
Effect of YC-1102 on the Improvement of Obesity in High-Fat Diet-Induced Obese Mice



Abstract
:1. Introduction
2. Materials and Methods
2.1. YC-1102 Preparation and Standardization
2.2. Cell Viability Assay
2.3. Triglyceride Assay
2.4. Real-Time PCR
2.5. Western Blot Analysis
2.6. Animals and Experimental Design
2.7. Serological Assay
2.8. Histopathological Test
2.9. Statistical Analysis
3. Results
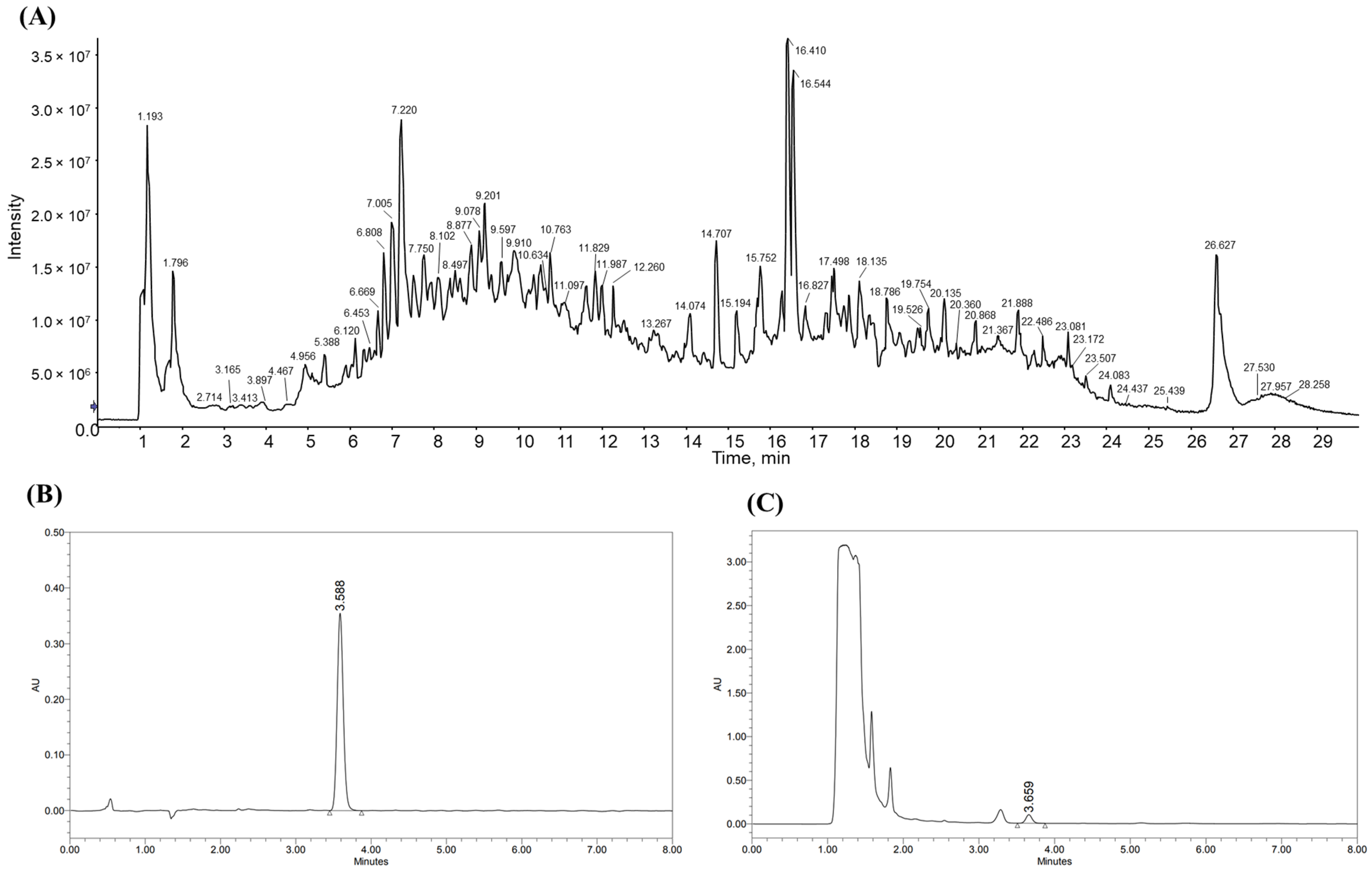
3.1. Identification of Rosamultin and Standardization of YC-1102 via LC-MS/MS and HPLC
3.2. YC-1102 Regulates Adipogenesis and Energy Metabolism in 3T3-L1 Cells
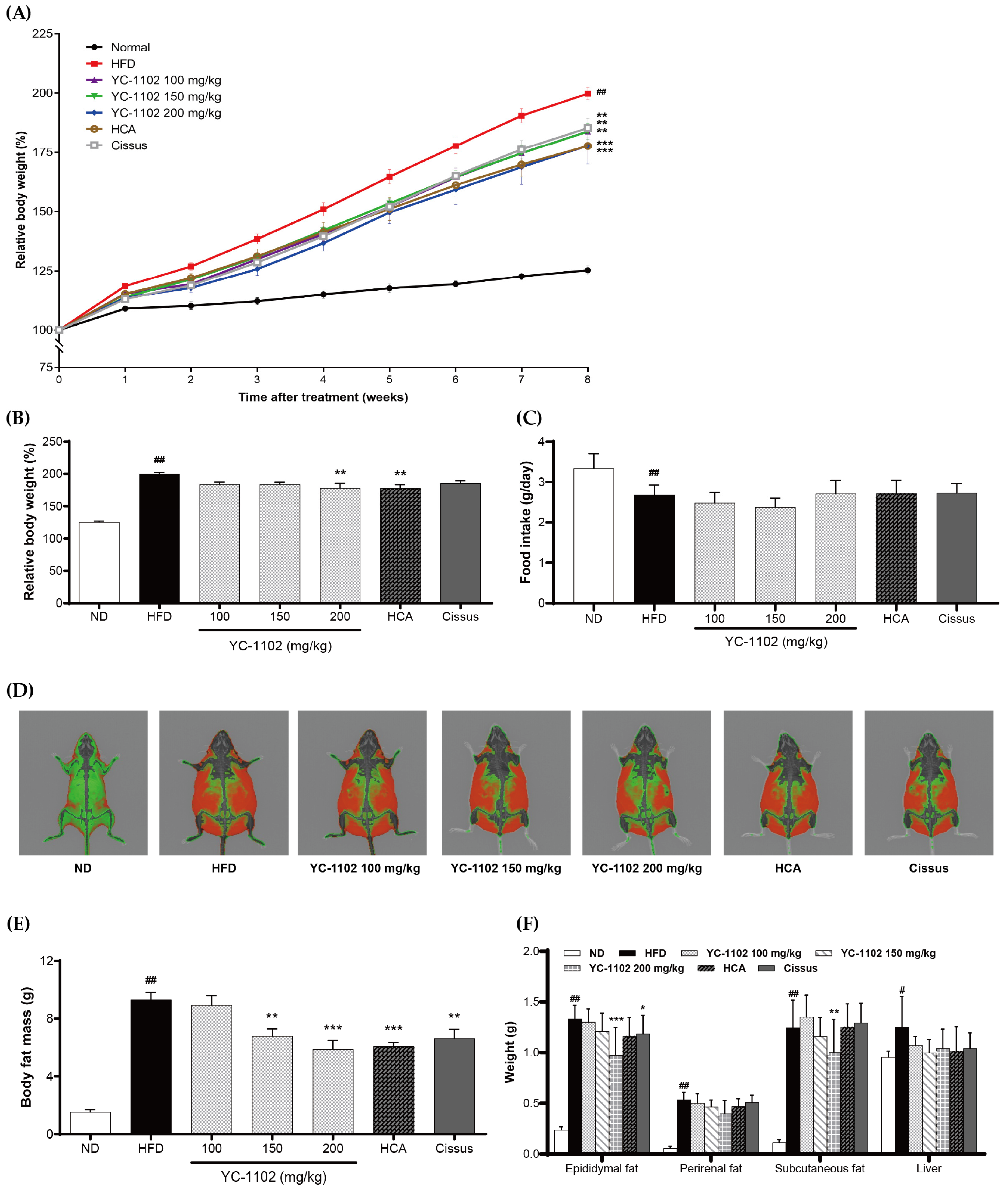
3.3. YC-1102 Reduces the Body Weight and Body Fat of HFD-Induced Obese Mice
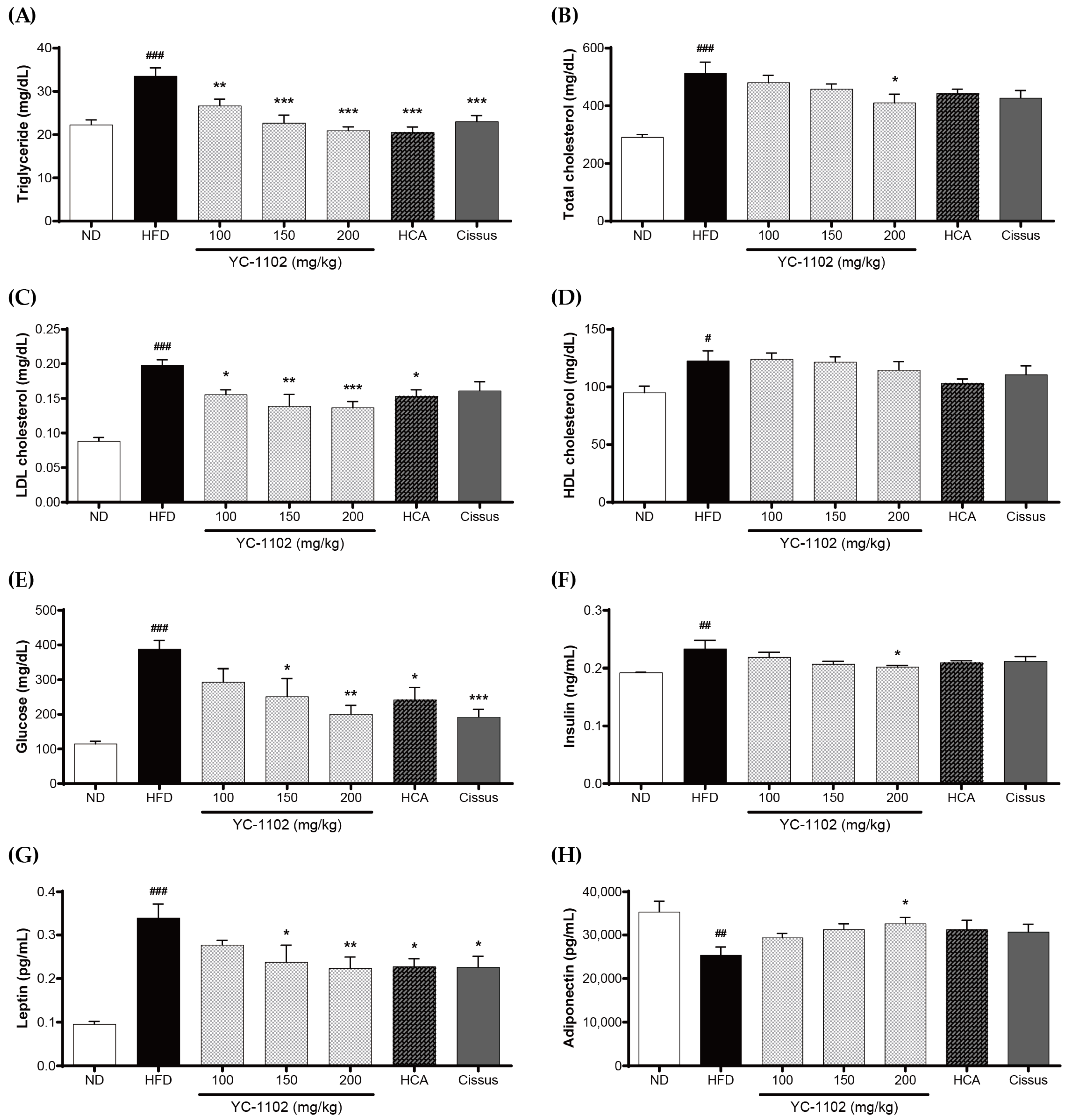
3.4. Effects of YC-1102 on Biochemical Parameters in the Serum of HFD-Induced Obese Mice
3.5. Effects of YC-1102 on eWAT Size and Gene Expressions Related to Adipogenesis and Thermogenesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef] [PubMed]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Otto, T.C.; Lane, M.D. Mitotic clonal expansion: A synchronous process required for adipogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Yudkin, J.S. Inflammation, obesity, and the metabolic syndrome. Horm. Metab. Res. 2007, 39, 707–709. [Google Scholar] [CrossRef] [PubMed]
- Calle, E.E.; Rodriguez, C.; Walker-Thurmond, K.; Thun, M.J. Overweight, obesity, and mortality from cancer in a prospectively studied cohort of U.S. adults. N. Engl. J. Med. 2003, 348, 1625–1638. [Google Scholar] [CrossRef] [PubMed]
