Complement Properdin Determines Disease Activity in MRL/lpr Mice

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Properdin Deficient Fas Mutant Mice
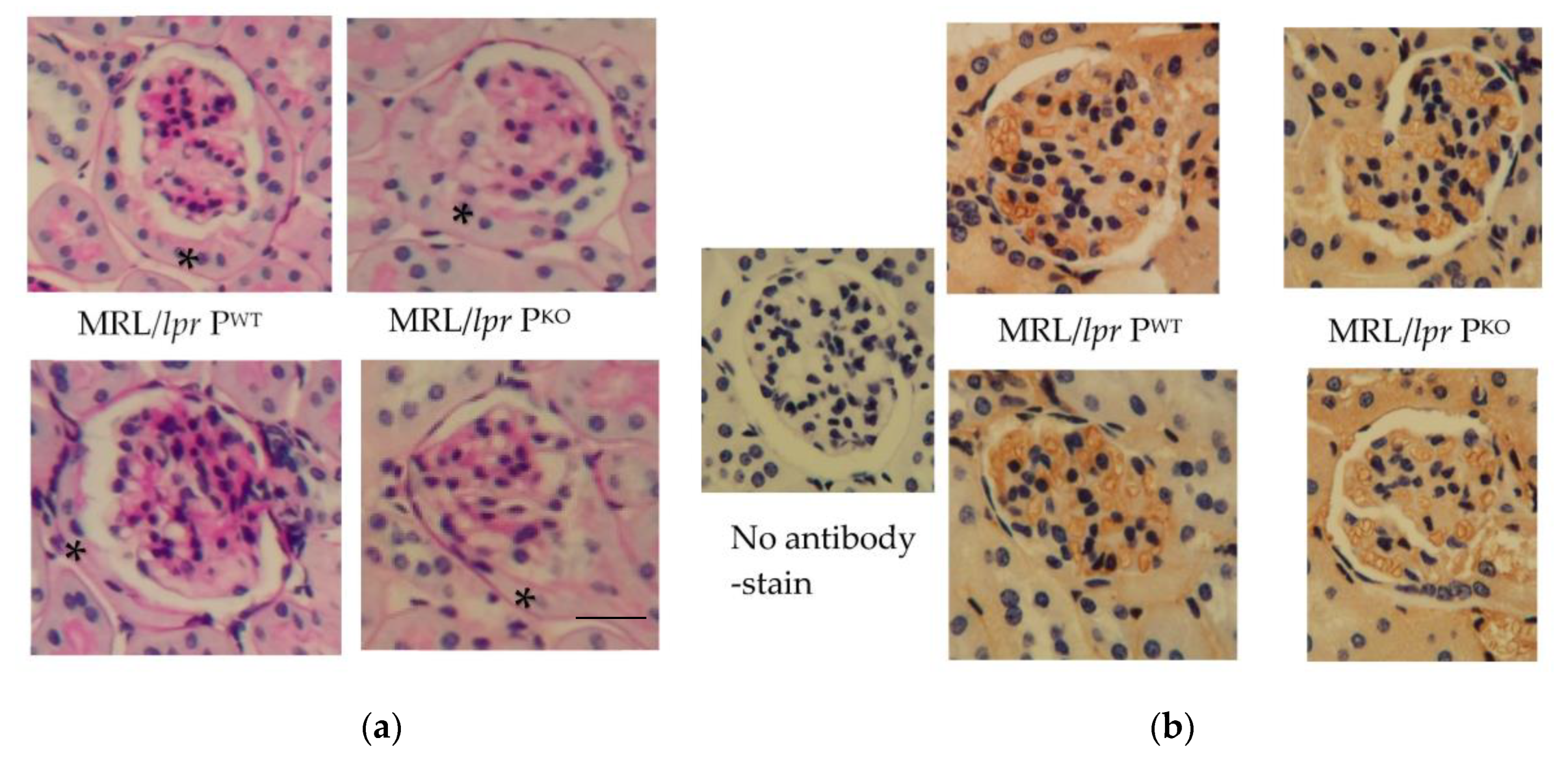
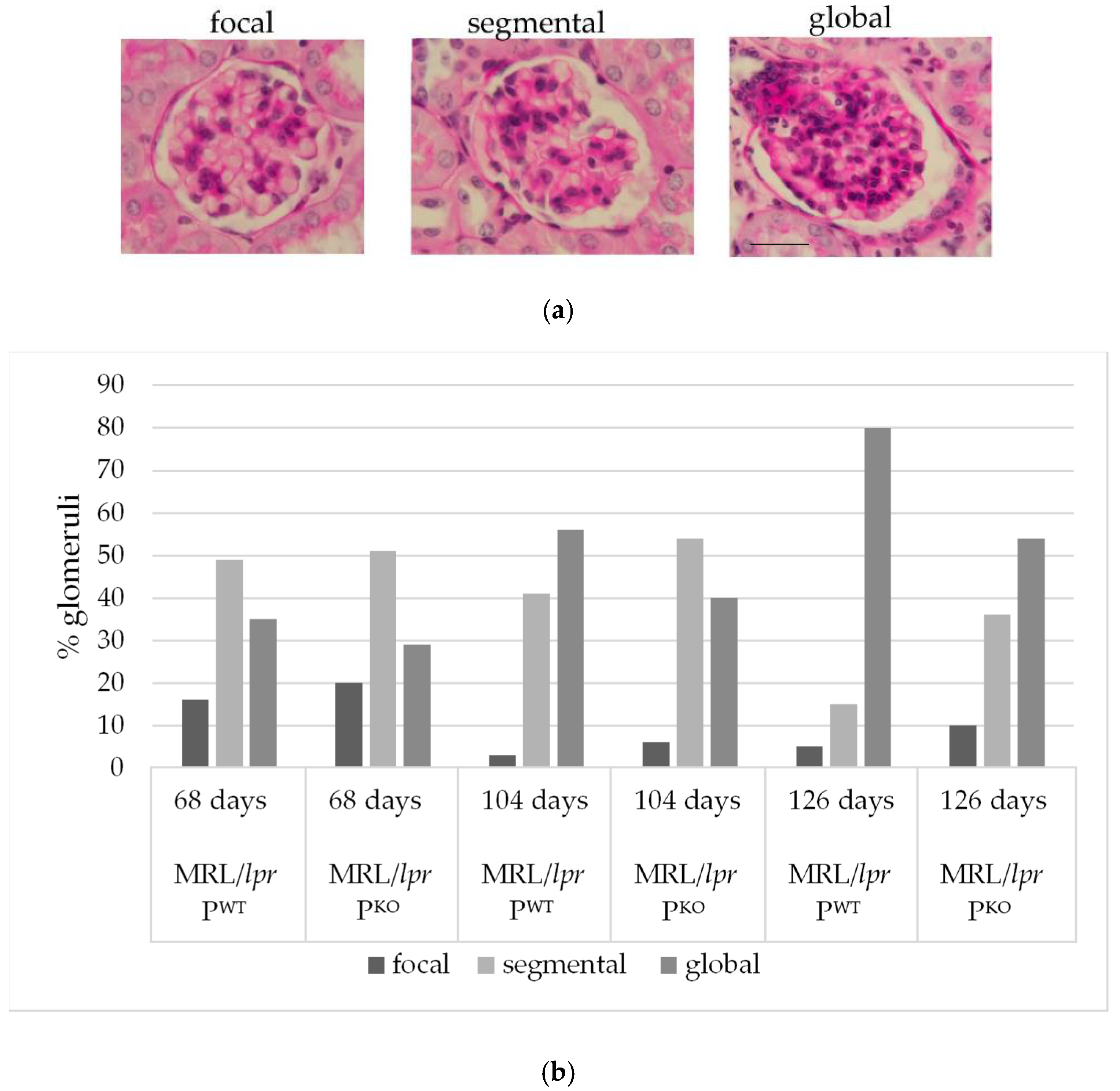
2.2. Histopathology
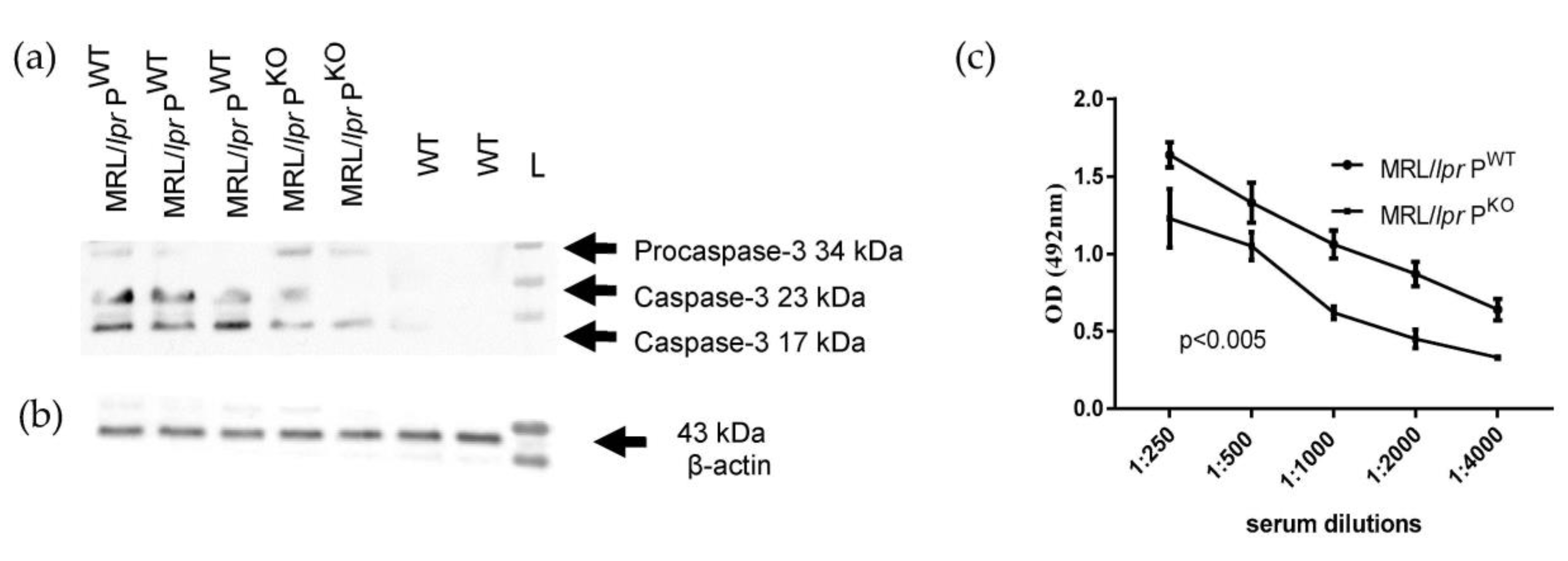
2.3. Anti-DNA ELISA
2.4. C9 Functional Complements ELISAs
2.5. BAFF and TNF-α ELISAs
2.6. Serum Creatinine
2.7. Caspase-3 Western Blot
2.8. Statistical Analysis
3. Results
3.1. Assessment of Renal Histopathology
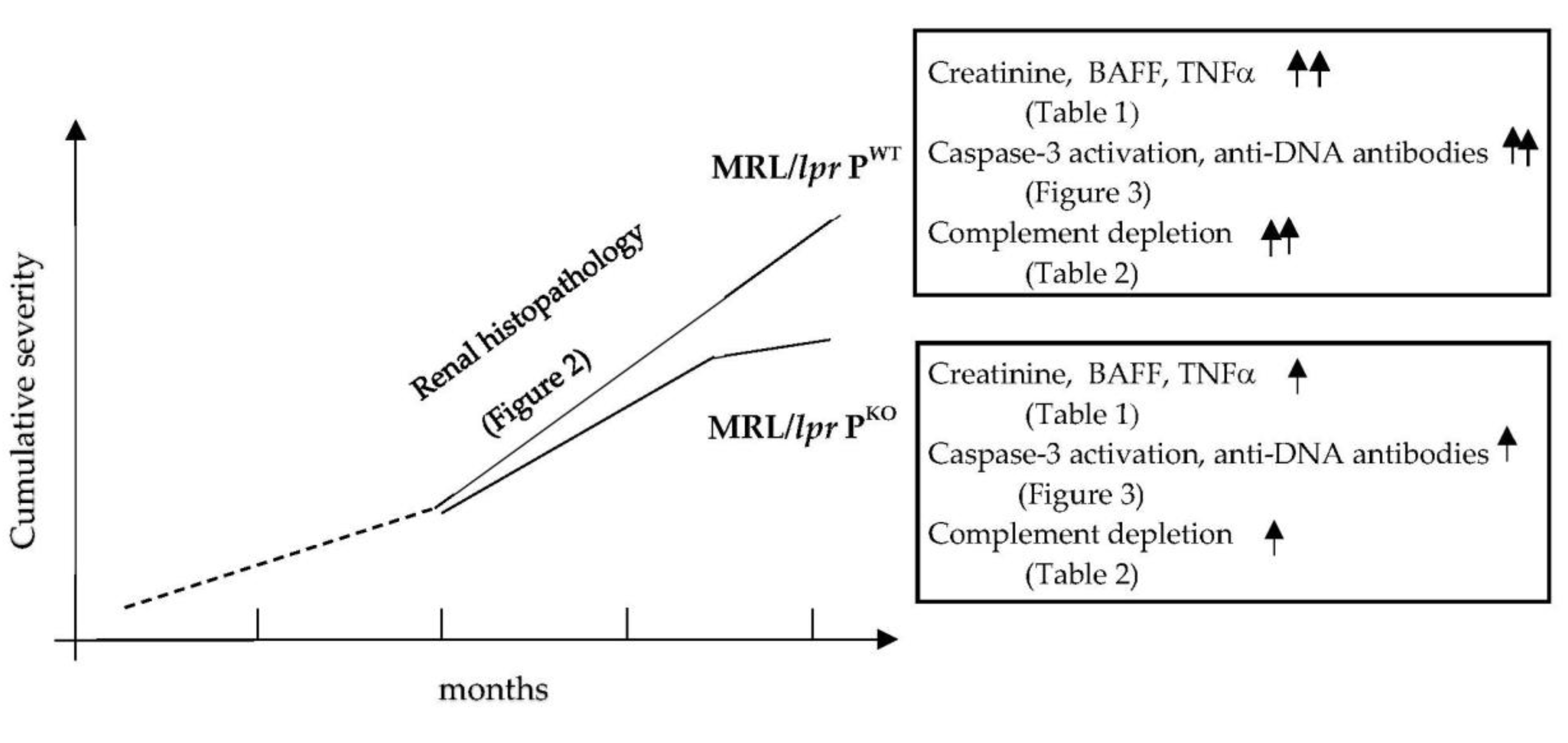
3.2. Assessment of Disease Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part I—Molecular Mechanisms of Activation and Regulation. Front. Immunol. 2015, 6, 262. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, A.K.; Moore, T.L. Presence of plasma complement regulatory proteins clusterin (Apo J) and vitronectin (S40) on circulating immune complexes (CIC). Clin. Exp. Immunol. 2006, 145, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Pisetsky, D.S. Anti-DNA antibodies—Quintessential biomarkers of SLE. Nat. Rev. Rheumatol. 2016, 12, 102–110. [Google Scholar] [CrossRef] [PubMed]
