On the Neurobiology of Meditation: Comparison of Three Organizing Strategies to Investigate Brain Patterns during Meditation Practice


Abstract
:1. Introduction
2. First Strategy: Identify Underlying Brain Patterns of Meditation Practices
2.1. Changes in Gray Matter Volumes
2.2. Changes in White Matter Volumes
2.3. Summary of this Section
3. Second Strategy: Researching Individual Meditation Practices
- Loving-kindness/compassion meditation in expert meditators activated limbic areas and a network associated with Theory of Mind—right temporal lobes, temporo-parietal junction, medial prefrontal and posterior cingulate cortices [21,22,23].A meta-analysis of 16 fMRI studies of compassion meditation revealed patterns of activation in four brain areas: periaqueductal grey, anterior insula, anterior cingulate, and inferior frontal gyrus. This meta-analysis replicated earlier findings but did not find activation in areas typically connected with compassion including the dorsolateral prefrontal cortex, orbital prefrontal cortex and the amygdala [24].
- Mindfulness meditation was marked by deactivation of medial prefrontal cortices but activation of lateral prefrontal cortices, the secondary somatosensory cortex, the inferior parietal lobule and the insula [25,26,27,28].A meta-analysis of 21 fMRI studies of mindful practices used activation likelihood estimate methods to compare mindfulness with control conditions. Loci of activation were found in the frontal regions including the medial prefrontal gyrus, anterior cingulate, insula, and globus pallidus [29].
- Network analysis of imaging data from 12 experienced Zen meditators and 12 controls during an attention to breathing protocol reported extensive connections of frontoparietal circuits with early visual and executive control areas [36].
- Mantra recitation with meaning activated the hippocampi/parahippocampal area, middle cingulate cortex and the precentral cortex bilaterally [41].
Summary of this Section
4. Third Strategy: Organizing Meditations into Larger Classes
4.1. Grouping Meditations by Their Source: Indian or Buddhist Traditions
4.2. Grouping Meditations by their Procedures: Four Categories of Meditation
4.3. Grouping Meditations by EEG Activation Patterns: Three Categories of Meditation
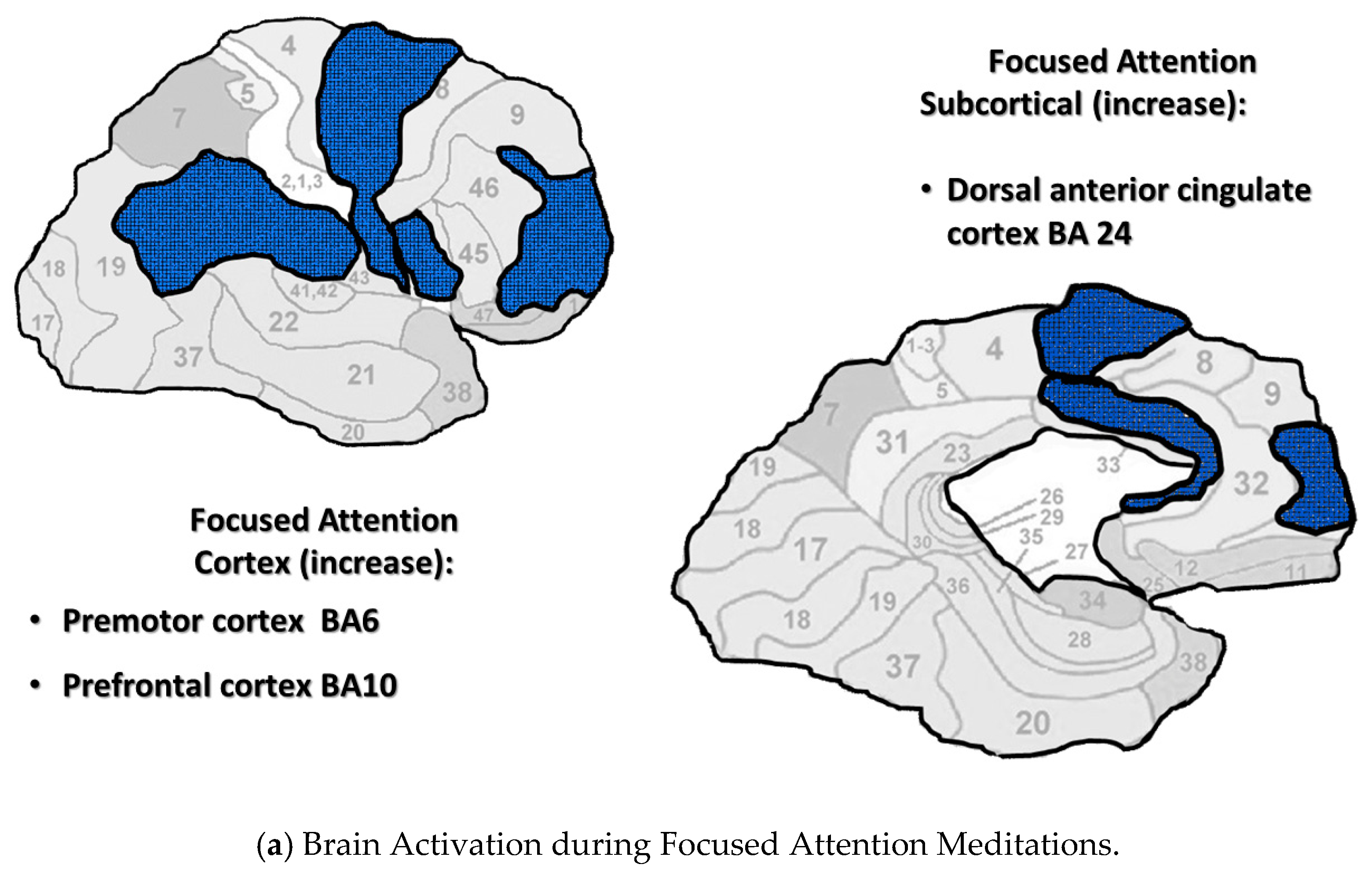
4.3.1. Focused Attention
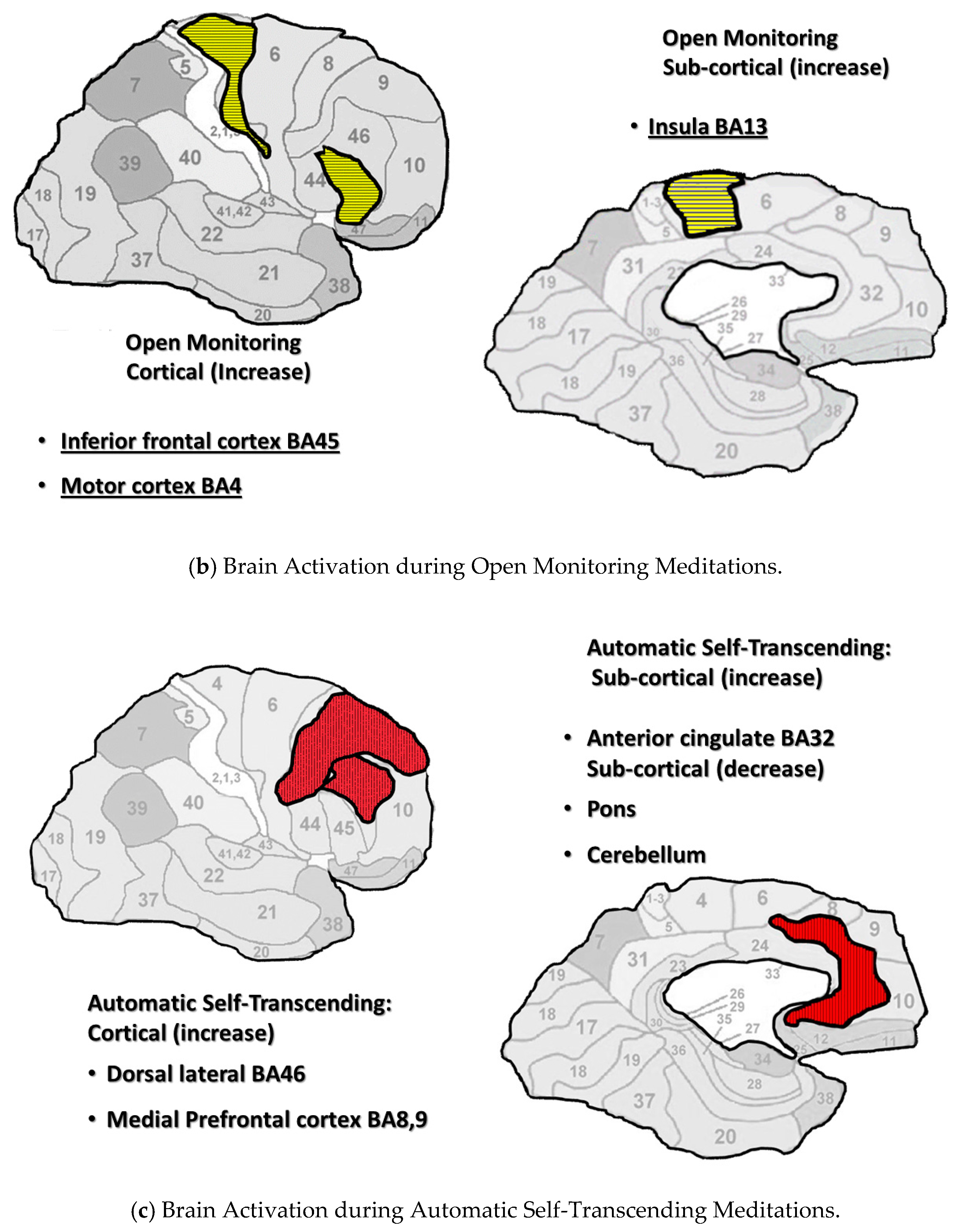
4.3.2. Open monitoring
4.3.3. Automatic Self-Transcending
4.3.4. Application of this Model
Automaticity: Why Zen Buddhism and Qigong Might Be in This Category
Default Mode Network Activation—A Marker of Automaticity
Default Mode Network and Mind Wandering
4.4. Summary of this Section
5. Recommended Grouping Strategy
Funding
Acknowledgments
Conflicts of Interest
References
- Stulberg, B.; Magness, S. Peak Performance: Elevate Your Game, Avoid Burnout, and Thrive with the New Science of Success; Rodale Books: New York, NY, USA, 2017. [Google Scholar]
- Spearman, C. General intelligence: Objectively determined and measured. Am. J. Psychol. 1904, 15, 201–292. [Google Scholar] [CrossRef]
- Undheim, J.O. On intelligence II: A neo-Spearman model to replace Cattell’s theory of fluid and crystallized intelligence. Scand. J. Psychol. 1981, 22, 181–187. [Google Scholar] [CrossRef]
- Helfrich, R.F.; Knight, R.T. Cognitive neurophysiology: Event-related potentials. Handb. Clin. Neurol. 2019, 160, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Luders, E.; Toga, A.W.; Lepore, N.; Gaser, C. The underlying anatomical correlates of long-term meditation: Larger hippocampal and frontal volumes of gray matter. Neuroimage 2009, 45, 672–678. [Google Scholar] [CrossRef] [Green Version]