- Botchlett, R.; Woo, S.L.; Liu, M.; Pei, Y.; Guo, X.; Li, H.; Wu, C. Nutritional approaches for managing obesity-associated metabolic diseases. J. Endocrinol. 2017, 233, R145–R171. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Han, J.H.; Yu, K.H.; Hong, M.; Lee, S.Y.; Park, K.H.; Lee, S.U.; Kwon, T.H. Antioxidant and Anti-Obesity Activities of Polygonum cuspidatum Extract through Alleviation of Lipid Accumulation on 3T3-L1 Adipocytes. J. Microbiol. Biotechnol. 2020, 30, 21–30. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef]
- Hong, F.; Pan, S.; Guo, Y.; Xu, P.; Zhai, Y. PPARs as Nuclear Receptors for Nutrient and Energy Metabolism. Molecules 2019, 24, 2545. [Google Scholar] [CrossRef]
- Giralt, M.; Villarroya, F. White, brown, beige/brite: Different adipose cells for different functions? Endocrinology 2013, 154, 2992–3000. [Google Scholar] [CrossRef]
- Cheng, C.F.; Ku, H.C.; Lin, H. PGC-1alpha as a Pivotal Factor in Lipid and Metabolic Regulation. Int. J. Mol. Sci. 2018, 19, 3447. [Google Scholar] [CrossRef]
- Kim, H.L.; Park, J.; Jung, Y.; Ahn, K.S.; Um, J.Y. Platycodin D, a novel activator of AMP-activated protein kinase, attenuates obesity in db/db mice via regulation of adipogenesis and thermogenesis. Phytomedicine 2019, 52, 254–263. [Google Scholar] [CrossRef]
- Qiang, L.; Wang, L.; Kon, N.; Zhao, W.; Lee, S.; Zhang, Y.; Rosenbaum, M.; Zhao, Y.; Gu, W.; Farmer, S.R.; et al. Brown remodeling of white adipose tissue by SirT1-dependent deacetylation of Ppargamma. Cell 2012, 150, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.T.; Hochfeld, W.E.; Myburgh, R.; Pepper, M.S. Adipocyte and adipogenesis. Eur. J. Cell Biol. 2013, 92, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Van Gaal, L.; Pi-Sunyer, X.; Despres, J.P.; McCarthy, C.; Scheen, A. Efficacy and safety of rimonabant for improvement of multiple cardiometabolic risk factors in overweight/obese patients: Pooled 1-year data from the Rimonabant in Obesity (RIO) program. Diabetes Care 2008, 31 (Suppl. S2), S229–S240. [Google Scholar] [CrossRef] [PubMed]
