Tadalafil Treatment Ameliorates Hypoxia and Alters Placental Expression of Proteins Downstream of mTOR Signaling in Fetal Growth Restriction

 ,
,  , , ,
, , ,
Abstract
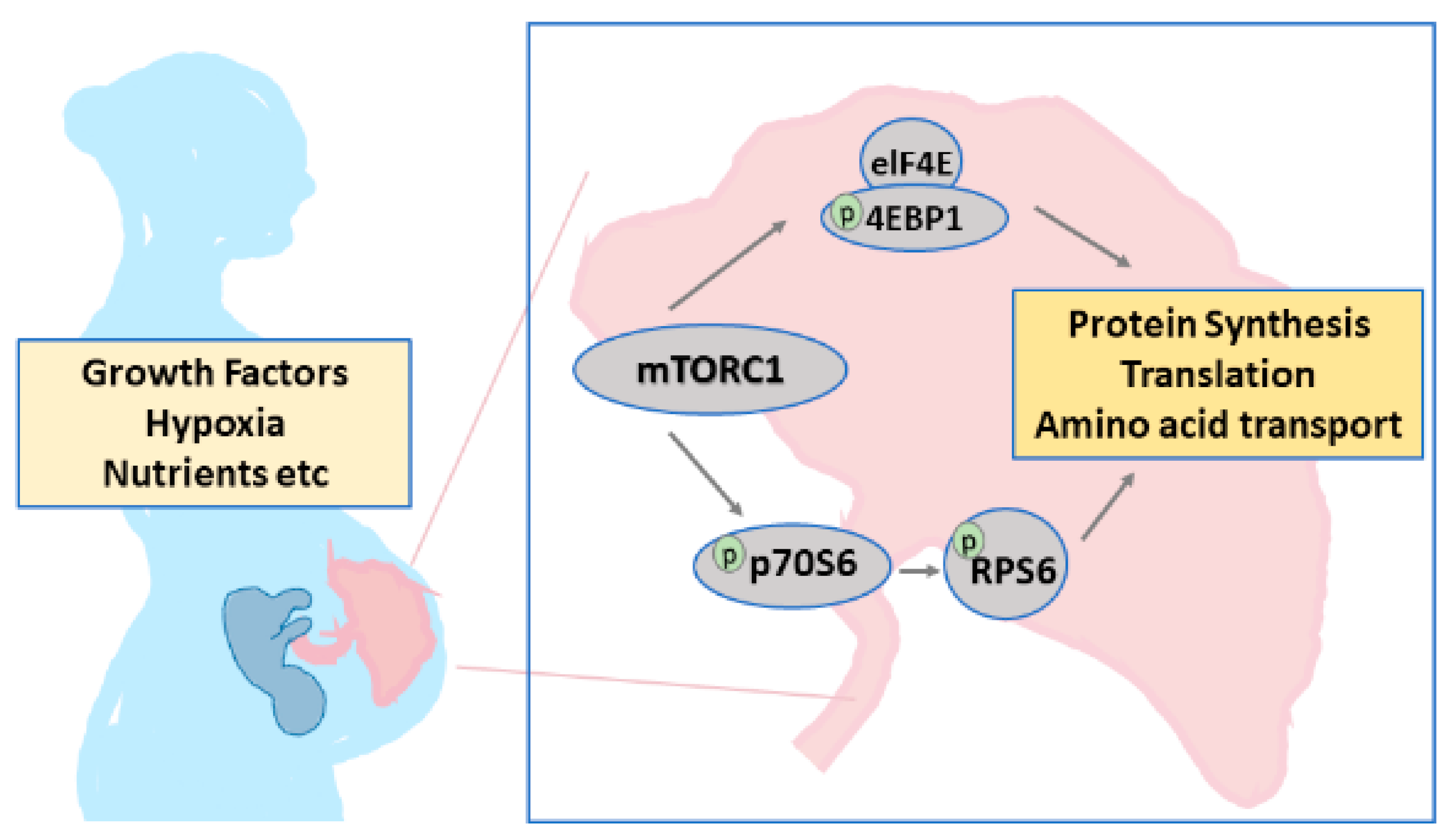
:1. Introduction
2. Materials and Methods
2.1. Collection of Placental Samples
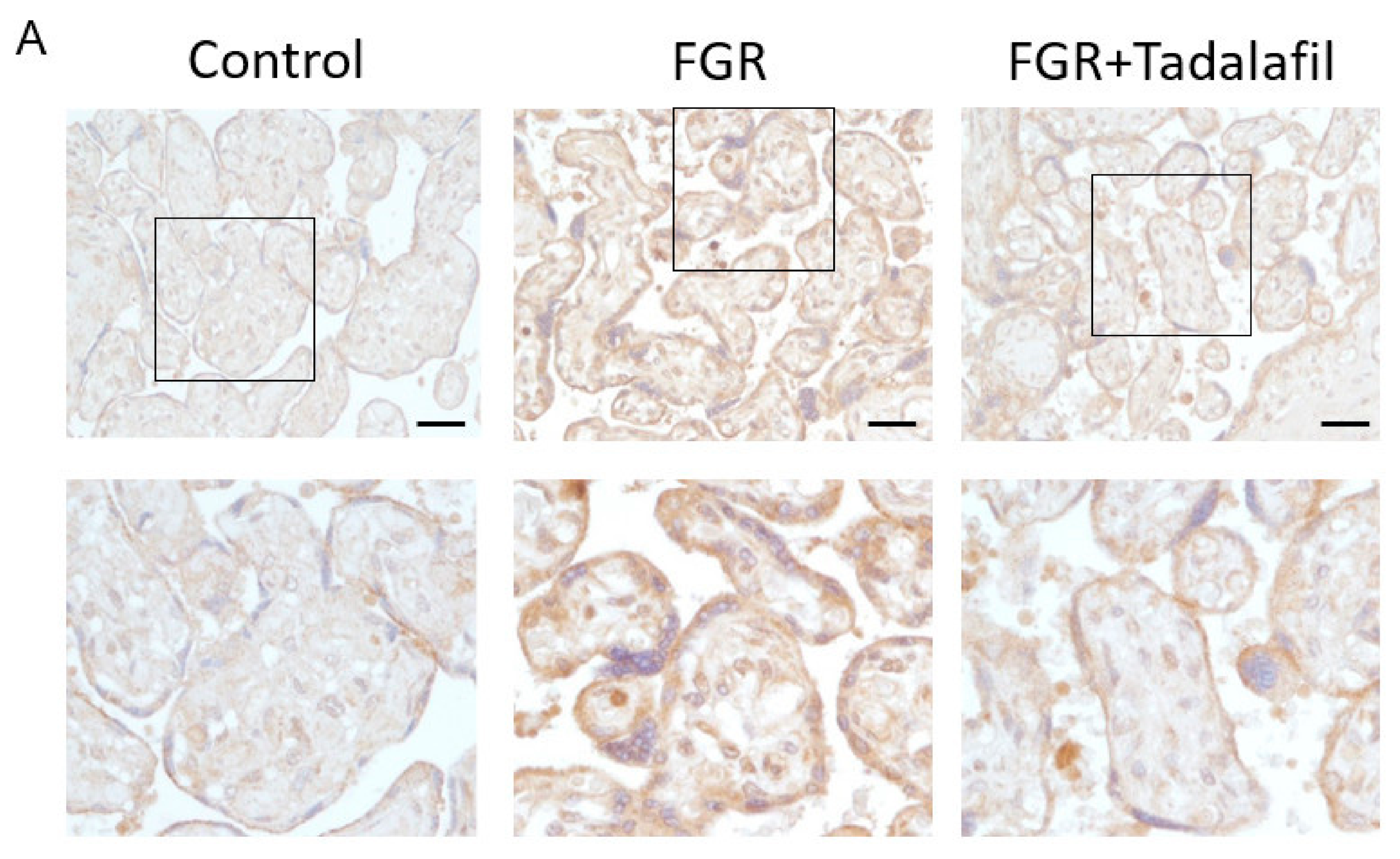
2.2. Histological Analysis and Immunohistochemistry (IHC) of Placenta
2.3. Protein Extraction
2.4. Western Blot Analysis
2.5. Enzyme-Linked Immunosorbent Assay
2.6. Statistical Analysis
3. Results
3.1. Clinical Characteristics
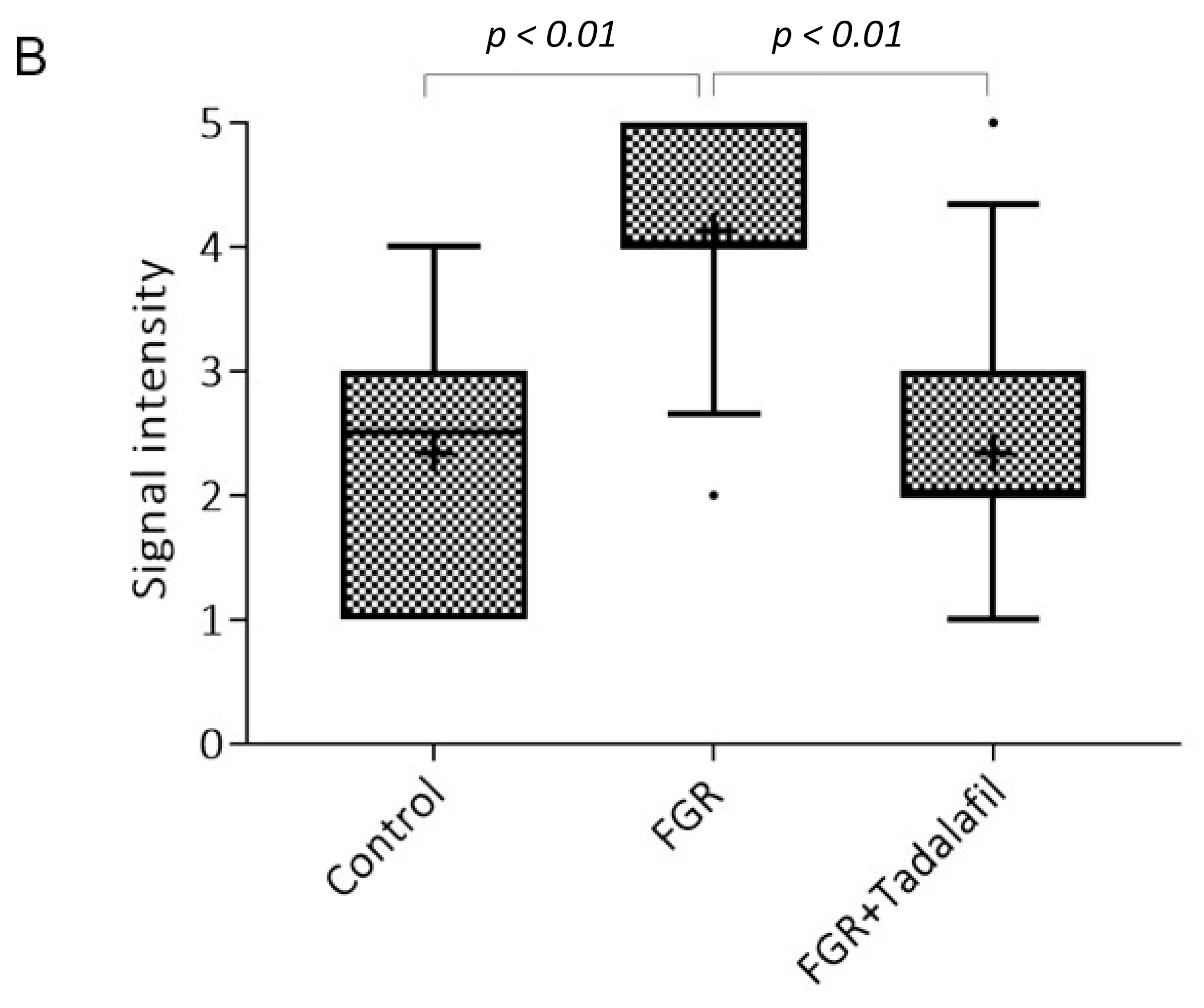
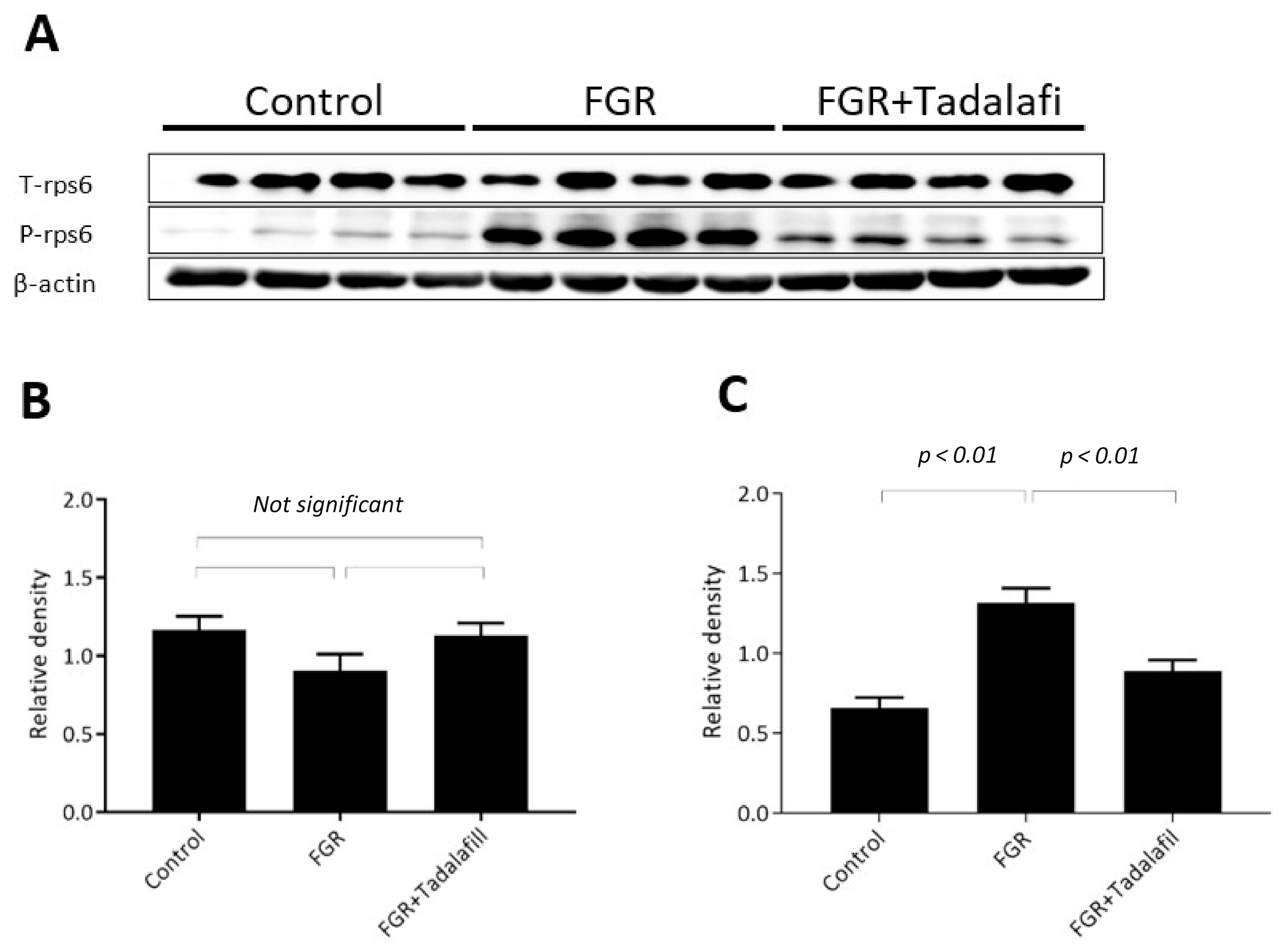
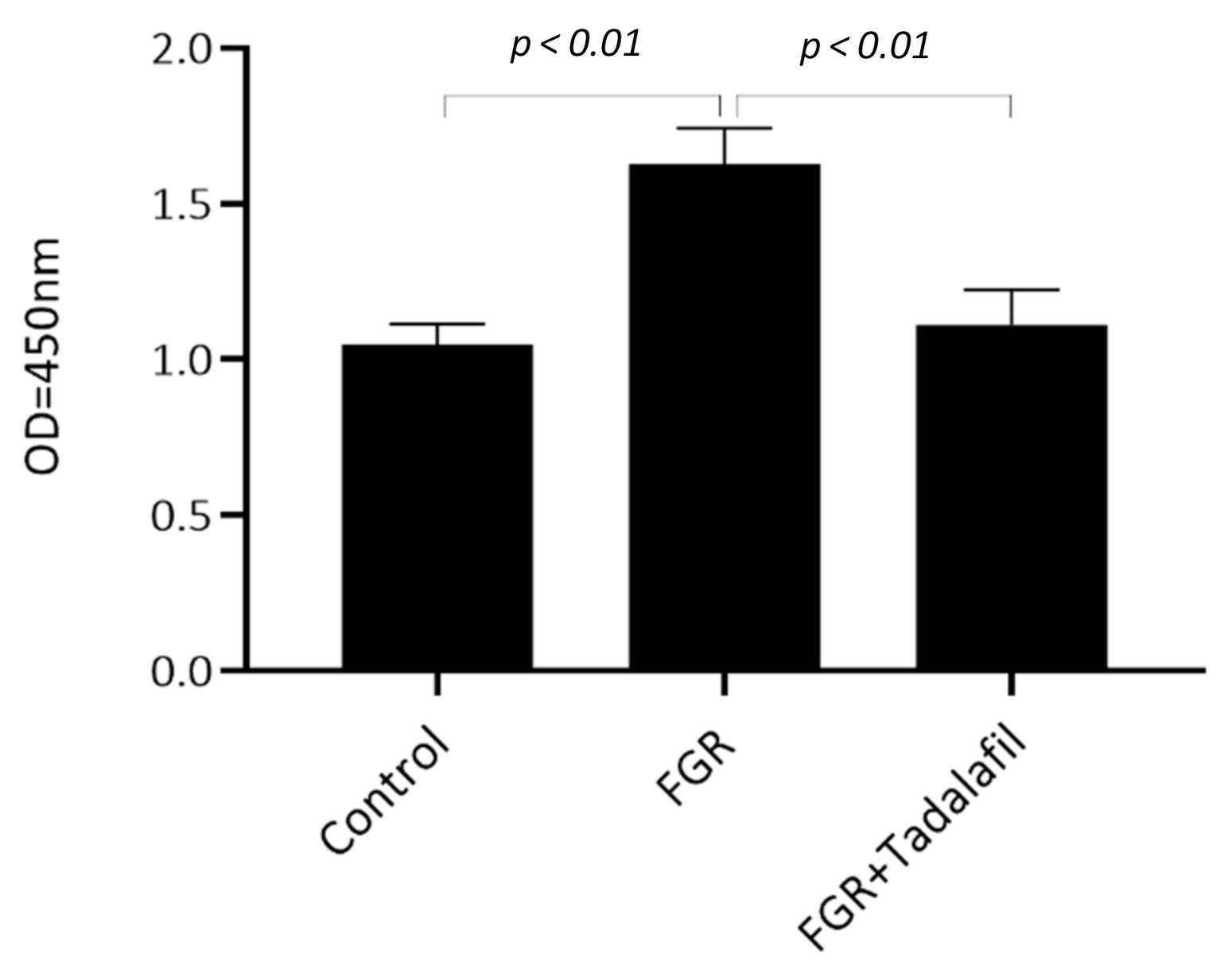
3.2. Effect of Tadalafil Treatment on mTOR Signaling in FGR Placenta
3.3. Effect of Tadalafil Treatment on rpS6 and eIF-4E in FGR Placentas
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- American College of Obstetricians and Gynecologists. ACOG Practice bulletin no. 134: Fetal growth restriction. Obstet. Gynecol. 2013, 121, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Beebe, L.A.; Cowan, L.D.; Altshuler, G. The epidemiology of placental features: Associations with gestational age and neonatal outcome. Obstet. Gynecol. 1996, 87, 771–778. [Google Scholar] [CrossRef]