- Luders, E.; Clark, K.; Narr, K.L.; Toga, A.W. Enhanced brain connectivity in long-term meditation practitioners. Neuroimage 2011, 57, 1308–1316. [Google Scholar] [CrossRef] [Green Version]
- Luders, E.; Cherbuin, N.; Kurth, F. Forever Young(er): Potential age-defying effects of long-term meditation on gray matter atrophy. Front. Psychol. 2014, 5, 1551. [Google Scholar] [CrossRef] [Green Version]
- Fox, K.C.; Nijeboer, S.; Dixon, M.L.; Floman, J.L.; Ellamil, M.; Rumak, S.P.; Sedlmeier, P.; Christoff, K. Is meditation associated with altered brain structure? A systematic review and meta-analysis of morphometric neuroimaging in meditation practitioners. Neurosci. Biobehav. Rev. 2014, 43, 48–73. [Google Scholar] [CrossRef]
- Lomas, T.; Ivtzan, I.; Fu, C.H. A systematic review of the neurophysiology of mindfulness on EEG oscillations. Neurosci. Biobehav. Rev. 2015, 57, 401–410. [Google Scholar] [CrossRef]
- Boccia, M.; Piccardi, L.; Guariglia, P. The Meditative Mind: A Comprehensive Meta-Analysis of MRI Studies. Biomed. Res. Int. 2015, 2015, 419808. [Google Scholar] [CrossRef] [Green Version]
- Luders, E.; Cherbuin, N.; Gaser, C. Estimating brain age using high-resolution pattern recognition: Younger brains in long-term meditation practitioners. Neuroimage 2016, 134, 508–513. [Google Scholar] [CrossRef]
- Luders, E.; Kurth, F. The neuroanatomy of long-term meditators. Curr. Opin. Psychol. 2019, 28, 172–178. [Google Scholar] [CrossRef]
- van Aalst, J.; Ceccarini, J.; Demyttenaere, K.; Sunaert, S.; Van Laere, K. What Has Neuroimaging Taught Us on the Neurobiology of Yoga? A Review. Front. Integr. Neurosci. 2020, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Hubener, M.; Bonhoeffer, T. Neuronal plasticity: Beyond the critical period. Cell 2014, 159, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackey, A.P.; Whitaker, K.J.; Bunge, S.A. Experience-dependent plasticity in white matter microstructure: Reasoning training alters structural connectivity. Front. Neuroanat. 2012, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Scholz, J.; Klein, M.C.; Behrens, T.E.; Johansen-Berg, H. Training induces changes in white-matter architecture. Nat. Neurosci. 2009, 12, 1370–1371. [Google Scholar] [CrossRef] [Green Version]
- Fox, K.C.; Dixon, M.L.; Nijeboer, S.; Girn, M.; Floman, J.L.; Lifshitz, M.; Ellamil, M.; Sedlmeier, P.; Christoff, K. Functional neuroanatomy of meditation: A review and meta-analysis of 78 functional neuroimaging investigations. Neurosci. Biobehav. Rev. 2016, 65, 208–228. [Google Scholar] [CrossRef] [Green Version]
- Ponlop, R.D. Wild Awakening: The Heart of Mahamudra and Dzogchen; Shambhala Publications: Boston, MA, USA, 2013. [Google Scholar]
- Brefczynski-Lewis, J.A.; Lutz, A.; Schaefer, H.S.; Levinson, D.B.; Davidson, R.J. Neural correlates of attentional expertise in long-term meditation practitioners. Proc. Natl. Acad. Sci. USA 2007, 104, 11483–11488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blase, K.L.; van Waning, A. Heart Rate Variability, Cortisol and Attention Focus During Shamatha Quiescence Meditation. Appl. Psychophysiol. Biofeedback 2019, 44, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Lutz, A.; Greischar, L.L.; Perlman, D.M.; Davidson, R.J. BOLD signal in insula is differentially related to cardiac function during compassion meditation in experts vs. novices. Neuroimage 2009, 47, 1038–1046. [Google Scholar] [CrossRef] [Green Version]
