
The Role of Aminopeptidase ERAP1 in Human Pathology—A Review

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
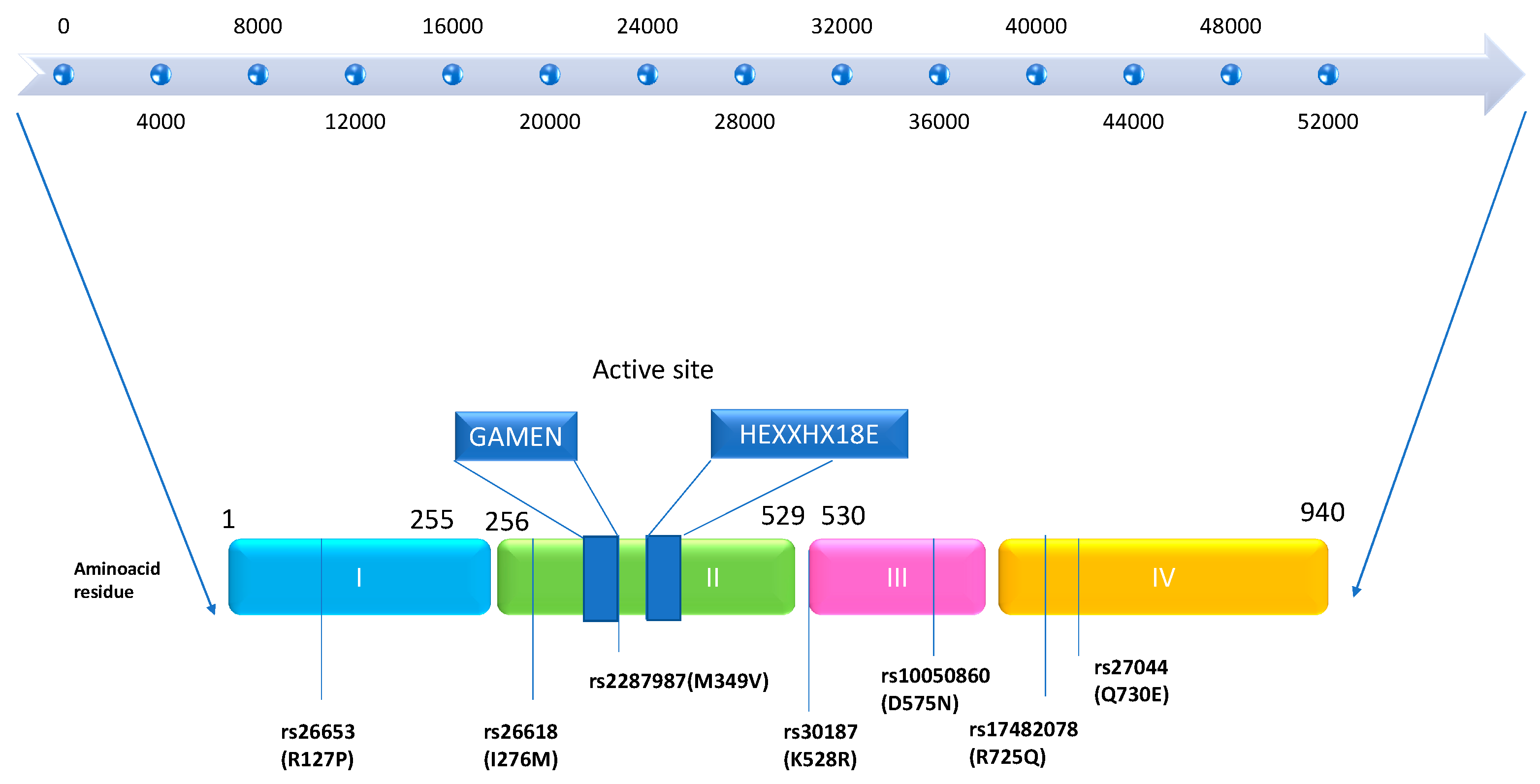
2. Genetics
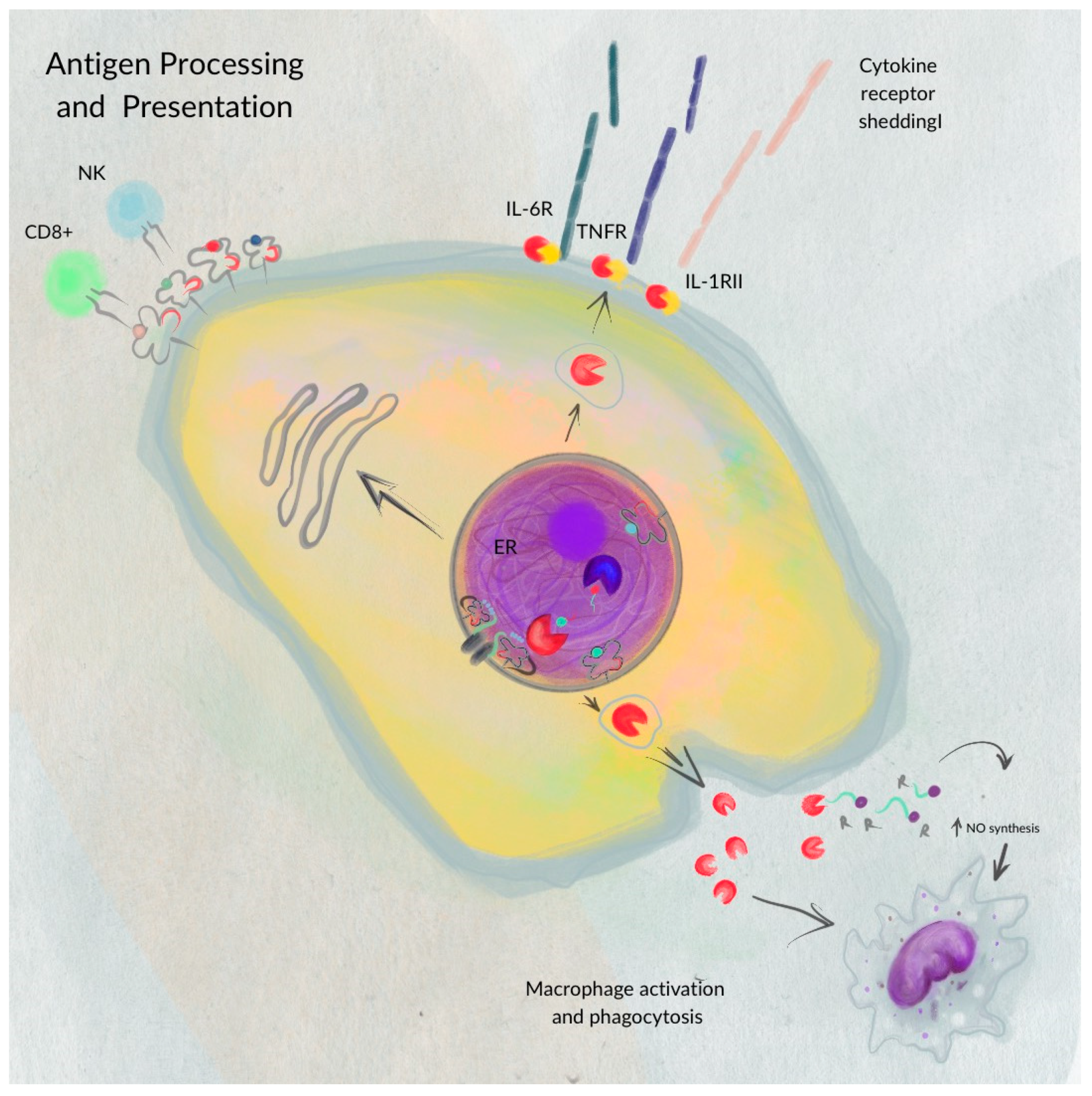
3. Function of ERAP1 and ERAP2
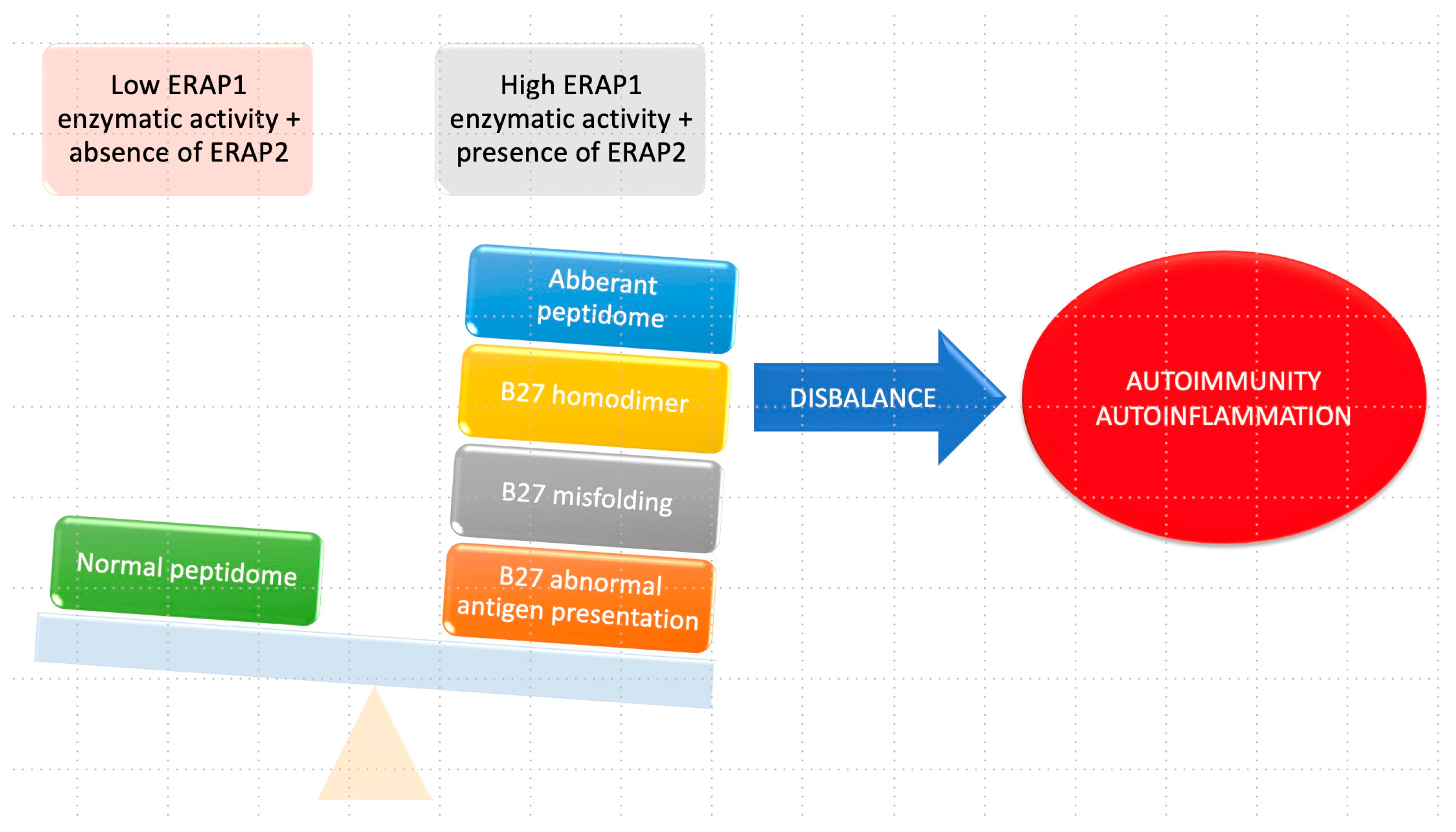