- Baschat, A.A. Planning management and delivery of the growth-restricted fetus. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 49, 53–65. [Google Scholar] [CrossRef]
- Hochner, H.; Friedlander, Y.; Calderon-Margalit, R.; Meiner, V.; Sagy, Y.; Avgil-Tsadok, M.; Burger, A.; Savitsky, B.; Siscovick, D.S.; Manor, O. Associations of maternal prepregnancy body mass index and gestational weight gain with adult offspring cardiometabolic risk factors: The Jerusalem Perinatal Family Follow-up Study. Circulation 2012, 125, 1381–1389. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.L.; Huppi, P.S.; Mallard, C. The consequences of fetal growth restriction on brain structure and neurodevelopmental outcome. J. Physiol. 2016, 594, 807–823. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, K.; Umekawa, T.; Maki, S.; Kubo, M.; Nii, M.; Tanaka, K.; Tanaka, H.; Osato, K.; Kamimoto, Y.; Kondo, E.; et al. Tadalafil improves L-NG-nitroarginine methyl ester-induced preeclampsia with fetal growth restriction-like symptoms in pregnant mice. Am. J. Hypertens 2017, 31, 89–96. [Google Scholar] [CrossRef]
- Kubo, M.; Umekawa, T.; Maekawa, Y.; Tanaka, H.; Nii, M.; Murabayashi, N.; Osato, K.; Kamimoto, Y.; Ikeda, T. Retrospective study of tadalafil for fetal growth restriction: Impact on maternal and perinatal outcomes. J. Obstet. Gynaecol. Res. 2017, 43, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Kubo, M.; Tanaka, H.; Maki, S.; Nii, M.; Murabayashi, N.; Osato, K.; Kamimoto, Y.; Umekawa, T.; Ikeda, T. Safety and dose-finding trial of tadalafil administered for fetal growth restriction: A phase-1 clinical study. J. Obstet. Gynaecol. Res. 2017, 43, 1159–1168. [Google Scholar] [CrossRef]
- Tanaka, H.; Kubo, M.; Nii, M.; Maki, S.; Umekawa, T.; Ikeda, T. Treatment using tadalafil for severe pre-eclampsia with fetal growth restriction. J. Obstet. Gynaecol. Res. 2017, 43, 1205–1208. [Google Scholar] [CrossRef]
- Furuhashi, F.H.; Tanaka, H.; Kaneda, M.; Maki, S.; Nii, M.; Umekawa, T.; Osato, K.; Kamimoto, Y.; Ikeda, T. Safety trial of tadalafil administered for the treatment of preeclampsia. J. Matern. Fetal Neonatal. Med. 2020, 33, 167–170. [Google Scholar] [CrossRef]
- Tanaka, K.; Tanaka, H.; Maki, S.; Kubo, M.; Nii, M.; Magawa, S.; Hatano, F.; Tsuji, M.; Osato, K.; Kamimoto, Y.; et al. Cardiac function and tadalafil used for treating fetal growth restriction in pregnant women without cardiovascular disease. J. Matern. Fetal Neonatal. Med. 2019, 32, 2460–2462. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, R.; Umekawa, T.; Yoshikawa, K.; Owa, T.; Magawa, S.; Furuhashi, F.; Tsuji, T.; Maki, S.; Shimada, K.; Kaneda, M.K.; et al. Tadalafil treatment in mice for preeclampsia with fetal growth restriction has neuro-benefic effects in offspring through modulating prenatal hypoxic conditions. Sci. Rep. 2019, 9, 234. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.J. Comparison of phosphodiesterase type 5 (PDE5) inhibitors. Int. J. Clin. Pract. 2006, 60, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Peng, T.; Golub, T.R.; Sabatini, D.M. The Immunosuppressant Rapamycin Mimics a Starvation-Like Signal Distinct from Amino Acid and Glucose Deprivation. Mol. Cell Biol. 2002, 22, 5575–5584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tee, A.R.; Blenis, J. mTor, translational control and human disease. Sem. Cell Dev. Biol. 2005, 16, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.E.; Hall, M.N. The expanding TOR network. Curr. Opin. Cell Biol. 2005, 17, 158–166. [Google Scholar] [CrossRef]
- Hay, N.; Soneneberg, N. Upstream and downstream of mTOR. Genes Dev. 2004, 18, 1926–1945. [Google Scholar] [CrossRef] [Green Version]
- Jacinto, E.; Hall, M.N. TOR signalling in bugs, brain and brawn. Nature Rev. Mol. Cell Biol. 2003, 4, 117–126. [Google Scholar] [CrossRef]
- Maki, S.; Tanaka, H.; Tsuji, M.; Furuhashi, F.; Magawa, S.; Kaneda, M.K.; Nii, M.; Tanaka, K.; Kondo, E.; Tamaru, S.; et al. Safety Evaluation of Tadalafil Treatment for Fetuses with Early-Onset Growth Restriction (TADAFER): Results from the Phase II Trial. J. Clin. Med. 2019, 8, 856. [Google Scholar] [CrossRef] [Green Version]
- Betz, C.; Hall, M.N. Where is mTOR and what is it doing there? J. Cell Biol. 2013, 203, 563–574. [Google Scholar] [CrossRef] [Green Version]
- Blommaart, E.F.; Luiken, J.J.; Blommaart, P.J.; van Woerkom, G.M.; Meijer, A.J. Phosphorylation of Ribosomal Protein S6 Is Inhibitory for Autophagy in Isolated Rat Hepatocytes. J. Biol. Chem. 1995, 270, 2320–2326. [Google Scholar] [CrossRef] [Green Version]
- Lacey, H.; Haigh, T.; Westwood, M.; Aplin, J.D. Mesenchymally-derived insulin-like growth factor 1 provides a paracrine stimulus for trophoblast migration. BMC Dev. Biol. 2002, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, H.; Sakai, K.; Kabir-Salmani, M.; Wachi, Y.; Iwashita, M. Polymerization of insulin-like growth factor-binding protein-1 (IGFBP-1) potentiates IGF-I actions in placenta. J. Cell Physiol. 