [Google Scholar] [PubMed]
- Kong, F.L.; Ali, M.S.; Rollo, A.; Smith, D.L.; Zhang, Y.; Yu, D.F.; Yang, D.J. Development of tyrosine-based radiotracer 99mTc-N4-Tyrosine for breast cancer imaging. J. Biomed. Biotechnol. 2012, 2012, 671708. [Google Scholar] [CrossRef]
- Kaim, A.H.; Weber, B.; Kurrer, M.O.; Westera, G.; Schweitzer, A.; Gottschalk, J.; von Schulthess, G.K.; Buck, A. (18)f-fdg and (18)f-fet uptake in experimental soft tissue infection. Eur. J. Nucl. Med. Mol. Imaging 2002, 29, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Rau, F.C.; Weber, W.A.; Wester, H.J.; Herz, M.; Becker, I.; Kruger, A.; Schwaiger, M.; Senekowitsch-Schmidtke, R. O-(2-[(18)f]fluoroethyl)-l-tyrosine (fet): A tracer for differentiation of tumour from inflammation in murine lymph nodes. Eur. J. Nucl. Med. Mol. Imaging 2002, 29, 1039–1046. [Google Scholar] [CrossRef]
- Ren, W.; Liu, G.; Yin, J.; Tan, B.; Wu, G.; Bazer, F.W.; Peng, Y.; Yin, Y. Amino-acid transporters in t-cell activation and differentiation. Cell Death Dis. 2017, 8, e2655. [Google Scholar] [CrossRef]
- Yu, W.; McConathy, J.; Olson, J.J.; Goodman, M.M. System a amino acid transport-targeted brain and systemic tumor PET imaging agents 2-amino-3-[(18)F]fluoro-2-methylpropanoic acid and 3-[(18)F]fluoro-2-methyl2-(methylamino)propanoic acid. Nucl. Med. Biol. 2015, 42, 8–18. [Google Scholar] [CrossRef]
- Way, J.D.; Wang, M.; Hamann, I.; Wuest, M.; Wuest, F. Synthesis and evaluation of 2-amino-5-(4-[(18)f]fluorophenyl)pent-4-ynoic acid ([(18)f]fphpa): A novel (18)f-labeled amino acid for oncologic pet imaging. Nucl. Med. Biol. 2014, 41, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Michelhaugh, S.K.; Muzik, O.; Guastella, A.R.; Klinger, N.V.; Polin, L.A.; Cai, H.; Xin, Y.; Mangner, T.J.; Zhang, S.; Juhász, C.; et al. Assessment of tryptophan uptake and kinetics using 1-(2-18F-fluoroethyl)-l-tryptophan and alpha-11Cmethyl-l-tryptophan PET imaging in mice implanted with patient-derived brain tumor xenografts. J. Nucl. Med. 2017, 58, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Chin, B.B.; McDougald, D.; Weitzel, D.H.; Hawk, T.; Reiman, R.E.; Zalutsky, M.R.; Vaidyanathan, G. Synthesis and preliminary evaluation of 5-[18F]fluoroleucine. Curr. Radiopharm. 2017, 10, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Pantel, A.R.; Li, S.; Lieberman, B.P.; Ploessl, K.; Choi, H.; Blankemeyer, E.; Lee, H.; Kung, H.F.; Mach, R.H.; et al. [18F](2S,4R)4-fluoroglutamine PET detects glutamine pool size changes in triple-negative breast cancer in response to glutaminase inhibition. Cancer Res. 2017, 77, 1476–1484. [Google Scholar] [CrossRef] [PubMed]
- Kopka, K.; Riemann, B.; Friedrich, M.; Winters, S.; Halfter, H.; Weckesser, M.; Stögbauer, F.; Bernd Ringelstein, E.; Schober, O. Characterization of 3-[123i]iodo-l-α-methyl tyrosine transport in astrocytes of neonatal rats. J. Neurochem. 2001, 76, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Jager, P.L.; Franssen, E.J.; Kool, W.; Szabo, B.G.; Hoekstra, H.J.; Groen, H.J.; de Vries, E.G.; van Imhoff, G.W.; Vaalburg, W.; Piers, D.A. Feasibility of tumor imaging using l-3-[iodine-123]-iodo-alpha-methyl-tyrosine in extracranial tumors. J. Nucl. Med. 1998, 39, 1736–1743. [Google Scholar]