4. Role of ERAP1 in Human Pathogenesis
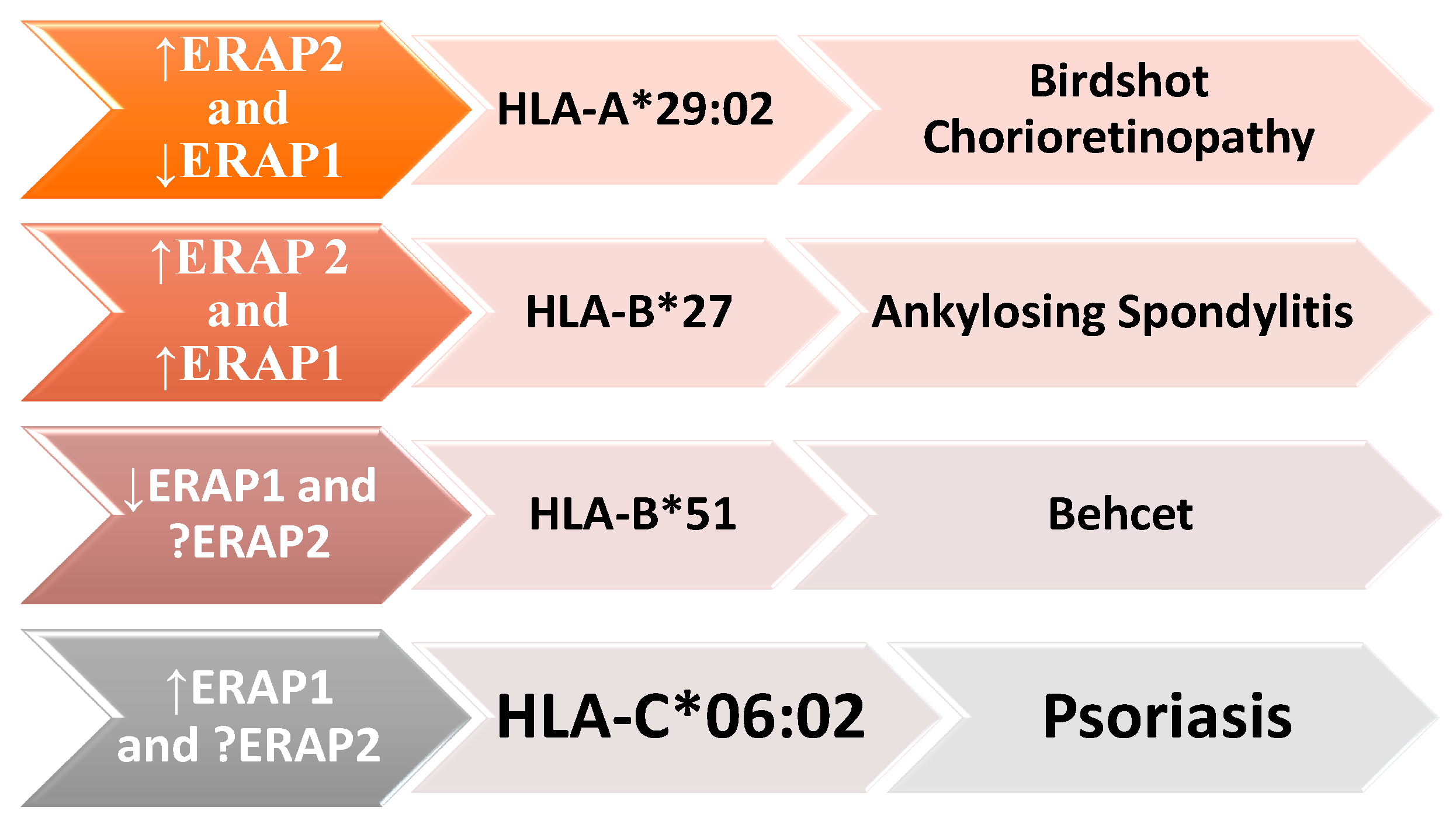
4.1. Autoimmune and Autoinflammatory Diseases
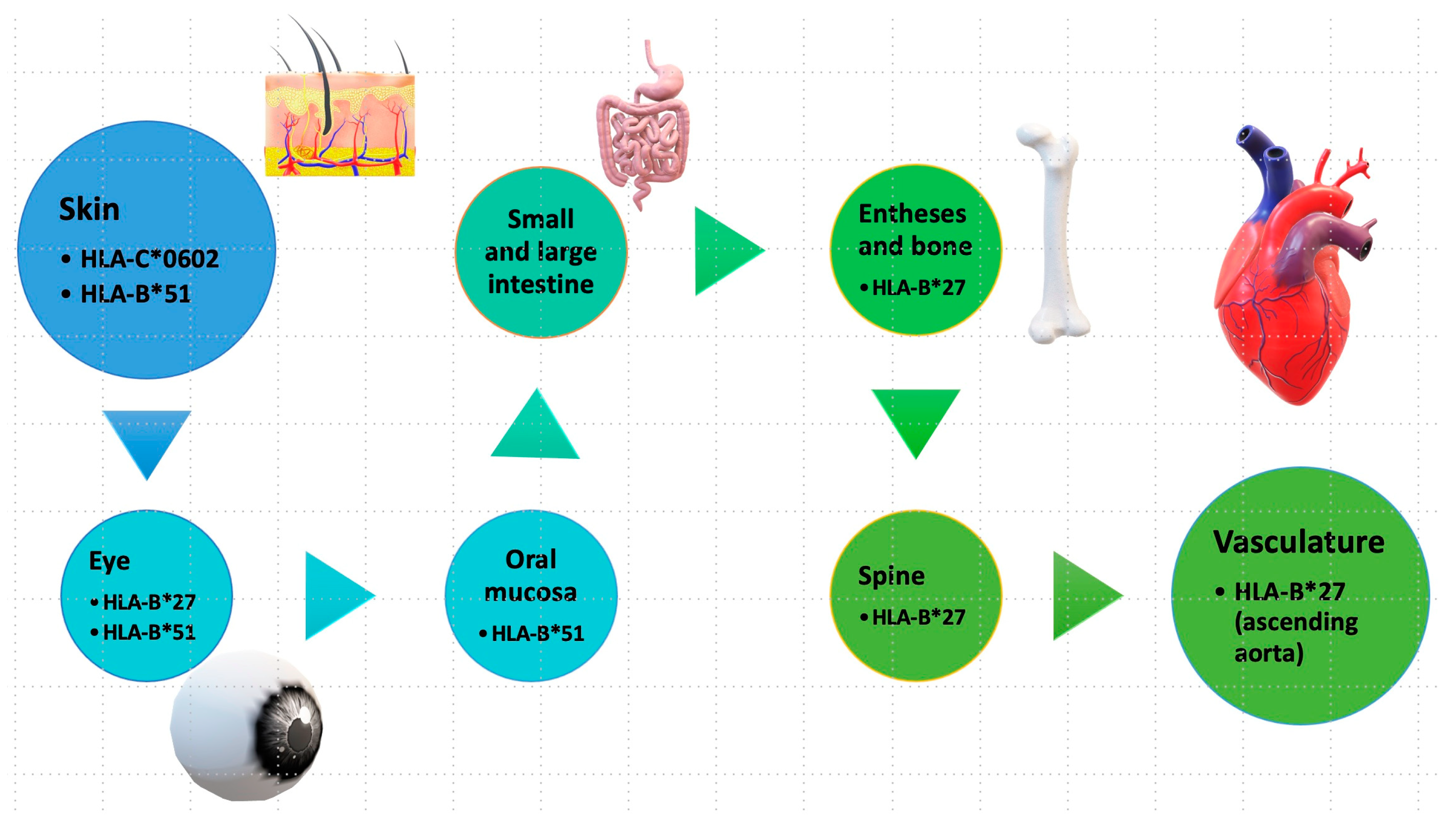
4.1.1. Ankylosing Spondylitis (AS)
4.1.2. Behçet’s Disease (BD)
4.1.3. Birdshot Chorioretinopathy (BSCR)
4.1.4. Type 1 Diabetes Mellitus (DM1)
4.1.5. Psoriasis (Ps)
4.2. Role in Cancer
- Low expression of both enzymes is the most common cause of tumor localization, regardless of its location.
- Upregulation (tissues with undetectable levels of ERAP1 and ERAP2) is observed in thyroid and colon cancers.