- Le Nguyen, K.D.; Lin, J.; Algoe, S.B.; Brantley, M.M.; Kim, S.L.; Brantley, J.; Salzberg, S.; Fredrickson, B.L. Loving-kindness meditation slows biological aging in novices: Evidence from a 12-week randomized controlled trial. Psychoneuroendocrinology 2019, 108, 20–27. [Google Scholar] [CrossRef]
- Leung, M.K.; Chan, C.C.; Yin, J.; Lee, C.F.; So, K.F.; Lee, T.M. Increased gray matter volume in the right angular and posterior parahippocampal gyri in loving-kindness meditators. Soc. Cogn. Affect. Neurosci. 2013, 8, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Cunnington, R.; Kirby, J.N. The neurophysiological basis of compassion: An fMRI meta-analysis of compassion and its related neural processes. Neurosci. Biobehav. Rev. 2020, 108, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Farb, N.A.S.; Segal, Z.V.; Mayberg, H.; Bean, J.; McKeon, D.; Fatima, Z. Attending to the present: Mindfulness meditation reveals distinct neural modes of self-reference. Soc. Cogn. Affect. Neurosci. 2007, 2, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, P.; Moore, A.W.; Mead, B.R.; Gruber, T. Mindful Aging: The Effects of Regular Brief Mindfulness Practice on Electrophysiological Markers of Cognitive and Affective Processing in Older Adults. Mindfulness 2017, 8, 78–94. [Google Scholar] [CrossRef] [Green Version]
- Singleton, O.; Holzel, B.K.; Vangel, M.; Brach, N.; Carmody, J.; Lazar, S.W. Change in Brainstem Gray Matter Concentration Following a Mindfulness-Based Intervention is Correlated with Improvement in Psychological Well-Being. Front. Hum. Neurosci. 2014, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Gotink, R.A.; Meijboom, R.; Vernooij, M.W.; Smits, M.; Hunink, M.G. 8-week Mindfulness Based Stress Reduction induces brain changes similar to traditional long-term meditation practice–A systematic review. Brain Cogn. 2016, 108, 32–41. [Google Scholar] [CrossRef]
- Falcone, G.; Jerram, M. Brain Activity in Mindfulness Depends on Experience: A Meta-Analysis of fMRI Studies. Mindfulness 2018, 9, 1319–1329. [Google Scholar] [CrossRef]
- Holzel, B.K.; Ott, U.; Hempel, H.; Hackl, A.; Wolf, K.; Stark, R. Differential engagement of anterior cingulate and adjacent medial frontal cortex in adept meditators and non-meditators. Neurosci. Lett. 2007, 421, 16–21. [Google Scholar] [CrossRef]
- Chiesa, A. Vipassana meditation: Systematic review of current evidence. J. Altern. Complement. Med. 2010, 16, 37–46. [Google Scholar] [CrossRef]
- Kakumanu, R.J.; Nair, A.K.; Venugopal, R.; Sasidharan, A.; Ghosh, P.K.; John, J.P.; Mehrotra, S.; Panth, R.; Kutty, B.M. Dissociating meditation proficiency and experience dependent EEG changes during traditional Vipassana meditation practice. Biol. Psychol. 2018, 135, 65–75. [Google Scholar] [CrossRef]
- Chiesa, A. Zen meditation: An integration of current evidence. J. Altern. Complement. Med. 2009, 15, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Lo, P.-C.; Tian, W.J.M.; Liu, F.-L. Macrostate and Microstate of EEG Spatio-Temporal Nonlinear Dynamics in Zen Meditation. J. Behav. Brain Sci. 2017, 7, 705–721. [Google Scholar] [CrossRef] [Green Version]
- Pagnoni, G.; Cekic, M.; Guo, Y. “Thinking about not-thinking”: Neural correlates of conceptual processing during Zen meditation. PLoS ONE 2008, 3, e3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemmer, P.B.; Guo, Y.; Wang, Y.; Pagnoni, G. Network-based characterization of brain functional connectivity in Zen practitioners. Front. Psychol. 2015, 6, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, H.C.; Kjaer, T.W.; Friberg, L.; Wildschiodtz, G.; Holm, S.; Nowak, M. A 15O–H2O PET study of meditation and the resting state of normal consciousness. Hum. Brain Mapp. 1999, 7, 98–105. [Google Scholar] [CrossRef]