- Sharma, S.; Singh, B.; Mishra, A.K.; Rathod, D.; Hazari, P.P.; Chuttani, K.; Chopra, S.; Singh, P.M.; Abrar, M.L.; Mittal, B.R.; et al. Lat-1 based primary breast cancer detection by [99m]tc-labeled dtpa-bis-methionine scintimammography: First results using indigenously developed single vial kit preparation. Cancer Biotherapy Radiopharm. 2014, 29, 283–288. [Google Scholar] [CrossRef]
- Brandon, F.; Ashley, S. Deberardinis RJ. Metabolic reprogramming and cancer progression. Science 2020, 368, eaaw5473. [Google Scholar]
- Chowdhry, S.; Zanca, C.; Rajkumar, U.; Koga, T.; Diao, Y.; Raviram, R.; Liu, F.; Turner, K.; Yang, H.; Brunk, E.; et al. NAD metabolic dependency in cancer is shaped by gene amplification and enhancer remodelling. Nature 2019, 569, 570–575. [Google Scholar] [CrossRef]
- Bian, Y.; Li, W.; Kremer, D.M.; Sajjakulnukit, P.; Li, S.; Crespo, J.; Nwosu, Z.C.; Zhang, L.; Czerwonka, A.; Pawłowska, A.; et al. Cancer SLC43A2 alters T cell methionine metabolism and histone methylation. Nature 2020, 585, 277–282. [Google Scholar] [CrossRef]
- Gao, X.; Sanderson, S.M.; Dai, Z.; Reid, M.A.; Cooper, D.E.; Lu, M.; Richie, J.P., Jr.; Ciccarella, A.; Calcagnotto, A.; Mikhael, P.G.; et al. Dietary methionine influences therapy in mouse cancer models and alters human metabolism. Nature 2019, 572, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Kanarek, N.; Keys, H.R.; Cantor, J.R.; Lewis, C.A.; Chan, S.H.; Kunchok, T.; Abu-Remaileh, M.; Freinkman, E.; Schweitzer, L.D.; Sabatini, D.M. Histidine catabolism is a major determinant of methotrexate sensitivity. Nature 2018, 559, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Sadik, A.; Somarribas Patterson, L.F.; Öztürk, S.; Mohapatra, S.R.; Panitz, V.; Secker, P.F.; Pfänder, P.; Loth, S.; Salem, H.; Prentzell, M.T.; et al. IL4I1 is a metabolic immune checkpoint that activates the AHR and promotes tumor progression. Cell 2020, 182, 1252–1270. [Google Scholar] [CrossRef] [PubMed]
- Strekalova, E.; Malin, D.; Weisenhorn, E.M.M.; Russell, J.D.; Hoelper, D.; Jain, A.; Coon, J.J.; Lewis, P.W.; Cryns, V.L. S-adenosylmethionine biosynthesis is a targetable metabolic vulnerability of cancer stem cells. Breast Cancer Res. Treat. 2019, 175, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Li, L.; Coyaud, E.; Luna, A.; Sander, C.; Raught, B.; Asara, J.M.; Brown, M.; Muthuswamy, S.K. LLGL2 rescues nutrient stress by promoting leucine uptake in ER+ breast cancer. Nature 2019, 569, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Y.; Li, C.F.; Kuo, C.C.; Tsai, K.K.; Hou, M.F.; Hung, W.C. Cancer/stroma interplay via cyclooxygenase-2 and indoleamine 2,3-dioxygenase promotes breast cancer progression. Breast Cancer Res. 2014, 16, 410. [Google Scholar] [CrossRef]
- Greene, L.I.; Bruno, T.C.; Christenson, J.L.; D’Alessandro, A.; Culp-Hill, R.; Torkko, K.; Borges, V.F.; Slansky, J.E.; Richer, J.K. A Role for Tryptophan-2,3-dioxygenase in CD8 T-cell Suppression and Evidence of Tryptophan Catabolism in Breast Cancer Patient Plasma. Mol. Cancer Res. 2019, 17, 131–139. [Google Scholar] [CrossRef]