4.3. Atopic Dermatitis
4.4. COVID Infection
4.5. Arterial Hypertension (HTA)
5. Conclusions and Future Perspectives
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mucha, A.; Drag, M.; Dalton, J.P.; Kafarski, P. Metallo-Aminopeptidase Inhibitors. Biochimie 2010, 92, 1509–1529. [Google Scholar] [CrossRef]
- Zhu, W.; He, X.; Cheng, K.; Zhang, L.; Chen, D.; Wang, X.; Qiu, G.; Cao, X.; Weng, X. Ankylosing Spondylitis: Etiology, Pathogenesis, and Treatments. Bone Res. 2019, 7, 22. [Google Scholar] [CrossRef]
- Paladini, F.; Fiorillo, M.T.; Tedeschi, V.; Mattorre, B.; Sorrentino, R. The Multifaceted Nature of Aminopeptidases ERAP1, ERAP2, and LNPEP: From Evolution to Disease. Front. Immunol. 2020, 11, 1576. [Google Scholar] [CrossRef]
- Wellcome Trust Case Control Consortium; Donnelly, P.; Barrett, J.C.; Davison, D.; Easton, D.; Evans, D.M.; Leung, H.-T.; Marchini, J.L.; Morris, A.P.; Spencer, C.C.A. Association Scan of 14,500 Nonsynonymous SNPs in Four Diseases Identifies Autoimmunity Variants. Nat. Genet 2007, 39, 1329–1337. [Google Scholar]
- Nguyen, T.T.; Chang, S.-C.; Evnouchidou, I.; York, I.A.; Zikos, C.; Rock, K.L.; Goldberg, A.L.; Stratikos, E.; Stern, L.J. Structural Basis for Antigenic Peptide Precursor Processing by the Endoplasmic Reticulum Aminopeptidase ERAP1. Nat. Struct. Mol. Biol. 2011, 18, 604–613. [Google Scholar] [CrossRef]
- Keidel, S.; Chen, L.; Pointon, J.; Wordsworth, P. ERAP1 and Ankylosing Spondylitis. Curr. Opin. Immunol. 2013, 25, 97–102. [Google Scholar] [CrossRef]
- Reeves, E.; James, E. The Role of Polymorphic ERAP1 in Autoinflammatory Disease. Biosci. Rep. 2018, 38, BSR20171503. [Google Scholar] [CrossRef]
- López de Castro, J.A. How ERAP1 and ERAP2 Shape the Peptidomes of Disease-Associated MHC-I Proteins. Front. Immunol. 2018, 9, 2463. [Google Scholar] [CrossRef] [PubMed]
- Kadi, A.; Izac, B.; Said-Nahal, R.; Leboime, A.; Van Praet, L.; de Vlam, K.; Elewaut, D.; Chiocchia, G.; Breban, M. Investigating the Genetic Association between ERAP1 and Spondyloarthritis. Ann. Rheum. Dis. 2013, 72, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Compagnone, M.; Cifaldi, L.; Fruci, D. Regulation of ERAP1 and ERAP2 Genes and Their Disfunction in Human Cancer. Hum. Immunol. 2019, 80, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Saulle, I.; Vicentini, C.; Clerici, M.; Biasin, M. Antigen Presentation in SARS-CoV-2 Infection: The Role of Class I HLA and ERAP Polymorphisms. Hum. Immunol. 2021, 82, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Cherciu, M.; Popa, L.O.; Bojinca, M.; Dutescu, M.I.; Bojinca, V.; Bara, C.; Popa, O.M. Functional Variants of ERAP1 Gene Are Associated with HLA-B27 Positive Spondyloarthritis: ERAP1gene SNPs Are Associated with Spondyloarthritis in HLA-B27+ Romanians. Tissue Antigens 2013, 82, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Piekarska, K.; Radwan, P.; Tarnowska, A.; Wiśniewski, A.; Radwan, M.; Wilczyński, J.R.; Malinowski, A.; Nowak, I. ERAP, KIR, and HLA-C Profile in Recurrent Implantation Failure. Front. Immunol. 2021, 12, 755624. [Google Scholar] [CrossRef] [PubMed]
- Serwold, T.; Gonzalez, F.; Kim, J.; Jacob, R.; Shastri, N. ERAAP Customizes Peptides for MHC Class I Molecules in the Endoplasmic Reticulum. Nature 2002, 419, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Hawari, F.; Alsaaty, S.; Lawrence, M.; Combs, C.A.; Geng, W.; Rouhani, F.N.; Miskinis, D.; Levine, S.J. Identification of ARTS-1 as a Novel TNFR1-Binding Protein That Promotes TNFR1 Ectodomain Shedding. J. Clin. Investig. 2002, 110, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Benigni, A.; Cassis, P.; Remuzzi, G. Angiotensin II Revisited: New Roles in Inflammation, Immunology and Aging: Angiotensin II in Inflammation, Immunology, Aging. EMBO Mol. Med. 2010, 2, 247–257. [Google Scholar] [CrossRef]
- Yang, S.; Liu, X.; Gao, Y.; Ding, M.; Li, B.; Sun, H.; He, Y.; Yang, P. Association of Single Nucleotide Polymorphisms in the 3′UTR of ERAP1 Gene with Essential Hypertension in the Northeastern Han Chinese. Gene 2015, 560, 211–216. [Google Scholar] [CrossRef]
- Cooper, G.S.; Stroehla, B.C. The Epidemiology of Autoimmune Diseases. Autoimmun. Rev. 2003, 2, 119–125. [Google Scholar] [CrossRef]
- Liu, S.; Lu, J.; Wu, J.; Feng, D.; Wang, Y.; Su, X.; Cao, H. Structural and Biochemical Insights into the Association between ERAP1 Polymorphism and Autoimmune Diseases. Biochem. Biophys. Res. Commun. 2022, 632, 189–194. [Google Scholar] [CrossRef]
- Damian, L.; Spârchez, M.; Lupșe, M.; Felea, I.; Rednic, S.; Pamfil, C.; Bucșa, C.; Vulturar, R. Sindroame autoinflamatorii granulomatoase associate Cu NOD2—o Scurtă Actualizare Pentru Clinicieni. Romanian J. Pediatr. 2021, LXX, 179–185. [Google Scholar] [CrossRef]
- McGonagle, D.; McDermott, M.F. A Proposed Classification of the Immunological Diseases. PLoS Med. 2006, 3, e297. [Google Scholar] [CrossRef]
- Reeves, E.; Colebatch-Bourn, A.; Elliott, T.; Edwards, C.J.; James, E. Functionally Distinct ERAP1 Allotype Combinations Distinguish Individuals with Ankylosing Spondylitis. Proc. Natl. Acad. Sci. USA 2014, 111, 17594–17599. [Google Scholar] [CrossRef]
- Zenewicz, L.A.; Abraham, C.; Flavell, R.A.; Cho, J.H. Unraveling the Genetics of Autoimmunity. Cell 2010, 140, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Matzaraki, V.; Kumar, V.; Wijmenga, C.; Zhernakova, A. The MHC Locus and Genetic Susceptibility to Autoimmune and Infectious Diseases. Genome Biol. 2017, 18, 76. [Google Scholar] [CrossRef] [PubMed]