- Parker, S. Training attention for conscious non-REM sleep: The yogic practice of yoga-nidra and its implications for neuroscience research. Prog. Brain Res. 2019, 244, 255–272. [Google Scholar] [CrossRef]
- Khalsa, D.S.; Amen, D.; Hanks, C.; Money, N.; Newberg, A. Cerebral blood flow changes during chanting meditation. Nucl. Med. Commun. 2009, 30, 956–961. [Google Scholar] [CrossRef]
- Arambula, P.; Peper, E.; Kawakami, M.; Gibney, K.H. The physiological correlates of Kundalini Yoga meditation: A study of a yoga master. Appl. Psychophysiol. Biofeedback 2001, 26, 147–153. [Google Scholar] [CrossRef]
- Engstrom, M.; Pihlsgard, J.; Lundberg, P.; Soderfeldt, B. Functional magnetic resonance imaging of hippocampal activation during silent mantra meditation. J. Altern. Complement. Med. 2010, 16, 1253–1258. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.J.; Rao, H.; Korczykowski, M.; Wintering, N.; Pluta, J.; Khalsa, D.S. Cerebral blood flow changes associated with different meditation practices and perceived depth of meditation. Psychiatry Res. 2011, 191, 60–67. [Google Scholar] [CrossRef]
- Moss, A.S.; Wintering, N.; Roggenkamp, H.; Khalsa, D.S.; Waldman, M.R.; Monti, D.; Newberg, A.B. Effects of an 8-week meditation program on mood and anxiety in patients with memory loss. J. Altern. Complement. Med. 2012, 18, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Aalst, J.; Ceccarini, J.; Schramm, G.; Van Weehaeghe, D.; Rezaei, A.; Demyttenaere, K.; Sunaert, S.; Van Laere, K. Long-term Ashtanga yoga practice decreases medial temporal and brainstem glucose metabolism in relation to years of experience. EJNMMI Res. 2020, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- Travis, F.; Haaga, D.A.; Hagelin, J.; Tanner, M.; Arenander, A.; Nidich, S.; Gaylord-King, C.; Grosswald, S.; Rainforth, M.; Schneider, R.H. A self-referential default brain state: Patterns of coherence, power, and eLORETA sources during eyes-closed rest and Transcendental Meditation practice. Cogn. Process. 2010, 11, 21–30. [Google Scholar] [CrossRef]
- Travis, F. Temporal and spatial characteristics of meditation EEG. Psychol. Trauma 2019, 12, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Travis, F.T.; Tecce, J.; Arenander, A.; Wallace, R.K. Patterns of EEG Coherence, Power, and Contingent Negative Variation Characterize the Integration of Transcendental and Waking States. Biol. Psychol. 2002, 61, 293–319. [Google Scholar] [CrossRef]
- Yamamoto, S.; Kitamura, Y.; Yamada, N.; Nakashima, Y.; Kuroda, S. Medial profrontal cortex and anterior cingulate cortex in the generation of alpha activity induced by transcendental meditation: A magnetoencephalographic study. Acta Med. Okayama 2006, 60, 51–58. [Google Scholar] [CrossRef]
- Mahone, M.C.; Travis, F.; Gevirtz, R.; Hubbard, D. fMRI during Transcendental Meditation practice. Brain Cogn. 2018, 123, 30–33. [Google Scholar] [CrossRef]
- Nash, J.D.; Newberg, A.; Awasthi, B. Toward a unifying taxonomy and definition for meditation. Front. Psychol. 2013, 4, 806. [Google Scholar] [CrossRef] [Green Version]
- Wynne, A. The Origin of Buddhist Meditation; Routledge: London, UK, 2009. [Google Scholar]
- Siegel, R.D.; Germer, C.K.; Olendzki, A. Mindfulness: What Is It? Where Did It Come from; Springer: New York, NY, USA, 2009. [Google Scholar]
- Nelson, J.B. Mindful Eating: The Art of Presence While You Eat. Diabetes Spectr. 2017, 3, 171–174. [Google Scholar] [CrossRef] [Green Version]