- Zhao, Y.; Pu, C.; Liu, Z. Essential amino acids deprivation is a potential strategy for breast cancer treatment. Breast 2022, 62, 152–161. [Google Scholar] [CrossRef]
- Zhang, L.; Han, J. Branched-chain amino acid transaminase 1 (BCAT1) promotes the growth of breast cancer cells through improving mTOR-mediated mitochondrial biogenesis and function. Biochem. Biophys. Res. Commun. 2017, 486, 224–231. [Google Scholar] [CrossRef]
- El Ansari, R.; Craze, M.L.; Miligy, I.; Diez-Rodriguez, M.; Nolan, C.C.; Ellis, I.O.; Rakha, E.A.; Green, A.R. The amino acid transporter SLC7A5 confers a poor prognosis in the highly proliferative breast cancer subtypes and is a key therapeutic target in luminal B tumours. Breast Cancer Res. 2018, 20, 21. [Google Scholar] [CrossRef]
- Cao, M.D.; Lamichhane, S.; Lundgren, S.; Bofin, A.; Fjøsne, H.; Giskeødegård, G.F.; Bathen, T.F. Metabolic characterization of triple negative breast cancer. BMC Cancer 2014, 14, 941. [Google Scholar] [CrossRef]
- Lampa, M.; Arlt, H.; He, T.; Ospina, B.; Reeves, J.; Zhang, B.; Murtie, J.; Deng, G.; Barberis, C.; Hoffmann, D.; et al. Glutaminase is essential for the growth of triple-negative breast cancer cells with a deregulated glutamine metabolism pathway and its suppression synergizes with mTOR inhibition. PLoS ONE 2017, 12, e0185092. [Google Scholar] [CrossRef] [PubMed]
- Mates, J.M.; Campos-Sandoval, J.A.; Marquez, J. Glutaminase isoenzymes in the metabolic therapy of cancer. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Stalnecker, C.; Zhang, C.; McDermott, L.A.; Iyer, P.; O’Neill, J.; Reimer, S.; Cerione, R.A.; Katt, W.P. Characterization of the interactions of potent allosteric inhibitors with glutaminase C, a key enzyme in cancer cell glutamine metabolism. J. Biol. Chem. 2018, 293, 3535–3545. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Fu, R.; Duan, Z.; Shen, S.; Zhu, C.; Fan, D. Ginsenoside CK induces apoptosis in triple-negative breast cancer cells by targeting glutamine metabolism. Biochem. Pharmacol. 2022, 202, 115101. [Google Scholar] [CrossRef]
- Morotti, M.; Zois, C.E.; El-Ansari, R.; Craze, M.L.; Rakha, E.A.; Fan, S.J.; Valli, A.; Haider, S.; Goberdhan, D.C.I.; Green, A.R.; et al. Increased expression of glutamine transporter SNAT2/SLC38A2 promotes glutamine dependence and oxidative stress resistance, and is associated with worse prognosis in triple-negative breast cancer. Br. J. Cancer 2021, 124, 494–505. [Google Scholar] [CrossRef]
- Timmerman, L.A.; Holton, T.; Yuneva, M.; Louie, R.J.; Padró, M.; Daemen, A.; Hu, M.; Chan, D.A.; Ethier, S.P.; van ‘t Veer, L.J.; et al. Glutamine sensitivity analysis identifies the xCT antiporter as a common triple-negative breast tumor therapeutic target. Cancer Cell 2013, 24, 450–465. [Google Scholar] [CrossRef]
- Lanzardo, S.; Conti, L.; Rooke, R.; Ruiu, R.; Accart, N.; Bolli, E.; Arigoni, M.; Macagno, M.; Barrera, G.; Pizzimenti, S.; et al. Immunotargeting of Antigen xCT Attenuates Stem-like Cell Behavior and Metastatic Progression in Breast Cancer. Cancer Res. 2016, 76, 62–72. [Google Scholar] [CrossRef]