- Guerini, F.R.; Cagliani, R.; Forni, D.; Agliardi, C.; Caputo, D.; Cassinotti, A.; Galimberti, D.; Fenoglio, C.; Biasin, M.; Asselta, R.; et al. A Functional Variant in ERAP1 Predisposes to Multiple Sclerosis. PLoS ONE 2012, 7, e29931. [Google Scholar] [CrossRef] [PubMed]
- Paladini, F.; Fiorillo, M.T.; Tedeschi, V.; Cauli, A.; Mathieu, A.; Sorrentino, R. Ankylosing Spondylitis: A Trade off HLA-B27, ERAP, and Pathogen Interconnections? Focus on Sardinia. Front. Immunol. 2019, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- McGonagle, D.; Aydin, S.Z.; Gül, A.; Mahr, A.; Direskeneli, H. ‘MHC-I-Opathy’—Unified Concept for Spondyloarthritis and Behçet Disease. Nat. Rev. Rheumatol. 2015, 11, 731–740. [Google Scholar] [CrossRef]
- Evans, D.M.; Spencer, C.C.A.; Pointon, J.J.; Su, Z.; Harvey, D.; Kochan, G.; Oppermann, U.; Dilthey, A.; Pirinen, M.; Stone, M.A.; et al. Interaction between ERAP1 and HLA-B27 in Ankylosing Spondylitis Implicates Peptide Handling in the Mechanism for HLA-B27 in Disease Susceptibility. Nat. Genet. 2011, 43, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.M.; Colbert, R.A. Endoplasmic Reticulum Aminopeptidase 1 and Rheumatic Disease: Functional Variation. Curr. Opin. Rheumatol. 2015, 27, 357–363. [Google Scholar] [CrossRef]
- Brown, M.A.; Kennedy, L.G.; Macgregor, A.J.; Darke, C.; Duncan, E.; Shatford, J.L.; Taylor, A.; Calin, A.; Wordsworth, P. Susceptibility to Ankylosing Spondylitis in Twins the Role of Genes, HLA, and the Environment. Arthritis Rheum. 1997, 40, 1823–1828. [Google Scholar] [CrossRef]
- Caffrey, M.F.; James, D.C. Human Lymphocyte Antigen Association in Ankylosing Spondylitis. Nature 1973, 242, 121. [Google Scholar] [CrossRef]
- Vitulano, C.; Tedeschi, V.; Paladini, F.; Sorrentino, R.; Fiorillo, M.T. The Interplay between HLA-B27 and ERAP1/ERAP2 Aminopeptidases: From Anti-Viral Protection to Spondyloarthritis. Clin. Exp. Immunol. 2017, 190, 281–290. [Google Scholar] [CrossRef]
- Sharip, A.; Kunz, J. Understanding the Pathogenesis of Spondyloarthritis. Biomolecules 2020, 10, 1461. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-M.; Ho, H.-H.; Chang, S.-W.; Wu, Y.-J.J.; Lin, J.-C.; Chang, P.-Y.; Wu, J.; Chen, J.-Y. ERAP1 Genetic Variations Associated with HLA-B27 Interaction and Disease Severity of Syndesmophytes Formation in Taiwanese Ankylosing Spondylitis. Arthritis Res. Ther. 2012, 14, R125. [Google Scholar] [CrossRef] [PubMed]
- Colbert, R.A.; DeLay, M.L.; Klenk, E.I.; Layh-Schmitt, G. From HLA-B27 to Spondyloarthritis: A Journey through the ER. Immunol. Rev. 2010, 233, 181–202. [Google Scholar] [CrossRef] [PubMed]
- Rudwaleit, M.; Haibel, H.; Baraliakos, X.; Listing, J.; Märker-Hermann, E.; Zeidler, H.; Braun, J.; Sieper, J. The Early Disease Stage in Axial Spondylarthritis: Results from the German Spondyloarthritis Inception Cohort. Arthritis Rheum. 2009, 60, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Gallego, C.; Aydin, S.Z.; Emery, P.; McGonagle, D.G.; Marzo-Ortega, H. Magnetic Resonance Imaging Assessment of Axial Psoriatic Arthritis: Extent of Disease Relates to HLA-B27: HLA-B27 Status and Axial PsA Findings on MRI. Arthritis Rheum. 2013, 65, 2274–2278. [Google Scholar] [CrossRef] [PubMed]
- Costantino, F.; Breban, M.; Garchon, H.-J. Genetics and Functional Genomics of Spondyloarthritis. Front. Immunol. 2018, 9, 2933. [Google Scholar] [CrossRef]
- Ombrello, M.J.; Kastner, D.L.; Remmers, E.F. Endoplasmic Reticulum-Associated Aminopeptidase 1, and Rheumatic Disease: Genetics. Curr. Opin. Rheumatol. 2015, 27, 349–356. [Google Scholar] [CrossRef]
- Kochan, G.; Krojer, T.; Harvey, D.; Fischer, R.; Chen, L.; Vollmar, M.; Delft, F.; Kavanagh, K.L.; Brown, M.A.; Bowness, P.; et al. Crystal Structures of the Endoplasmic Reticulum Aminopeptidase-1 (ERAP1) Reveal the Molecular Basis for N-Terminal Peptide Trimming. Proc. Natl. Acad. Sci. USA 2011, 108, 7745–7750. [Google Scholar] [CrossRef]
- Costantino, F.; Talpin, A.; Evnouchidou, I.; Kadi, A.; Leboime, A.; Said-Nahal, R.; Bonilla, N.; Letourneur, F.; Leturcq, T.; Ka, Z.; et al. ERAP1 Gene Expression Is Influenced by Nonsynonymous Polymorphisms Associated with Predisposition to Spondyloarthritis. Arthritis Rheumatol. 2015, 67, 1525–1534. [Google Scholar] [CrossRef]
- Díaz-Peña, R.; Castro-Santos, P.; Durán, J.; Santiago, C.; Lucia, A. The Genetics of Spondyloarthritis. J. Pers. Med. 2020, 10, 151. [Google Scholar] [CrossRef]
- Bremander, A.; Petersson, I.F.; Bergman, S.; Englund, M. Population-Based Estimates of Common Comorbidities and Cardiovascular Disease in Ankylosing Spondylitis. Arthritis Care Res. 2011, 63, 550–556. [Google Scholar] [CrossRef]
- Tsui, F.W.L.; Haroon, N.; Reveille, J.D.; Rahman, P.; Chiu, B.; Tsui, H.W.; Inman, R.D. Association of an ERAP1 ERAP2 Haplotype with Familial Ankylosing Spondylitis. Ann. Rheum. Dis. 2010, 69, 733–736. [Google Scholar] [CrossRef]
- Tang, Y.; Yang, P.; Wang, F.; Xu, H.; Zong, S.-Y. Association of Polymorphisms in ERAP1 and Risk of Ankylosing Spondylitis in a Chinese Population. Gene 2018, 646, 8–11. [Google Scholar] [CrossRef]