- Teut, M.; Roesner, E.J.; Ortiz, M.; Reese, F.; Binting, S.; Roll, S.; Fischer, H.F.; Michalsen, A.; Willich, S.N.; Brinkhaus, B. Mindful walking in psychologically distressed individuals: A randomized controlled trial. Evid. Based Complement. Altern. Med. 2013, 2013, 9–16. [Google Scholar] [CrossRef]
- Cho, H.; Ryu, S.; Noh, J.; Lee, J. The Effectiveness of Daily Mindful Breathing Practices on Test Anxiety of Students. PLoS ONE 2016, 11, e0164822. [Google Scholar] [CrossRef] [PubMed]
- Kabat-Zinn, J. Mindfulness-Based Interventions in Context: Past, Present, and Future. Clin. Psychol. Sci. Pract. 2003, 10, 144–156. [Google Scholar] [CrossRef]
- Tomasino, B.; Chiesa, A.; Fabbro, F. Disentangling the neural mechanisms involved in Hinduism- and Buddhism-related meditations. Brain Cogn. 2014, 90, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Chogyam, N.; Dechen, K. Roaring Silence: Discovering the Mind of Dozgchen; Shambhalam Publications: Boston, MA, USA, 2002. [Google Scholar]
- Maharishi, M.Y. Science of Being and Art of Living; Plume: New York, NY, USA, 2001. [Google Scholar]
- Tanahasi, K. Beyond THinking: A Guide to Zen Meidtation; Shambhala: Boston, MA, USA, 2004. [Google Scholar]
- Lutz, A.; Slagter, H.A.; Dunne, J.D.; Davidson, R.J. Attention regulation and monitoring in meditation. Trends Cogn. Sci. 2008, 12, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travis, F.; Shear, J. Focused attention, open monitoring and automatic self-transcending: Categories to organize meditations from Vedic, Buddhist and Chinese traditions. Conscious. Cogn. 2010, 19, 1110–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braboszcz, C.; Cahn, B.R.; Levy, J.; Fernandez, M.; Delorme, A. Increased Gamma Brainwave Amplitude Compared to Control in Three Different Meditation Traditions. PLoS ONE 2017, 12, e0170647. [Google Scholar] [CrossRef]
- Fries, P.; Womelsdorf, T.; Oostenveld, R.; Desimone, R. The effects of visual stimulation and selective visual attention on rhythmic neuronal synchronization in macaque area V4. J. Neurosci. 2008, 28, 4823–4835. [Google Scholar] [CrossRef] [Green Version]
- Fell, J.; Fernandez, G.; Klaver, P.; Elger, C.E.; Fries, P. Is synchronized neuronal gamma activity relevant for selective attention? Brain Res. Brain Res. Rev. 2003, 42, 265–272. [Google Scholar] [CrossRef]
- Jensen, O.; Mazaheri, A. Shaping functional architecture by oscillatory alpha activity: Gating by inhibition. Front. Hum. Neurosci. 2010, 4, 186. [Google Scholar] [CrossRef] [Green Version]
- van Lutterveld, R.; Houlihan, S.D.; Pal, P.; Sacchet, M.D.; McFarlane-Blake, C.; Patel, P.R.; Sullivan, J.S.; Ossadtchi, A.; Druker, S.; Bauer, C.; et al. Source-space EEG neurofeedback links subjective experience with brain activity during effortless awareness meditation. Neuroimage 2017, 151, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, L.T.; Ranganath, C. Frontal midline theta oscillations during working memory maintenance and episodic encoding and retrieval. Neuroimage 2014, 85 Pt 2, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Brandmeyer, T.; Delorme, A. Reduced mind wandering in experienced meditators and associated EEG correlates. Exp. Brain Res. 2018, 236, 2519–2528. [Google Scholar] [CrossRef]
- Palva, S.; Palva, J.M. Functional roles of alpha-band phase synchronization in local and large-scale cortical networks. Front. Psychol. 2011, 2, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litscher, G.; Wenzel, G.; Niederwieser, G.; Schwarz, G. Effects of QiGong on brain function. Neurol. Res. 2001, 23, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Cahn, B.R.; Delorme, A.; Polich, J. Occipital gamma activation during Vipassana meditation. Cogn. Process. 2010, 11, 39–56. [Google Scholar] [CrossRef] [Green Version]