- Pi-Sunyer, X.; Astrup, A.; Fujioka, K.; Greenway, F.; Halpern, A.; Krempf, M.; Lau, D.C.; le Roux, C.W.; Violante Ortiz, R.; Jensen, C.B.; et al. A randomized, controlled trial of 3.0 mg of liraglutide in weight management. N. Engl. J. Med. 2015, 373, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L. A new triterpenic acid from roots of Rosa multiflora var. cathayensis. Chin. Tradit. Herb. Drugs 2017, 48, 4208–4214. [Google Scholar]
- Park, K.H.; Kim, S.K.; Choi, S.E.; Kwon, J.H.; Oh, M.H.; Lee, M.W. Three new stereoisomers of condensed tannins from the roots of Rosa multiflora. Chem. Pharm. Bull. 2010, 58, 1227–1231. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhao, X.; Sun, Y.; Liu, H.; Shang, M.; Gong, J.; Ma, Q.; Piao, G.; Yuan, H. Inhibitory effects of compounds from the roots of Potentilla longifolia on lipid accumulation. PLoS ONE 2020, 15, e0238917. [Google Scholar] [CrossRef]
- Cheol Park, J.; Chul Kim, S.; Moon Hur, J.; Choi, S.H.; Yeon Lee, K.; Won Choi, J. Anti-hepatotoxic effects of Rosa rugosa root and its compound, rosamultin, in rats intoxicated with bromobenzene. J. Med. Food 2004, 7, 436–441. [Google Scholar] [CrossRef]
- Choi, J.H.; Baek, J.Y.; Choi, H.J. Effects of Rosa multiflora and Rosa multiflora complex on lipid content in rats fed a high-fat high-cholesterol diet. J. Korean Soc. Food Sci. Nutr. 2015, 44, 791–799. [Google Scholar] [CrossRef]
- Park, K.H.; Choi, S.E.; Choi, Y.W.; Lee, D.I.; Joo, S.S.; Jeong, M.S.; Bang, H.; Lee, C.S.; Lee, M.K.; Seo, S.J.; et al. Topical application of two condensed tannins from the root of Rosa multiflora Thunberg for the treatment of atopic dermatitis (AD) in NC/Nga mice. Phytother. Res. 2011, 25, 1564–1569. [Google Scholar] [CrossRef]
- Semwal, R.B.; Semwal, D.K.; Vermaak, I.; Viljoen, A. A comprehensive scientific overview of Garcinia cambogia. Fitoterapia 2015, 102, 134–148. [Google Scholar] [CrossRef] [PubMed]