- Ozen, G.; Deniz, R.; Eren, F.; Erzik, C.; Unal, A.U.; Yavuz, S.; Aydin, S.Z.; Inanc, N.; Direskeneli, H.; Atagunduz, P. Association of ERAP1, IL23R and PTGER4 Polymorphisms with Radiographic Severity of Ankylosing Spondylitis. Open Rheumatol. J. 2017, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, H.; Babaie, F.; Hemmatzadeh, M.; Azizi, G.; Hajaliloo, M.; Ebrahimi, A.A.; Kazemi, T.; Yousefi, M.; Rezaiemanesh, A.; Safarzadeh, E.; et al. Evaluation of ERAP1 Gene Single Nucleotide Polymorphism in Impressing the Inflammatory Cytokine Profile of Ankylosing Spondylitis Patients. Iran. J. Allergy Asthma Immunol. 2018, 17, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Harvey, D.; Pointon, J.J.; Evans, D.M.; Karaderi, T.; Farrar, C.; Appleton, L.H.; Sturrock, R.D.; Stone, M.A.; Oppermann, U.; Brown, M.A.; et al. Investigating the Genetic Association between ERAP1 and Ankylosing Spondylitis. Hum. Mol. Genet. 2009, 18, 4204–4212. [Google Scholar] [CrossRef] [PubMed]
- Haroon, N. Endoplasmic Reticulum Aminopeptidase 1 and Interleukin-23 Receptor in Ankylosing Spondylitis. Curr. Rheumatol. Rep. 2012, 14, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.A.; Abdul-Sattar, A.B.; Abdelal, I.T.; Baraka, A. Shedding Light on the Role of ERAP1 in Axial Spondyloarthritis. Cureus 2023, 15, e48806. [Google Scholar] [CrossRef]
- Mattioli, I.; Bettiol, A.; Saruhan-Direskeneli, G.; Direskeneli, H.; Emmi, G. Pathogenesis of Behçet’s Syndrome: Genetic, Environmental and Immunological Factors. Front. Med. 2021, 8, 713052. [Google Scholar] [CrossRef]
- Davatchi, F.; Chams-Davatchi, C.; Shams, H.; Shahram, F.; Nadji, A.; Akhlaghi, M.; Faezi, T.; Ghodsi, Z.; Sadeghi Abdollahi, B.; Ashofteh, F.; et al. Behcet’s Disease: Epidemiology, Clinical Manifestations, and Diagnosis. Expert Rev. Clin. Immunol. 2017, 13, 57–65. [Google Scholar] [CrossRef]
- Kirino, Y.; Bertsias, G.; Ishigatsubo, Y.; Mizuki, N.; Tugal-Tutkun, I.; Seyahi, E.; Ozyazgan, Y.; Sacli, F.S.; Erer, B.; Inoko, H.; et al. Genome-Wide Association Analysis Identifies New Susceptibility Loci for Behçet’s Disease and Epistasis between HLA-B*51 and ERAP1. Nat. Genet. 2013, 45, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Sousa, I.; Shahram, F.; Francisco, D.; Davatchi, F.; Abdollahi, B.S.; Ghaderibarmi, F.; Nadji, A.; Mojarad Shafiee, N.; Xavier, J.M.; Oliveira, S.A. Brief Report: Association of CCR1, KLRC4, IL12A-AS1, STAT4, and ERAP1 With Behçet’s Disease in Iranians: Susceptibility Genes for BD. Arthritis Rheumatol. 2015, 67, 2742–2748. [Google Scholar] [CrossRef] [PubMed]
- Guasp, P.; Barnea, E.; González-Escribano, M.F.; Jiménez-Reinoso, A.; Regueiro, J.R.; Admon, A.; López de Castro, J.A. The Behçet’s Disease-Associated Variant of the Aminopeptidase ERAP1 Shapes a Low-Affinity HLA-B*51 Peptidome by Differential Subpeptidome Processing. J. Biol. Chem. 2017, 292, 9680–9689. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Ombrello, M.J.; Kirino, Y.; Erer, B.; Tugal-Tutkun, I.; Seyahi, E.; Özyazgan, Y.; Watts, N.R.; Gül, A.; Kastner, D.L.; et al. A Single Endoplasmic Reticulum Aminopeptidase-1 Protein Allotype Is a Strong Risk Factor for Behçet’s Disease in HLA-B*51 Carriers. Ann. Rheum. Dis. 2016, 75, 2208–2211. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.H.; Levinson, R.D.; Yu, F.; Goldhardt, R.; Gordon, L.K.; Gonzales, C.R.; Heckenlively, J.R.; Kappel, P.J.; Holland, G.N. Birdshot Chorioretinopathy. Surv. Ophthalmol. 2005, 50, 519–541. [Google Scholar] [CrossRef] [PubMed]
- Brézin, A.P.; Monnet, D.; Cohen, J.H.M.; Levinson, R.D. HLA-A29 and Birdshot Chorioretinopathy. Ocul. Immunol. Inflamm. 2011, 19, 397–400. [Google Scholar] [CrossRef]
- Alvarez-Navarro, C.; Martín-Esteban, A.; Barnea, E.; Admon, A.; López de Castro, J.A. Endoplasmic Reticulum Aminopeptidase 1 (ERAP1) Polymorphism Relevant to Inflammatory Disease Shapes the Peptidome of the Birdshot Chorioretinopathy-Associated HLA-A*29:02 Antigen. Mol. Cell. Proteom. 2015, 14, 1770–1780. [Google Scholar] [CrossRef]
- Kuiper, J.J.W.; Van Setten, J.; Ripke, S.; Van’T Slot, R.; Mulder, F.; Missotten, T.; Baarsma, G.S.; Francioli, L.C.; Pulit, S.L.; De Kovel, C.G.F.; et al. A Genome-Wide Association Study Identifies a Functional ERAP2 Haplotype Associated with Birdshot Chorioretinopathy. Hum. Mol. Genet. 2014, 23, 6081–6087. [Google Scholar] [CrossRef]
- Kronenberg, D.; Knight, R.R.; Estorninho, M.; Ellis, R.J.; Kester, M.G.; de Ru, A.; Eichmann, M.; Huang, G.C.; Powrie, J.; Dayan, C.M.; et al. Circulating Preproinsulin Signal Peptide-Specific CD8 T Cells Restricted by the Susceptibility Molecule HLA-A24 Are Expanded at Onset of Type 1 Diabetes and Kill β-Cells. Diabetes 2012, 61, 1752–1759. [Google Scholar] [CrossRef]
- Kronenberg-Versteeg, D.; Eichmann, M.; Russell, M.A.; de Ru, A.; Hehn, B.; Yusuf, N.; van Veelen, P.A.; Richardson, S.J.; Morgan, N.G.; Lemberg, M.K.; et al. Molecular Pathways for Immune Recognition of Preproinsulin Signal Peptide in Type 1 Diabetes. Diabetes 2018, 67, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Fung, E.Y.M.G.; Smyth, D.J.; Howson, J.M.M.; Cooper, J.D.; Walker, N.M.; Stevens, H.; Wicker, L.S.; Todd, J.A. Analysis of 17 Autoimmune Disease-Associated Variants in Type 1 Diabetes Identifies 6q23/TNFAIP3 as a Susceptibility Locus. Genes Immun. 2009, 10, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Paldino, G.; Fierabracci, A. Shedding New Light on the Role of ERAP1 in Type 1 Diabetes: A Perspective on Disease Management. Autoimmun. Rev. 2023, 22, 103291. [Google Scholar] [CrossRef]
- Thomaidou, S.; Kracht, M.J.L.; van der Slik, A.; Laban, S.; de Koning, E.J.; Carlotti, F.; Hoeben, R.C.; Roep, B.O.; Zaldumbide, A. Β-Cell Stress Shapes CTL Immune Recognition of Preproinsulin Signal Peptide by Posttranscriptional Regulation of Endoplasmic Reticulum Aminopeptidase 1. Diabetes 2020, 69, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Bu, J.; Ding, R.; Zhou, L.; Chen, X.; Shen, E. Epidemiology of psoriasis and comorbid diseases: A narrative review. Front. Immunology. 2022, 13, 880201. [Google Scholar] [CrossRef]