- Schoenberg, P.L.A.; Ruf, A.; Churchill, J.; Brown, D.P.; Brewer, J.A. Mapping complex mind states: EEG neural substrates of meditative unified compassionate awareness. Conscious. Cogn. 2018, 57, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Pagnoni, G. The contemplative exercise through the lenses of predictive processing: A promising approach. Prog. Brain Res. 2019, 244, 299–322. [Google Scholar] [CrossRef] [PubMed]
- Aftanas, L.I.; Golocheikine, S.A. Human anterior and frontal midline theta and lower alpha reflect emotionally positive state and internalized attention: High-resolution EEG investigation of meditation. Neurosci. Lett. 2001, 310, 57–60. [Google Scholar] [CrossRef]
- Baijal, S.; Srinivasan, N. Theta activity and meditative states: Spectral changes during concentrative meditation. Cogn. Process. 2010, 11, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.; Jin, Y.; Lin, S.; Hermanowicz, N.S. A forty-five year follow-up EEG study of Qigong practice. Int. J. Neurosci. 2009, 119, 538–552. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lo, P.C. EEG dynamics of experienced Zen meditation practitioners probed by complexity index and spectral measure. J. Med. Eng. Technol. 2009, 33, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Brandmeyer, T.; Delorme, A.; Wahbeh, H. The neuroscience of meditation: Classification, phenomenology, correlates, and mechanisms. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2019; Volume 244, pp. 1–29. [Google Scholar]
- Schneider, W.; Pimm-Smith, M.; Worden, M. Neurobiology of attention and automaticity. Curr. Opin. Neurobiol. 1994, 4, 177–182. [Google Scholar] [CrossRef]
- Garrison, K.A.; Santoyo, J.F.; Davis, J.H.; Thornhill, T.A.T.; Kerr, C.E.; Brewer, J.A. Effortless awareness: Using real time neurofeedback to investigate correlates of posterior cingulate cortex activity in meditators’ self-report. Front. Hum. Neurosci. 2013, 7, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maharishi, M.Y. Maharishi Mahesh Yogi on the Bhagavad Gita; Penguin Books: New York, NY, USA, 1969. [Google Scholar]
- Franklin, M.S.; Mrazek, M.D.; Anderson, C.L.; Smallwood, J.; Kingstone, A.; Schooler, J.W. The silver lining of a mind in the clouds: Interesting musings are associated with positive mood while mind-wandering. Front. Psychol. 2013, 4, 583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travis, F.; Parim, N. Default mode network activation and Transcendental Meditation practice: Focused Attention or Automatic Self-transcending? Brain Cogn. 2017, 111, 86–94. [Google Scholar] [CrossRef]
- Fox, M.D.; Raichle, M.E. Spontaneous fluctuations in brain activity observed with functional magnetic resonance imaging. Nat. Rev. Neurosci. 2007, 8, 700–711. [Google Scholar] [CrossRef]
- Raichle, M.E. The brain’s default mode network. Annu. Rev. Neurosci. 2015, 38, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Raichle, M.E.; Macleod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Buckner, R.L.; Vincent, J.L. Unrest at rest: Default activity and spontaneous network correlations. Neuroimage 2007, 37, 1091–1096. [Google Scholar] [CrossRef]
- Garrison, K.A.; Zeffiro, T.A.; Scheinost, D.; Constable, R.T.; Brewer, J.A. Meditation leads to reduced default mode network activity beyond an active task. Cogn. Affect. Behav. Neurosci. 2015, 15, 712–720. [Google Scholar] [CrossRef] [Green Version]