2011, 226, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Mayama, R.; Izawa, T.; Sakai, K.; Suciu, N.; Iwashita, M. Improvement of insulin sensitivity promotes extravillous trophoblast cell migration stimulated by insulin-like growth factor-I. Endocr. J. 2013, 60, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Yukiyo, A.; Askew, D.J.; Aramaki, S.; Myoga, M.; Tomonaga, C.; Hachisuga, T.; Suga, R.; Kawamoto, T.; Tsuji, M.; Shibata, E. Differential Levels of Amino Acid Transporters System L and ASCT2, and the mTOR Protein in Placenta of Preeclampsia and IUGR. BMC Pregnancy Childbirth 2014, 14, 181. [Google Scholar]
- Minakami, H.; Maeda, T.; Fujii, T.; Hamada, H.; Iitsuka, Y.; Itakura, A.; Itoh, H.; Iwashita, M.; Kanagawa, T.; Kanai, M.; et al. Guidelines for obstetrical practice in Japan: Japan Society of Obstetrics and Gynecology (JSOG) and Japan Association of Obstetricians and Gynecologists (JAOG) 2014 edition. J. Obstet. Gynaecol. Res. 2014, 40, 1469–1499. [Google Scholar] [CrossRef]
- Hui, L.; Challis, D. Diagnosis and management of fetal growth restriction: The role of fetal therapy. Best Pract. Res. Clin. Obstet. Gynaecol. 2008, 22, 139–158. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 14) | FGR (n = 10) | FGR + Tadalafil (n = 12) | |
|---|---|---|---|
| Maternal age (year) | 32.5 ± 1.0 | 35.8 ± 0.8 | 32.9 ± 1.2 |
| Primiparity | 7 (50%) | 6 (60%) | 5 (45%) |
| Gestational age at birth (week) | 38.1 ± 0.4 | 35.3 ± 0.5 | 36.7 ± 0.6 |
| Early onset FGR | - | 10 (100%) | 12 (100%) |
| Maternal complication | 0 (0%) | 0 (0%) | 0 (0%) |
| Obstetric event without FGR | 0 (0%) | 0 (0%) | 0 (0%) |
| Delivery mode | |||
| Caesarian delivery | 11 (78%) | 8 (80%) | 6 (45%) |
| Vaginal delivery | 3 (22%) | 2 (20%) | 5 (55%) |
| Sex of newborn infant | |||
| Male | 6 (43%) | 5 (50%) | 5 (42%) |
| Female | 8 (57%) | 5 (50%) | 7 (58%) |
| Birth weight (g) | 2955 ± 68 | 1702 ± 80 | 1903 ± 96 |
| Standard Deviation of birth weight | 0.1 ± 0.5 | −1.9 ± 0.3 | −1.8 ± 0.3 |
| Placental weight (g) | 491 ± 21 | 322 ± 25 | 417 ± 23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuchiya, K.; Tanaka, K.; Tanaka, H.; Maki, S.; Enomoto, N.; Takakura, S.; Nii, M.; Toriyabe, K.; Katsuragi, S.; Ikeda, T. Tadalafil Treatment Ameliorates Hypoxia and Alters Placental Expression of Proteins Downstream of mTOR Signaling in Fetal Growth Restriction. Medicina 2020, 56, 722. https://doi.org/10.3390/medicina56120722