- Wu, X.; Zhao, Z. Associations between ERAP1 Gene Polymorphisms and Psoriasis Susceptibility: A Meta-Analysis of Case-Control Studies. Biomed Res. Int. 2021, 2021, 5515868. [Google Scholar] [CrossRef]
- Trembath, R.C.; Lee Clough, R.; Rosbotham, J.L.; Jones, A.B.; Camp, R.D.R.; Frodsham, A.; Browne, J.; Barber, R.; Terwilliger, J.; Mark Lathrop, G.; et al. Identification of a Major Susceptibility Locus on Chromosome 6p and Evidence for Further Disease Loci Revealed by a Two Stage Genome-Wide Search in Psoriasis. Hum. Mol. Genet. 1997, 6, 813–820. [Google Scholar] [CrossRef]
- Burden, A.D.; Hodgins, M.; Tillman, D.; Javed, S.; Connor, M.; Bailey, M. Genetics of Psoriasis: Paternal Inheritance and a Locus on Chromosome 6p. J. Investig. Dermatol. 1998, 110, 958–960. [Google Scholar] [CrossRef]
- Owczarek, W. The role of HLA-Cw6 in psoriasis and psoriatic arthritis. Reumatologia. 2022, 60, 303–305. [Google Scholar] [CrossRef]
- Fan, X.; Yang, S.; Huang, W.; Wang, Z.-M.; Sun, L.-D.; Liang, Y.-H.; Gao, M.; Ren, Y.-Q.; Zhang, K.-Y.; Du, W.-H.; et al. Fine Mapping of the Psoriasis Susceptibility Locus PSORS1 Supports HLA-C as the Susceptibility Gene in the Han Chinese Population. PLoS Genet. 2008, 4, e1000038. [Google Scholar] [CrossRef]
- Gudjónsson, J.E.; Kárason, A.; Antonsdóttir, A.A.; Rúnarsdóttir, E.H.; Gulcher, J.R.; Stefánsson, K.; Valdimarsson, H. HLA-Cw6-Positive and HLA-Cw6-Negative Patients with Psoriasis Vulgaris Have Distinct Clinical Features. J. Investig. Dermatol. 2002, 118, 362–365. [Google Scholar] [CrossRef]
- Dand, N.; Duckworth, M.; Baudry, D.; Russell, A.; Curtis, C.J.; Lee, S.H.; Evans, I.; Mason, K.J.; Alsharqi, A.; Becher, G.; et al. HLA-C*06:02 Genotype Is a Predictive Biomarker of Biologic Treatment Response in Psoriasis. J. Allergy Clin. Immunol. 2019, 143, 2120–2130. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Jin, X.; Li, Y.; Jiang, H.; Tang, X.; Yang, X.; Cheng, H.; Qiu, Y.; Chen, G.; Mei, J.; et al. A Large-Scale Screen for Coding Variants Predisposing to Psoriasis. Nat. Genet. 2014, 46, 45–50. [Google Scholar] [CrossRef]
- Capon, F.; Spencer, C.C.A.; Knight, J.; Weale, M.E.; Allen, M.H.; Barton, A.; Band, G.; Bellenguez, C.; Bergboer, J.G.M.; Blackwell, J.M.; et al. A Genome-Wide Association Study Identifies New Psoriasis Susceptibility Loci and an Interaction between HLA-C and ERAP1. Nat. Genet. 2010, 42, 985–990. [Google Scholar]
- Fu, Y.; Li, X.; Chen, Y.; Liu, R.; Wang, R.; Bai, N. Association of ERAP1 Gene Polymorphisms with the Susceptibility to Psoriasis Vulgaris: A Case-Control Study: A Case–Control Study. Medicine 2018, 97, e12828. [Google Scholar] [CrossRef]
- Țiburcă, L.; Bembea, M.; Zaha, D.C.; Jurca, A.D.; Vesa, C.M.; Rațiu, I.A.; Jurca, C.M. The Treatment with Interleukin 17 Inhibitors and Immune-Mediated Inflammatory Diseases. Curr. Issues Mol. Biol. 2022, 44, 1851–1866. [Google Scholar] [CrossRef]
- Weigle, N.; McBane, S. Psoriasis. Am. Fam. Physician 2013, 87, 626–633. [Google Scholar]
- Tedeschi, V.; Paldino, G.; Paladini, F.; Mattorre, B.; Tuosto, L.; Sorrentino, R.; Fiorillo, M.T. The Impact of the “mis-Peptidome” on HLA Class I-Mediated Diseases: Contribution of ERAP1 and ERAP2 and Effects on the Immune Response. Int. J. Mol. Sci. 2020, 21, 9608. [Google Scholar] [CrossRef] [PubMed]
- Evnouchidou, I.; Kamal, R.P.; Seregin, S.S.; Goto, Y.; Tsujimoto, M.; Hattori, A.; Voulgari, P.V.; Drosos, A.A.; Amalfitano, A.; York, I.A.; et al. Cutting Edge: Coding Single Nucleotide Polymorphisms of Endoplasmic Reticulum Aminopeptidase 1 Can Affect Antigenic Peptide Generation in Vitro by Influencing Basic Enzymatic Properties of the Enzyme. J. Immunol. 2011, 186, 1909–1913. [Google Scholar] [CrossRef] [PubMed]
- York, I.A.; Brehm, M.A.; Zendzian, S.; Towne, C.F.; Rock, K.L. Endoplasmic Reticulum Aminopeptidase 1 (ERAP1) Trims MHC Class I-Presented Peptides in Vivo and Plays an Important Role in Immunodominance. Proc. Natl. Acad. Sci. USA 2006, 103, 9202–9207. [Google Scholar] [CrossRef]
- Fruci, D.; Giacomini, P.; Nicotra, M.R.; Forloni, M.; Fraioli, R.; Saveanu, L.; van Endert, P.; Natali, P.G. Altered Expression of Endoplasmic Reticulum Aminopeptidases ERAP1 and ERAP2 in Transformed Non-Lymphoid Human Tissues. J. Cell. Physiol. 2008, 216, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, Y.; Yan, Z.; Dai, S.; Liu, S.; Wang, X.; Wang, J.; Zhang, X.; Shi, L.; Yao, Y. Polymorphisms in Endoplasmic Reticulum Aminopeptidase Genes Are Associated with Cervical Cancer Risk in a Chinese Han Population. BMC Cancer 2020, 20, 341. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.M.; Jordanova, E.S.; Kenter, G.G.; Ferrone, S.; Fleuren, G.-J. Association of Antigen Processing Machinery and HLA Class I Defects with Clinicopathological Outcome in Cervical Carcinoma. Cancer Immunol. Immunother. 2008, 57, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Hasim, A.; Abudula, M.; Aimiduo, R.; Ma, J.-Q.; Jiao, Z.; Akula, G.; Wang, T.; Abudula, A. Post-Transcriptional and Epigenetic Regulation of Antigen Processing Machinery (APM) Components and HLA-I in Cervical Cancers from Uighur Women. PLoS ONE 2012, 7, e44952. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.esmo.org/content/download/303704/6040931/1/RO-Cancerul-de-Plaman-Fara-Celule-Mici-Ghidul-pentru-Pacienti.pdf (accessed on 20 November 2021).