- Simon, R.; Engstrom, M. The default mode network as a biomarker for monitoring the therapeutic effects of meditation. Front. Psychol. 2015, 6, 776. [Google Scholar] [CrossRef] [PubMed]
- Bærentsen, K.B.; Stødkilde-Jørgensen, H.; Sommerlund, B.; Hartmann, T.; Damsgaard-Madsen, J.; Fosnæs, M. An investigation of brain processes supporting meditation. Cogn. Process. 2010, 11, 57–84. [Google Scholar] [CrossRef] [PubMed]
- Ives-Deliperi, V.L.; Solms, M.; Meintjes, E.M. The neural substrates of mindfulness: An fMRI investigation. Soc. Neurosci. 2011, 6, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.A.; Grant, J.; Daneault, V.; Scavone, G.; Breton, E.; Roffe-Vidal, S. Impact of mindfulness on the neural responses to emotional pictures in experienced and beginner meditators. Neuroimage 2011, 57, 1524–1533. [Google Scholar] [CrossRef]
- Brewer, J.A.; Worhunsky, P.D.; Gray, J.R.; Tang, Y.Y.; Weber, J.; Kober, H. Meditation experience is associated with differences in default mode network activity and connectivity. Proc. Natl. Acad. Sci. USA 2011, 108, 20254–20259. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.X.; Chen, X.; Shen, Y.Q.; Li, L.; Chen, N.X.; Zhu, Z.C.; Castellanos, F.X.; Yan, C.G. Rumination and the default mode network: Meta-analysis of brain imaging studies and implications for depression. Neuroimage 2020, 206, 116287. [Google Scholar] [CrossRef]
- Antrobus, J.S.; Singer, J.L.; Goldstein, S.; Fortgang, M.S. Mind wandering and cognitive structure. Trans. N. Y. Acad. Sci. 1970, 32, 242–252. [Google Scholar] [CrossRef]
- Singer, J.L. Navigating the stream of consciousness: Research in daydreaming and related inner experiences. Am. Psychol. 1975, 30, 727–738. [Google Scholar] [CrossRef]
- Killingsworth, M.A.; Gilbert, D.T. A wandering mind is an unhappy mind. Science 2010, 330, 932. [Google Scholar] [CrossRef] [Green Version]
- Stawarczyk, D.; Majerus, S.; Maj, M.; Van der Linden, M.; D’Argembeau, A. Mind-wandering: Phenomenology and function as assessed with a novel experience sampling method. Acta Psychol. 2011, 136, 370–381. [Google Scholar] [CrossRef]
- Baird, B.; Smallwood, J.; Schooler, J.W. Back to the future: Autobiographical planning and the functionality of mind-wandering. Conscious. Cogn. 2011, 20, 1604–1611. [Google Scholar] [CrossRef] [PubMed]
- Baird, B.; Smallwood, J.; Mrazek, M.D.; Kam, J.W.; Franklin, M.S.; Schooler, J.W. Inspired by distraction: Mind wandering facilitates creative incubation. Psychol. Sci. 2012, 23, 1117–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vess, M.; Leal, S.A.; Hoeldtke, R.T.; Schlegel, R.J.; Hicks, J.A. True self-alienation positively predicts reports of mindwandering. Conscious. Cogn. 2016, 45, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Hobbiss, M.H.; Fairnie, J.; Jafari, K.; Lavie, N. Attention, mindwandering, and mood. Conscious. Cogn. 2019, 72, 1–18. [Google Scholar] [CrossRef]
- Ottaviani, C.; Shapiro, D.; Couyoumdjian, A. Flexibility as the key for somatic health: From mind wandering to perseverative cognition. Biol. Psychol. 2013, 94, 38–43. [Google Scholar] [CrossRef]
- Ottaviani, C.; Couyoumdjian, A. Pros and cons of a wandering mind: A prospective study. Front. Psychol. 2013, 4, 524. [Google Scholar] [CrossRef] [Green Version]
- Travis, F. Negative and Positive Mindwandering; IGI Global: Hershey, PA, USA, 2020. [Google Scholar]



{kind=link}
{kind=link}
| Reference | Design | Result |
|---|---|---|