- Bolli, E.; O’Rourke, J.P.; Conti, L.; Lanzardo, S.; Rolih, V.; Christen, J.M.; Barutello, G.; Forni, M.; Pericle, F.; Cavallo, F. A Virus-Like-Particle immunotherapy targeting Epitope-Specific anti-xCT expressed on cancer stem cell inhibits the progression of metastatic cancer in vivo. Oncoimmunology 2017, 7, e1408746. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, Q.; Dong, C. Metabolic reprogramming in triple-negative breast cancer. Cancer Biol. Med. 2020, 17, 44–59. [Google Scholar] [CrossRef]
- Strekalova, E.; Malin, D.; Good, D.M.; Cryns, V.L. Methionine deprivation induces a targetable vulnerability in triple-negative breast cancer cells by enhancing TRAIL receptor-2 expression. Clin. Cancer Res. 2015, 21, 2780–2791. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Kim, J.H.; Lee, E.; Jang, Y.J.; Son, J.E.; Kwon, J.Y.; Lim, T.G.; Kim, S.; Park, J.H.; Kim, J.E.; et al. Methionine deprivation suppresses triple-negative breast cancer metastasis in vitro and in vivo. Oncotarget 2016, 7, 67223–67234. [Google Scholar] [CrossRef] [PubMed]
- Borrego, S.L.; Fahrmann, J.; Datta, R.; Stringari, C.; Grapov, D.; Zeller, M.; Chen, Y.; Wang, P.; Baldi, P.; Gratton, E.; et al. Metabolic changes associated with methionine stress sensitivity in MDA-MB-468 breast cancer cells. Cancer Metab. 2016, 4, 9. [Google Scholar] [CrossRef]
- Wang, Z.; Yip, L.Y.; Lee, J.H.J.; Wu, Z.; Chew, H.Y.; Chong, P.K.W.; Teo, C.C.; Ang, H.Y.; Peh, K.L.E.; Yuan, J.; et al. Methionine is a metabolic dependency of tumor-initiating cells. Nat. Med. 2019, 25, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shi, X.; Li, Y.; Fan, J.; Zeng, X.; Xian, Z.; Wang, Z.; Sun, Y.; Wang, S.; Song, P.; et al. Blocking autophagy enhanced cytotoxicity induced by recombinant human arginase in triple-negative breast cancer cells. Cell Death Dis. 2014, 5, e1563. [Google Scholar] [CrossRef] [PubMed]
- Hassabo, A.A.; Abdelraof, M.; Allam, R.M. L-arginase from Streptomyces diastaticus MAM5 as a potential therapeutic agent in breast cancer: Purification, characterization, G1 phase arrest and autophagy induction. Int. J. Biol. Macromol. 2023, 224, 634–645. [Google Scholar] [CrossRef]
- Wang, S.M.; Dowhan, D.H.; Muscat, G.E.O. Epigenetic arginine methylation in breast cancer: Emerging therapeutic strategies. J. Mol. Endocrinol. 2019, 62, R223–R237. [Google Scholar] [CrossRef]
- Oh, T.G.; Wang, S.M.; Muscat, G.E. Therapeutic implications of epigenetic signaling in breast cancer. Endocrinology 2017, 158, 431–447. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Khelifa, S.; Ratnikov, B.; Scott, D.A.; Feng, Y.; Parisi, F.; Ruller, C.; Lau, E.; Kim, H.; Brill, L.M.; et al. Regulation of glutamine carrier proteins by RNF5 determines breast Cancer response to ER stress-inducing chemotherapies. Cancer Cell 2015, 27, 354–369. [Google Scholar] [CrossRef]
- Broer, A.; Rahimi, F.; Broer, S. Deletion of amino acid transporter ASCT2 (SLC1A5) reveals an essential role for transporters SNAT1 (SLC38A1) and SNAT2 (SLC38A2) to sustain Glutaminolysis in Cancer cells. J. Biol. Chem. 2016, 291, 13194–13205. [Google Scholar] [CrossRef]