- Wiśniewski, A.; Sobczyński, M.; Pawełczyk, K.; Porębska, I.; Jasek, M.; Wagner, M.; Niepiekło-Miniewska, W.; Kowal, A.; Dubis, J.; Jędruchniewicz, N.; et al. Polymorphisms of Antigen-Presenting Machinery Genes in Non-Small Cell Lung Cancer: Different Impact on Disease Risk and Clinical Parameters in Smokers and Never-Smokers. Front. Immunol. 2021, 12, 664474. [Google Scholar] [CrossRef]
- Yao, Y.; Wiśniewski, A.; Ma, Q.; Kowal, A.; Porębska, I.; Pawełczyk, K.; Yu, J.; Dubis, J.; Żuk, N.; Li, Y.; et al. Single Nucleotide Polymorphisms of the ERAP1 Gene and Risk of NSCLC: A Comparison of Genetically Distant Populations, Chinese and Caucasian. Arch. Immunol. Ther. Exp. 2016, 64 (Suppl. 1), 117–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Z.; Cao, R.; Zhang, Q.; Chen, T.; You, C.; Tang, W.; Chen, S. Association of BTLA Polymorphisms with Susceptibility to Non-Small-Cell Lung Cancer in the Chinese Population. Biomed Res. Int. 2021, 2021, 9121824. [Google Scholar] [CrossRef]
- Koumantou, D.; Barnea, E.; Martin-Esteban, A.; Maben, Z.; Papakyriakou, A.; Mpakali, A.; Kokkala, P.; Pratsinis, H.; Georgiadis, D.; Stern, L.J.; et al. Editing the Immunopeptidome of Melanoma Cells Using a Potent Inhibitor of Endoplasmic Reticulum Aminopeptidase 1 (ERAP1). Cancer Immunol. Immunother 2019, 68, 1245–1261. [Google Scholar] [CrossRef]
- Stratikos, E. Modulating Antigen Processing for Cancer Immunotherapy. Oncoimmunology 2014, 3, e27568. [Google Scholar] [CrossRef]
- Cifaldi, L.; Romania, P.; Lorenzi, S.; Locatelli, F.; Fruci, D. Role of Endoplasmic Reticulum Aminopeptidases in Health and Disease: From Infection to Cancer. Int. J. Mol. Sci. 2012, 13, 8338–8352. [Google Scholar] [CrossRef]
- Bufalieri, F.; Fruci, D.; Di Marcotullio, L. ERAP1 as an Emerging Therapeutic Target for Medulloblastoma. Trends Cancer 2021, 8, 4–8. [Google Scholar] [CrossRef]
- Niepiekło-Miniewska, W.; Matusiak, Ł.; Narbutt, J.; Lesiak, A.; Kuna, P.; Wiśniewski, A.; Kuśnierczyk, P. Contribution of Antigen-Processing Machinery Genetic Polymorphisms to Atopic Dermatitis. Life 2021, 11, 333. [Google Scholar] [CrossRef]
- Turner, J.D.; Schwartz, R.A. Atopic Dermatitis. A Clinical Challenge. Acta Dermatovenerol. Alp. Panonica Adriat. 2006, 15, 59–68. [Google Scholar]
- Thomsen, S.F. Epidemiology and Natural History of Atopic Diseases. Eur. Clin. Respir. J. 2015, 2, 24642. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V.; Hayashi, Y.; Jung, S.-H. An Overview of Severe Acute Respiratory Syndrome-Coronavirus (SARS-CoV) 3CL Protease Inhibitors: Peptidomimetics and Small Molecule Chemotherapy. J. Med. Chem. 2016, 59, 6595–6628. [Google Scholar] [CrossRef]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.-C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-Converting Enzyme 2: SARS-CoV-2 Receptor and Regulator of the Renin-Angiotensin System: Celebrating the 20th Anniversary of the Discovery of ACE2: Celebrating the 20th Anniversary of the Discovery of ACE2. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef] [PubMed]
- Vicenzi, M.; Di Cosola, R.; Ruscica, M.; Ratti, A.; Rota, I.; Rota, F.; Bollati, V.; Aliberti, S.; Blasi, F. The Liaison between Respiratory Failure and High Blood Pressure: Evidence from COVID-19 Patients. Eur. Respir. J. 2020, 56, 2001157. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, S.; Tempora, P.; Lucarini, V.; Melaiu, O.; Gaspari, S.; Algeri, M.; Fruci, D. ERAP1 and ERAP2 Enzymes: A Protective Shield for RAS against COVID-19? Int. J. Mol. Sci. 2021, 22, 1705. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, G.; Samiotaki, M.; Temponeras, I.; Panayotou, G.; Stratikos, E. Allotypic Variation in Antigen Processing Controls Antigenic Peptide Generation from SARS-CoV-2 S1 Spike Glycoprotein. J. Biol. Chem. 2021, 297, 101329. [Google Scholar] [CrossRef]
- Yamamoto, N.; Nakayama, J.; Yamakawa-Kobayashi, K.; Hamaguchi, H.; Miyazaki, R.; Arinami, T. Identification of 33 Polymorphisms in the Adipocyte-Derived Leucine Aminopeptidase (ALAP) Gene and Possible Association with Hypertension. Hum. Mutat. 2002, 19, 251–257. [Google Scholar] [CrossRef]
- Goto, Y.; Hattori, A.; Ishii, Y.; Mizutani, S.; Tsujimoto, M. Enzymatic Properties of Human Aminopeptidase A. Regulation of Its Enzymatic Activity by Calcium and Angiotensin IV. J. Biol. Chem. 2006, 281, 23503–23513. [Google Scholar] [CrossRef] [PubMed]
- Taranta, A.; Gianviti, A.; Palma, A.; De Luca, V.; Mannucci, L.; Procaccino, M.A.; Ghiggeri, G.M.; Caridi, G.; Fruci, D.; Ferracuti, S.; et al. Genetic Risk Factors in Typical Haemolytic Uraemic Syndrome. Nephrol. Dial. Transplant 2009, 24, 1851–1857. [Google Scholar] [CrossRef] [PubMed]
- Zee, R.Y.L.; Rivera, A.; Inostroza, Y.; Ridker, P.M.; Chasman, D.I.; Romero, J.R. Gene Variation of Endoplasmic Reticulum Aminopeptidases 1 and 2, and Risk of Blood Pressure Progression and Incident Hypertension among 17,255 Initially Healthy Women. Int. J. Genom. 2018, 2018, 2308585. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNPs Polymorphism | Amino Acid Residue Position | Major Allele (Sense Chain) | Minor Allele (Sense Chain) | Mutation (Missense) Allele Major → Allele Minor | Alleles Predisposing to AS |
|---|---|---|---|---|---|
| rs30187 | 528 | G | A | AGG → AAG R(Arg) → K(Lys) | Minor allele (coding for Lys, K) |
| rs27044 | 730 | G | C | GAA → CAA E(Glu) → Q(Gln) | Minor allele (coding for Gln, Q) |
| rs17482078 | 725 | G | A | CGA → CAA R(Arg) → Q(Gln) | Major allele (coding for Arg, R) |
| rs10050860 | 575 | G | A | GAC → AAC D(Asp) → N(Asn) | Major allele (coding for Asp, D) |
| rs2287987 | 349 | A | G | ATG → GTG M(Met) → V(Val) | Major allele (coding for Met, M) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Țiburcă, L.; Zaha, D.C.; Jurca, M.C.; Severin, E.; Jurca, A.; Jurca, A.D. The Role of Aminopeptidase ERAP1 in Human Pathology—A Review. Curr. Issues Mol. Biol. 2024, 46, 1651-1667. https://doi.org/10.3390/cimb46030107
Țiburcă L, Zaha DC, Jurca MC, Severin E, Jurca A, Jurca AD. The Role of Aminopeptidase ERAP1 in Human Pathology—A Review. Current Issues in Molecular Biology. 2024; 46(3):1651-1667. https://doi.org/10.3390/cimb46030107
Chicago/Turabian StyleȚiburcă, Laura, Dana Carmen Zaha, Maria Claudia Jurca, Emilia Severin, Aurora Jurca, and Alexandru Daniel Jurca. 2024. "The Role of Aminopeptidase ERAP1 in Human Pathology—A Review" Current Issues in Molecular Biology 46, no. 3: 1651-1667. https://doi.org/10.3390/cimb46030107
APA StyleȚiburcă, L., Zaha, D. C., Jurca, M. C., Severin, E., Jurca, A., & Jurca, A. D. (2024). The Role of Aminopeptidase ERAP1 in Human Pathology—A Review. Current Issues in Molecular Biology, 46(3), 1651-1667. https://doi.org/10.3390/cimb46030107