| Luders [5] | 22 meditation and 22 age-matched controls. Meditations included Zazen, Shamatha, Vipassana, and Others: 63% used deep concentration, 36% control of breath, 32% visualization, 32% attention to external and internal stimuli, 14% withdrawal of sensory perceptions and 18% letting go of thoughts. | Total grey matter volumes were similar in both groups (meditator and control). There were local differences. Meditators had thicker:
|
| Luders [6] | 27 meditation and 27 age-matched controls. Average age: 52 years. | Fractional anisotropy (FA) (integrity of white matter) was higher in meditators compared to controls within major projection pathways, commissural pathways, and association pathways. |
| 55% practiced: Shamatha, Vipassana, or Zazen. | ||
| Luders [7] | 50 long-term meditators and 50 age-matched controls. Age range: 27–71 years. | Negative correlation of global and local gray matter with age for both meditators and controls. However, the group-by-age interaction was highly significant (p = 0.003) with lower grey matter by age for controls. |
| Meditations included Zazen, Shamatha, Vipassana. | ||
| Fox [8] | Meta-analysis of 21 neuroimaging studies with N = 300 meditation practitioners: Insight, Zen, Tibetan, Buddhist, Mindfulness Based Stress Reduction, Integrative Mind-Body Training, Soham, Loving-Kindness meditation, and “Various”. | Five regions were thicker in meditators:
|
| Lomas [9] | Meta-analysis of 54 studies of EEG from Mindfulness Based Cognitive Training (2), Mindfulness (13), Vipassana (6), mind/body training (2), Zen (13), various (5). |
|
| Boccio [10] | Meta-analysis with Mindfulness practices (5), Vipassana (12), Kundalini (11), Integrative Body Mind Training (3), MBSR (11), Zen (4). | Areas activated included the medial prefrontal cortex, motor cortex gyrus, anterior cingulate cortex, insula, claustrum, precuneus, parahippocampal gyrus, middle occipital gyrus, inferior parietal lobule, lentiform nucleus and thalamus. |
| Luders [11] | 50 long-term meditators and 50 age-matched controls. | Estimating brain ages.
|
| Meditations included Zazen, Shamatha, Vipassana, as in the first study. | ||
| Luders [12] | Meta-analysis of 9 studies: Dzogchen, Loving-kindness meditation, Sahaja yoga meditation, Shamatha, Vipassana, Zen. | Long term meditators (10 years or more) vs. controls had:
|
| Van Aalst [13] | Meta-analysis of 34 studies of Yoga practice: Ashtanga, Iyengar, Vinyasa, Kripalu, Kundalini, Nidra, Sahaja, Sivananda and Hatha yoga. | Yoga practitioners had increased gray matter volume in the insula and hippocampus and increased activation of prefrontal cortical regions. There was, however, high variability in the neuroimaging findings |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Travis, F. On the Neurobiology of Meditation: Comparison of Three Organizing Strategies to Investigate Brain Patterns during Meditation Practice. Medicina 2020, 56, 712. https://doi.org/10.3390/medicina56120712
Travis F. On the Neurobiology of Meditation: Comparison of Three Organizing Strategies to Investigate Brain Patterns during Meditation Practice. Medicina. 2020; 56(12):712. https://doi.org/10.3390/medicina56120712
Chicago/Turabian StyleTravis, Frederick. 2020. "On the Neurobiology of Meditation: Comparison of Three Organizing Strategies to Investigate Brain Patterns during Meditation Practice" Medicina 56, no. 12: 712. https://doi.org/10.3390/medicina56120712
APA StyleTravis, F. (2020). On the Neurobiology of Meditation: Comparison of Three Organizing Strategies to Investigate Brain Patterns during Meditation Practice. Medicina, 56(12), 712. https://doi.org/10.3390/medicina56120712