- De Amorim, J.C.; Frittoli, R.B.; Pereira, D.; Postal, M.; Dertkigil, S.S.J.; Reis, F.; Costallat, L.T.; Appenzeller, S. Epidemiology, characterization and diagnosis of neuropsychiatric events in systemic lupus erythematosus. Expert Rev. Clin. Immunol. 2019, 15, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Black, C.A. Delayed type hypersensitivity: Current theories with an historic perspective. Dermatol. Online J. 1999, 5, 7. [Google Scholar]
- Sturfelt, G.; Truedsson, L. Complement and its breakdown products in SLE. Rheumatology 2005, 44, 1227–1232. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.A.; Tian, C.; Taylor, K.E.; Lee, A.T.; Ortmann, W.A.; Hom, G.; Graham, R.R.; Nititham, J.; Kelly, J.A.; Morrisey, J.; et al. European population substructure is associated with mucocutaneous manifestations and autoantibody production in systemic lupus erythematosus. Arthritis Rheum. 2009, 60, 2448–2456. [Google Scholar] [CrossRef] [Green Version]
- Tazi Mezalek, Z.; Bono, W. Challenges for lupus management in emerging countries. Presse Med. 2014, 43, e209–e220. [Google Scholar] [CrossRef]
- Hochberg, M.C.; Boyd, R.E.; Ahearn, J.M.; Arnett, F.C.; Bias, W.B.; Provost, T.T.; Stevens, M.B. Systemic lupus erythematosus: A review of clinico-laboratory features and immunogenetic markers in 150 patients with emphasis on demographic subsets. Medicine 1985, 64, 285–295. [Google Scholar] [CrossRef]
- Manzi, S.; Rairie, J.E.; Carpenter, A.B.; Kelly, R.H.; Jagarlapudi, S.P.; Sereika, S.M.; Medsger, T.A., Jr.; Ramsey-Goldman, R. Sensitivity and specificity of plasma and urine complement split products as indicators of lupus disease activity. Arthritis Rheum. 1996, 39, 1178–1788. [Google Scholar] [CrossRef]
- Merle, N.S.; Noe, R.; Halbwachs-Mecarelli, L.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement System Part II: Role in Immunity. Front. Immunol. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Berger, S.P.; Trouw, L.A.; de Boer, H.C.; Schlagwein, N.; Mutsaers, C.; Daha, M.R.; van Kooten, C. Properdin binds to late apoptotic and necrotic cells independently of C3b and regulates alternative pathway complement activation. J. Immunol. 2008, 180, 7613–7621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrin, L.H.; Lambert, P.H.; Miescher, P.A. Properdin levels in systemic lupus erythematosus and membranoproliferative glomerulonephritis. Clin. Exp. Immunol. 1974, 16, 575–581. [Google Scholar] [PubMed]