- Spalding, K.L.; Arner, E.; Westermark, P.O.; Bernard, S.; Buchholz, B.A.; Bergmann, O.; Blomqvist, L.; Hoffstedt, J.; Naslund, E.; Britton, T.; et al. Dynamics of fat cell turnover in humans. Nature 2008, 453, 783–787. [Google Scholar] [CrossRef] [PubMed]
- Kusminski, C.M.; Bickel, P.E.; Scherer, P.E. Targeting adipose tissue in the treatment of obesity-associated diabetes. Nat. Rev. Drug Discov. 2016, 15, 639–660. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: Structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Payne, V.A.; Au, W.S.; Lowe, C.E.; Rahman, S.M.; Friedman, J.E.; O’Rahilly, S.; Rochford, J.J. C/EBP transcription factors regulate SREBP1c gene expression during adipogenesis. Biochem. J. 2009, 425, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, A.; Oh, K.J.; Lee, S.C.; Kim, W.K.; Bae, K.H. The role of adipose tissue mitochondria: Regulation of mitochondrial function for the treatment of metabolic diseases. Int. J. Mol. Sci. 2019, 20, 4924. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Wang, S.; Zhang, C.; Zhao, Y. Coordinated modulation of energy metabolism and inflammation by branched-chain amino acids and fatty acids. Front. Endocrinol. 2020, 11, 617. [Google Scholar] [CrossRef]
- Pan, R.; Zhu, X.; Maretich, P.; Chen, Y. Combating obesity with thermogenic fat: Current challenges and advancements. Front. Endocrinol. 2020, 11, 185. [Google Scholar] [CrossRef]
- de Mello, A.H.; Costa, A.B.; Engel, J.D.G.; Rezin, G.T. Mitochondrial dysfunction in obesity. Life Sci. 2018, 192, 26–32. [Google Scholar] [CrossRef]
- Serra, D.; Mera, P.; Malandrino, M.I.; Mir, J.F.; Herrero, L. Mitochondrial fatty acid oxidation in obesity. Antioxid. Redox Signal. 2013, 19, 269–284. [Google Scholar] [CrossRef]