- Dhankhar, R.; Gupta, V.; Kumar, S.; Kapoor, R.K.; Gulati, P. Microbial enzymes for deprivation of amino acid metabolism in malignant cells: Biological strategy for cancer treatment. Appl. Microbiol. Biotechnol. 2020, 104, 2857–2869. [Google Scholar] [CrossRef] [PubMed]
- Gwangwa, M.V.; Joubert, A.M.; Visagie, M.H. Effects of glutamine deprivation on oxidative stress and cell survival in breast cell lines. Biol. Res. 2019, 52, 15. [Google Scholar] [CrossRef] [PubMed]
- Bacci, M.; Lorito, N.; Ippolito, L.; Ramazzotti, M.; Luti, S.; Romagnoli, S.; Parri, M.; Bianchini, F.; Cappellesso, F.; Virga, F.; et al. Reprogramming of Amino Acid Transporters to Support Aspartate and Glutamate Dependency Sustains Endocrine Resistance in Breast Cancer. Cell Rep. 2019, 28, 104–118. [Google Scholar] [CrossRef] [PubMed]
- Maeng, H.J.; Kim, E.S.; Chough, C.; Joung, M.; Lim, J.W.; Shim, C.K.; Shim, W.S. Addition of amino acid moieties to lapatinib increases the anti-cancer effect via amino acid transporters. Biopharm. Drug Dispos. 2014, 35, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Mello-Andrade, F.; Guedes, A.P.M.; Pires, W.C.; Velozo-Sá, V.S.; Delmond, K.A.; Mendes, D.; Molina, M.S.; Matuda, L.; de Sousa, M.A.M.; Melo-Reis, P.; et al. Ru(II)/amino acid complexes inhibit the progression of breast cancer cells through multiple mechanism-induced apoptosis. J. Inorg. Biochem. 2022, 226, 111625. [Google Scholar] [CrossRef]
- Yoo, H.C.; Han, J.M. Amino acid metabolism in cancer drug resistance. Cells 2022, 11, 140. [Google Scholar] [CrossRef]
- Mitrevska, K.; Rodrigo, M.A.M.; Cernei, N.; Michalkova, H.; Splichal, Z.; Hynek, D.; Zitka, O.; Heger, Z.; Kopel, P.; Adam, V.; et al. Chick chorioallantoic membrane (CAM) assay for the evaluation of the antitumor and antimetastatic activity of platinum-based drugs in association with the impact on the amino acid metabolism. Mater. Today Bio 2023, 19, 100570. [Google Scholar] [CrossRef]
- Ji, Y.; Li, J.; Xiao, S.; Kwan, H.Y.; Bian, Z.; Chu, C.C. Optimization of amino acid-based poly(ester urea urethane) nanoparticles for the systemic delivery of gambogic acid for treating triple negative breast cancer. Biomater. Sci. 2023, 11, 4370–4384. [Google Scholar] [CrossRef]
- Dunstan, R.H.; Sparkes, D.L.; Macdonald, M.M.; Roberts, T.K.; Wratten, C.; Kumar, M.B.; Baines, S.; Denham, J.W.; Gallagher, S.A.; Rothkirch, T. Altered amino acid homeostasis and the development of fatigue by breast cancer radiotherapy patients: A pilot study. Clin. Biochem. 2011, 44, 208–215. [Google Scholar] [CrossRef]
- Poon, R.T.P.; Yu, W.C.; Fan, S.T.; Wong, J. Long-term oral branched chain amino acids in patients undergoing chemoembolization for hepatocellular carcinoma: A randomized trial. Aliment. Pharmacol. Ther. 2004, 19, 779–788. [Google Scholar] [CrossRef]