Tsuchiya K, Tanaka K, Tanaka H, Maki S, Enomoto N, Takakura S, Nii M, Toriyabe K, Katsuragi S, Ikeda T. Tadalafil Treatment Ameliorates Hypoxia and Alters Placental Expression of Proteins Downstream of mTOR Signaling in Fetal Growth Restriction. Medicina. 2020; 56(12):722. https://doi.org/10.3390/medicina56120722
Chicago/Turabian StyleTsuchiya, Kyoka, Kayo Tanaka, Hiroaki Tanaka, Shintaro Maki, Naosuke Enomoto, Sho Takakura, Masafumi Nii, Kuniaki Toriyabe, Shinji Katsuragi, and Tomoaki Ikeda. 2020. "Tadalafil Treatment Ameliorates Hypoxia and Alters Placental Expression of Proteins Downstream of mTOR Signaling in Fetal Growth Restriction" Medicina 56, no. 12: 722. https://doi.org/10.3390/medicina56120722
APA StyleTsuchiya, K., Tanaka, K., Tanaka, H., Maki, S., Enomoto, N., Takakura, S., Nii, M., Toriyabe, K., Katsuragi, S., & Ikeda, T. (2020). Tadalafil Treatment Ameliorates Hypoxia and Alters Placental Expression of Proteins Downstream of mTOR Signaling in Fetal Growth Restriction. Medicina, 56(12), 722. https://doi.org/10.3390/medicina56120722