- Fernandez-Marcos, P.J.; Auwerx, J. Regulation of PGC-1alpha, a nodal regulator of mitochondrial biogenesis. Am. J. Clin. Nutr. 2011, 93, 884S–890S. [Google Scholar] [CrossRef] [PubMed]
- Jornayvaz, F.R.; Shulman, G.I. Regulation of mitochondrial biogenesis. Essays Biochem. 2010, 47, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Busiello, R.A.; Savarese, S.; Lombardi, A. Mitochondrial uncoupling proteins and energy metabolism. Front. Physiol. 2015, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Lancha, A.; Fruhbeck, G.; Gomez-Ambrosi, J. Peripheral signalling involved in energy homeostasis control. Nutr. Res. Rev. 2012, 25, 223–248. [Google Scholar] [CrossRef] [PubMed]
- Bluher, M.; Mantzoros, C.S. From leptin to other adipokines in health and disease: Facts and expectations at the beginning of the 21st century. Metabolism 2015, 64, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Neuhofer, A.; Zeyda, M.; Mascher, D.; Itariu, B.K.; Murano, I.; Leitner, L.; Hochbrugger, E.E.; Fraisl, P.; Cinti, S.; Serhan, C.N.; et al. Impaired local production of proresolving lipid mediators in obesity and 17-HDHA as a potential treatment for obesity-associated inflammation. Diabetes 2013, 62, 1945–1956. [Google Scholar] [CrossRef] [PubMed]
- Bluher, M.; Michael, M.D.; Peroni, O.D.; Ueki, K.; Carter, N.; Kahn, B.B.; Kahn, C.R. Adipose tissue selective insulin receptor knockout protects against obesity and obesity-related glucose intolerance. Dev. Cell 2002, 3, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Yu, Y.H.; Shew, J.Y.; Lee, W.J.; Hwang, J.J.; Chen, Y.H.; Chen, Y.R.; Wei, P.C.; Chuang, L.M.; Lee, W.H. Deficiency of NPGPx, an oxidative stress sensor, leads to obesity in mice and human. EMBO Mol. Med. 2013, 5, 1165–1179. [Google Scholar] [CrossRef]