- Gramignano, G.; Lusso, M.R.; Madeddu, C.; Massa, E.; Serpe, R.; Deiana, L.; Lamonica, G.; Dessì, M.; Spiga, C.; Astara, G.; et al. Efficacy of L-carnitine administration on fatigue, nutritional status, oxidative stress, and related quality of life in 12 advanced cancer patients undergoing anticancer therapy. Nutrition 2006, 22, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lockwood, M.B.; Schlaeger, J.M.; Liu, T.; Danciu, O.C.; Doorenbos, A.Z. Tryptophan and Kynurenine Pathway Metabolites and Psychoneurological Symptoms Among Breast Cancer Survivors. Pain Manag. Nurs. 2023, 24, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Nees, J.; Schafferer, S.; Yuan, B.; Tang, Q.; Scheffler, M.; Hartkopf, A.; Golatta, M.; Schneeweiß, A.; Burwinkel, B.; Wallwiener, M. How previous treatment changes the metabolomic profile in patients with metastatic breast cancer. Arch. Gynecol. Obstet. 2022, 306, 2115–2122. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Jung, W.H.; Koo, J.S. Diferential expression of enzymes associated with serine/glycine metabolism in diferent breast cancer subtypes. PLoS ONE 2014, 9, e101004. [Google Scholar]
- Prokhorova, E.; Zobel, F.; Smith, R.; Zentout, S.; Gibbs-Seymour, I.; Schützenhofer, K.; Peters, A.; Groslambert, J.; Zorzini, V.; Agnew, T.; et al. Serine-linked PARP1 auto-modification controls PARP inhibitor response. Nat. Commun. 2021, 12, 4055. [Google Scholar] [CrossRef] [PubMed]
- Possemato, R.; Marks, K.M.; Shaul, Y.D.; Pacold, M.E.; Kim, D.; Birsoy, K.; Sethumadhavan, S.; Woo, H.K.; Jang, H.G.; Jha, A.K.; et al. Functional genomics reveal that the serine synthesis pathway is essential in breast cancer. Nature 2011, 476, 346–350. [Google Scholar] [CrossRef]


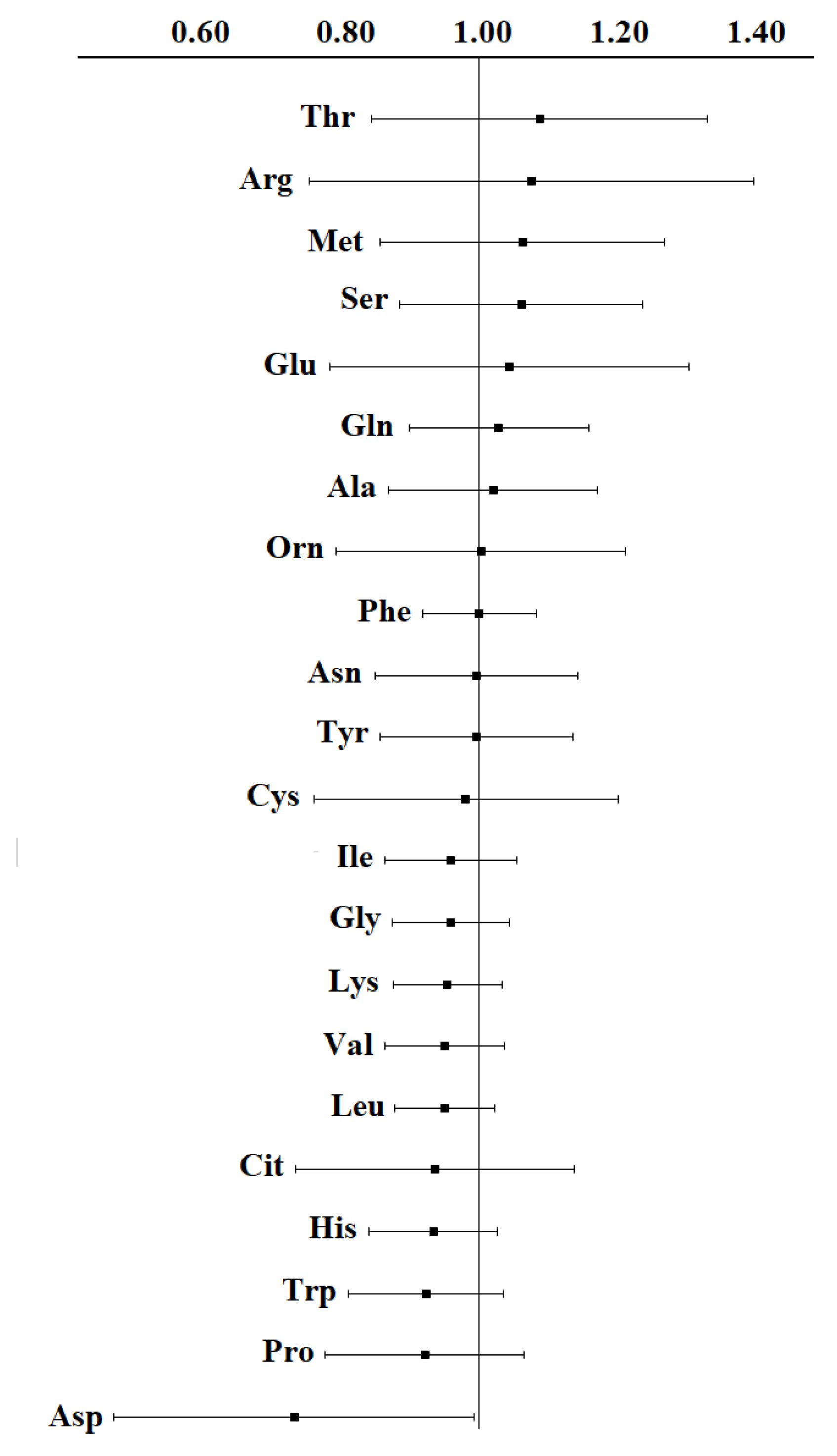{kind=link}
| No. | Author, Year | BC/Control | BC Stages | Method | Up-Regulated AAs | Down-Regulated AAs |
|---|---|---|---|---|---|---|
| 1 | Kubota A., 1992 [44] | 22/11 | St I + II—22 | AA analyzer | Ala, Arg, Thr | Cys, Gln |
| 2 | Cascino A., 1995 [45] | 33/28 | - | AA analyzer | Glu, Orn, Trp | - |
| 3 | Proenza A.M.A., 2003 [46] | 16/18 | St I—2, St II—5, St III—3, St IV—5, Unknown—1 | HPLC | Asn, Gln, Pro (Hydroxyproline) | Asp |
| 4 | Minet-Quinard R., 2004 [47] | 19/18 | T0—16%, T1—42%, T2—42% | AA analyzer | Ser, Glu, Orn | - |
| 5 | Vissers Y.L.J., 2005 [48] | 22/22 | St I—6, St II—7, St III—8 | HPLC | - | Arg |
| 6 | Okamoto N., 2009 [49] | 61/51 | St 0—8, St I—30, St II—18, St III–5 | HPLC–ESI–MS | Thr, Ser, Glu, Orn | Met, Ile, Phe, Arg |
| 7 | Miyagi Y., 2011 [34] | 196/976 | St II—95, St III—19, St IV—15, Unknown—5 | HPLC–ESI–MS | Thr, Pro, Ser, Gly, Ala, Orn | Gln, Trp, His, Phe, Tyr |
| 8 | Shen J., 2013 [8] | 60/60 | ER+/PR+—30, triple negative—30 African Americans and Caucasian Americans data sets | UPLC-MS/MS or GC-MS | - | Ala, His, Asp, Lys, Tyr, Trp, Met, Arg, Pro |
| 9 | Poschke I., 2013 [50] | 41/9 | - | HPLC | Glu, Ser, Gln, Ala, Val, Phe, Ile, Leu | - |
| 10 | Barnes T., 2014 [51] | 8/8 | St I + II—8 | HPLC | Ala, Asp, Gln, Lys, Met, Tyr | Arg, Gly, Pro, Ser, Ile, Val, Orn |
| 11 | Gu Y., 2015 [52] | 28/137 | St I—7, St II—18, St III—3 | AA analyzer | Thr, Arg | Asp, Gln, Gly, His |
| 12 | Jové M., 2017 [5] | 91/20 | St I—2, St IIA—40, St IIB—30, St IIIA—13, St IIIB—7 | ESI-Q-TOF MS/MS | - | Gln, Arg, Lys, Val |
| 13 | Wang X., 2018 [7] | 44/34 | - | UPLC-MS | Ala, Asp, Cys, Gly, Glu, Gln, His, Ile, Met, Pro, Phe, Ser, Tyr, Val | Arg |
| 14 | Jasbi P., 2018 [6] | 102/99 | St I—24, St II—42, St III—36 | UPLC-MS | Asp | Pro |
| 15 | Eniu D.T., 2018 [28] | 30/26 | St I—3, St II—17, St III—10 | HPLC-MS | - | Tyr, Arg, Ala, Ile, Trp, Leu |
| 16 | Cala M.P., 2018 [33] | 29/29 | St I—3, St II—15, St III—11 Colombian Hispanic data set | GC-MS | Val, Ala, Ile, Ser, Glu, 4-Hydroxyproline | - |
| 17 | Park J., 2019 [53] | 40/30 | St I—15, St II—15, St III—10 | HPLC-MS | 5-oxoproline, Phe, Ile + Leu | - |
| 18 | Li L., 2020 [54] | 105/35 | St I—65, St II—40 | NMR spectroscopy | Leu, Phe | Arg, Glu, Lys, Tyr, His |
| 19 | Politi C., 2021 [55] | 38/10 | - | GC-MS | Glu, Ile, Leu, Phe, Pro, Ser | Cys |
| 20 | An R., 2022 [56] | 75/20 | St I—31, St II—33, St III—11 | UPLC-MS | Gln, Arg | Glu, Asp, Cys |
| 21 | Baranovicova E., 2022 [57] | 50/46 | St I + II—50 | NMR spectroscopy | - | Leu, Ile, Val, Ala, His |
| 22 | Han X., 2022 [58] | 30/30 | ТНРМЖ | MALDI-TOF-MS | - | Ala, Ser, Pro, Val, Thr, His, Phe, Arg, Tyr, Trp |
| 23 | Santaliz-Casiano A., 2023 [59] | 103/150 | ER + breast cancer African American (AA) and non-Hispanic White (NHW) data sets | GC-MS | - | Arg (AA), His (AA), Met (AA) |
| 24 | Panigoro S.S., 2023 [60] | 29/28 | St I—13, St II—13, St III—3 | HPLC | Cys | Glu, His, Orn, Thr, Tyr, Val |
| Amino acid | Kubota, 1992 [44] | Proenza, 2003 [46] | Minet-Quinard, 2004 [47] | Vissers, 2005 [48] | Okamoto, 2009 [49] | Miyagi, 2011 [34] | Shen, 2013 [8] | Barnes, 2014 [51] | Gu, 2015 [52] | Wang, 2018 [7] | Eniu, 2018 [28] | Cala, 2018 [33] | Park, 2019 [53] | Baranovicova, 2022 [57] | An, 2022 [56] | Panigoro, 2023 [60] |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ala | 1.96 | 0.93 | 1.11 | 0.94 | 1.06 | 1.08 | 0.82/0.80 * | 1.16 | 0.93 | 1.11 | 0.59 | 1.25 | 0.80 | 0.92 | 0.88 | |
| Arg | 1.33 | 0.86 | 0.84 | 0.98 | 0.90/0.86 | 0.91 | 1.78 | 0.81 | 0.32 | 2.35 | 0.99 | |||||
| Asn | 1.54 | 0.95 | 0.91 | 0.97 | 0.99 | 0.84/0.82 | 1.16 | 1.23 | 0.65 | 0.91 | ||||||
| Asp | 0.45 | 0.80 | 1.33 | 0.67 | 0.71 | 0.43 | ||||||||||
| Cys | 0.58 | 0.93/1.01 | 1.04 | 1.31 | 0.97 | 0.66 | 1.35 | |||||||||
| Gln | 0.73 | 1.16 | 1.05 | 0.92 | 1.00 | 0.97 | 0.94/1.00 | 1.08 | 1.53 | 0.77 | 1.21 | |||||
| Glu | 1.02 | 1.42 | 1.24 | 1.40 | 1.05/1.01 | 0.46 | 1.81 | 0.42 | 1.27 | 0.58 | 0.86 | |||||
| Gly | 0.94 | 1.01 | 1.08 | 1.12 | 0.90/0.91 | 0.91 | 0.90 | 1.16 | 0.68 | 0.94 | ||||||
| His | 1.08 | 1.04 | 1.17 | 1.04 | 0.95 | 0.97 | 0.88/0.91 | 0.87 | 1.20 | 0.61 | 0.99 | 0.63 | 0.93 | 0.75 | ||
| Ile | 0.94 | 1.00 | 0.93 | 0.85 | 1.02 | 0.93/0.94 | 0.72 | 0.93 | 1.11 | 0.55 | 1.21 | 1.32 | 0.82 | 1.12 | 0.96 | |
| Leu | 1.00 | 0.81 | 0.94 | 1.00 | 0.94/0.93 | 0.95 | 1.15 | 0.64 | 0.85 | 0.95 | 0.88 | |||||
| Lys | 1.04 | 1.07 | 1.02 | 0.99 | 1.03 | 0.91/0.86 | 1.19 | 0.92 | 0.98 | 0.62 | 0.94 | 0.84 | ||||
| Met | 1.01 | 1.04 | 0.87 | 0.93 | 0.99 | 0.88/0.87 | 1.08 | 1.05 | 2.02 | 0.67 | 1.36 | |||||
| Phe | 1.09 | 1.08 | 0.83 | 0.89 | 0.98 | 0.92/0.90 | 1.00 | 1.00 | 1.21 | 0.78 | 1.30 | 1.04 | ||||