- Spalding, K.L.; Bernard, S.; Naslund, E.; Salehpour, M.; Possnert, G.; Appelsved, L.; Fu, K.Y.; Alkass, K.; Druid, H.; Thorell, A.; et al. Impact of fat mass and distribution on lipid turnover in human adipose tissue. Nat. Commun. 2017, 8, 15253. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.; McGee, S.L. 3T3-L1 adipocytes display phenotypic characteristics of multiple adipocyte lineages. Adipocyte 2015, 4, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Ringholm, S.; Grunnet Knudsen, J.; Leick, L.; Lundgaard, A.; Munk Nielsen, M.; Pilegaard, H. PGC-1alpha is required for exercise- and exercise training-induced UCP1 up-regulation in mouse white adipose tissue. PLoS ONE 2013, 8, e64123. [Google Scholar] [CrossRef]
- Myers, M.G.; Cowley, M.A.; Munzberg, H. Mechanisms of leptin action and leptin resistance. Annu. Rev. Physiol. 2008, 70, 537–556. [Google Scholar] [CrossRef] [PubMed]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A novel serum protein similar to C1q, produced exclusively in adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef]
- Scheid, M.P.; Sweeney, G. The role of adiponectin signaling in metabolic syndrome and cancer. Rev. Endocr. Metab. Disord. 2014, 15, 157–167. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamauchi, T.; Kubota, N.; Hara, K.; Ueki, K.; Tobe, K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Investig. 2006, 116, 1784–1792. [Google Scholar] [CrossRef]
- Park, H.J.; Nam, J.H.; Jung, H.J.; Lee, M.S.; Lee, K.T.; Jung, M.H.; Choi, J. Inhibitory effect of euscaphic acid and and tormentic acid from the roots of Rosa rugosa on high fat diet-induced obesity in the rat. Korean J. Pharmacogn. 2005, 36, 324–331. [Google Scholar]
- Feng, S.; Dai, Z.; Liu, A.B.; Huang, J.; Narsipur, N.; Guo, G.; Kong, B.; Reuhl, K.; Lu, W.; Luo, Z.; et al. Intake of stigmasterol and beta-sitosterol alters lipid metabolism and alleviates NAFLD in mice fed a high-fat western-style diet. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 1274–1284. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequences (5′ → 3′) |
|---|---|---|
| C/EBPα | Forward | GCGGGAACGCAACAACATC |
| Reverse | GTCACTGGTCAACTCCAGCAC | |
| PPARγ | Forward | TTCGCTGATGCACTGCCTATGA |
| Reverse | AAGGAATGCGAGTGGTCTTCCA | |
| PGC-1α | Forward | TGTTCCCGATCACCATATTCC |
| Reverse | GGTGTCTGTAGTGGCTTGATTC | |
| UCP-1 | Forward | GTGAACCCGACAACTTCCGAA |
| Reverse | TGAAACTCCGGCTGAGAAGAT | |
| GAPDH | Forward | AGGTCTGGTGTGAACGGATTTG |
| Reverse | TGTAGACCATGTAGTTGAGGTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.-Y.; Kim, K.K.; Baek, S.H.; Park, C.I.; Jeon, H.J.; Song, A.R.; Park, H.-J.; Park, I.B.; Kang, J.S.; Kim, J.M.; et al. Effect of YC-1102 on the Improvement of Obesity in High-Fat Diet-Induced Obese Mice. Curr. Issues Mol. Biol. 2024, 46, 1437-1450. https://doi.org/10.3390/cimb46020093
Yu H-Y, Kim KK, Baek SH, Park CI, Jeon HJ, Song AR, Park H-J, Park IB, Kang JS, Kim JM, et al. Effect of YC-1102 on the Improvement of Obesity in High-Fat Diet-Induced Obese Mice. Current Issues in Molecular Biology. 2024; 46(2):1437-1450. https://doi.org/10.3390/cimb46020093
Chicago/Turabian StyleYu, Hwa-Young, Kyoung Kon Kim, Sin Hwa Baek, Cho I Park, Hye Jin Jeon, Ae Ri Song, Hyun-Je Park, Il Bum Park, Jong Soo Kang, Jung Min Kim, and et al. 2024. "Effect of YC-1102 on the Improvement of Obesity in High-Fat Diet-Induced Obese Mice" Current Issues in Molecular Biology 46, no. 2: 1437-1450. https://doi.org/10.3390/cimb46020093
APA StyleYu, H. -Y., Kim, K. K., Baek, S. H., Park, C. I., Jeon, H. J., Song, A. R., Park, H. -J., Park, I. B., Kang, J. S., Kim, J. M., Kim, T. W., Jang, S. M., Cha, J. Y., & Kim, J. (2024). Effect of YC-1102 on the Improvement of Obesity in High-Fat Diet-Induced Obese Mice. Current Issues in Molecular Biology, 46(2), 1437-1450. https://doi.org/10.3390/cimb46020093