| Pro | 1.07 | 0.85 | 0.97 | 1.12 | 0.85/0.87 | 0.95 | 0.51 | 1.19 | 0.59 | 1.17 | ||||||
| Ser | 1.01 | 1.13 | 1.11 | 1.10 | 1.04 | 0.92/0.89 | 0.86 | 0.98 | 1.14 | 0.79 | 2.10 | 0.87 | 0.93 | |||
| Thr | 1.48 | 1.01 | 0.94 | 0.92 | 1.07 | 1.08 | 0.89/0.94 | 2.46 | 1.00 | 0.63 | 1.08 | 0.93 | 0.82 | |||
| Trp | 0.82 | 0.93 | 0.94 | 0.94/0.88 | 1.02 | 0.61 | 1.15 | 1.02 | ||||||||
| Tyr | 0.98 | 1.09 | 0.89 | 0.92 | 0.96 | 0.92/0.80 | 1.55 | 1.07 | 1.20 | 0.56 | 1.24 | 0.78 | ||||
| Val | 0.93 | 1.02 | 0.79 | 1.01 | 1.01 | 0.95/0.94 | 0.90 | 0.93 | 1.15 | 0.68 | 1.36 | 0.85 | 0.96 | 0.79 | ||
| Cit | 1.23 | 0.97 | 0.90 | 1.01 | 0.47 | 1.04 | ||||||||||
| Orn | 1.28 | 1.07 | 1.47 | 0.65 | 1.25 | 1.12 | 0.98 | 0.53 | 0.68 |
| No. | Author, Year | BC/Control | Subgroup | AAs | HR (95% CI) |
|---|---|---|---|---|---|
| 1 | Nagata C. et al., 2014 [88] | 350 | premenopausal | Arg, Leu, Tyr, Asp | - |
| 2 | Lécuyer L. et al., 2018 [89] | 206/396 | - | Val ↑ | 1.83 (1.15–2.92) |
| Gln ↑ | 1.61 (1.02–2.55) | ||||
| Lys + Creatine + Creatinine ↑ | 1.84 (1.19–2.85) | ||||
| Lys + Arg ↑ | 1.62 (1.05–2.48) | ||||
| 3 | His M. et al., 2019 [90] | 1624/1624 | - | Arg ↑ | 0.89 (0.80–0.99) |
| Asp ↑ | 0.87 (0.80–0.95) | ||||
| Gln ↑ | 0.91 (0.84–0.99) | ||||
| Gly ↑ | 0.90 (0.83–0.97) | ||||
| His ↑ | 0.91 (0.84–0.99) | ||||
| Lys ↑ | 0.90 (0.83–0.98) | ||||
| Thr ↑ | 0.92 (0.85–0.99) | ||||
| 4 | Zhang J. et al., 2020 [91] | 735/735 | - | Orn ↑ | 0.70 (0.53, 0.94) |
| 5 | Zeleznik O.A. et al., 2021 [87] | 1997/1997 | premenopausal | Ile ↑ | 0.86 (0.65–1.13) |
| postmenopausal | Ile ↑ | 1.63 (1.12–2.39) | |||
| 6 | Jobard E. et al., 2021 [92] | 791/791 | premenopausal | His ↑ | 1.70 (1.19–2.41) |
| Orn ↑ | 1.43 (1.06–1.95) | ||||
| Leu ↑ | 1.37 (1.01–1.86) | ||||
| Gln ↑ | 1.33 (1.00–1.78) | ||||
| Glu ↑ | 1.34 (1.00–1.79) | ||||
| 7 | Stevens V.L. et al., 2023 [93] | 1687/1983 | - | Ser ↓ | 0.89 (0.83–0.96) |
| Asp ↓ | 0.91 (0.84–0.97) | ||||
| Gln ↓ | 0.91 (0.85–0.98) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bel’skaya, L.V.; Gundyrev, I.A.; Solomatin, D.V. The Role of Amino Acids in the Diagnosis, Risk Assessment, and Treatment of Breast Cancer: A Review. Curr. Issues Mol. Biol. 2023, 45, 7513-7537. https://doi.org/10.3390/cimb45090474
Bel’skaya LV, Gundyrev IA, Solomatin DV. The Role of Amino Acids in the Diagnosis, Risk Assessment, and Treatment of Breast Cancer: A Review. Current Issues in Molecular Biology. 2023; 45(9):7513-7537. https://doi.org/10.3390/cimb45090474
Chicago/Turabian StyleBel’skaya, Lyudmila V., Ivan A. Gundyrev, and Denis V. Solomatin. 2023. "The Role of Amino Acids in the Diagnosis, Risk Assessment, and Treatment of Breast Cancer: A Review" Current Issues in Molecular Biology 45, no. 9: 7513-7537. https://doi.org/10.3390/cimb45090474
APA StyleBel’skaya, L. V., Gundyrev, I. A., & Solomatin, D. V. (2023). The Role of Amino Acids in the Diagnosis, Risk Assessment, and Treatment of Breast Cancer: A Review. Current Issues in Molecular Biology, 45(9), 7513-7537. https://doi.org/10.3390/cimb45090474