- Harboe, M.; Ulvund, G.; Vien, L.; Fung, M.; Mollnes, T.E. The quantitative role of alternative pathway amplification in classical pathway induced terminal complement activation. Clin. Exp. Immunol. 2004, 138, 439–446. [Google Scholar] [CrossRef]
- Dupont, A.; Mohamed, F.; Salehen, N.; Glenn, S.; Francescut, L.; Adib, R.; Byrne, S.; Brewin, H.; Elliott, I.; Richards, L.; et al. Septicaemia models using Streptococcus pneumoniae and Listeria monocytogenes: Understanding the role of complement properdin. Med. Microbiol. Immunol. 2014, 203, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Zwaini, Z.; Dai, H.; Stover, C.; Yang, B. Role of Complement Properdin in Renal Ischemia-Reperfusion Injury. Curr. Gene Ther. 2017, 17, 411–423. [Google Scholar] [CrossRef]
- Watanabe, H.; Garnier, G.; Circolo, A.; Wetsel, R.A.; Ruiz, P.; Holers, V.M.; Boackle, S.A.; Colten, H.R.; Gilkeson, G.S. Modulation of renal disease in MRL/lpr mice genetically deficient in the alternative complement pathway factor B. J. Immunol. 2000, 164, 786–794. [Google Scholar] [CrossRef] [Green Version]
- Elliott, M.K.; Jarmi, T.; Ruiz, P.; Xu, Y.; Holers, V.M.; Gilkeson, G.S. Effects of complement factor D deficiency on the renal disease of MRL/lpr mice. Kidney Int. 2004, 65, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, F.M.M.; Stover, C.M. Characterisation of the properdin-deficient immune phenotype. Curr. Trends Immunol. 2018, 19, 83–96. [Google Scholar]
- Stover, C.M.; Luckett, J.C.; Echtenacher, B.; Dupont, A.; Figgitt, S.E.; Brown, J.; Männel, D.N.; Schwaeble, W.J. Properdin plays a protective role in polymicrobial septic peritonitis. J. Immunol. 2008, 180, 3313–3318. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Xu, J.; Madaio, M.P.; Peng, Q.; Zhang, J.; Grewal, I.S.; Flavell, R.A.; Craft, J. Autoimmune lpr/lpr mice deficient in CD40 ligand: Spontaneous Ig class switching with dichotomy of autoantibody responses. J. Immunol. 1996, 157, 417–426. [Google Scholar] [PubMed]
- Seery, J.P.; Carroll, J.M.; Cattell, V.; Watt, F.M. Antinuclear autoantibodies and lupus nephritis in transgenic mice expressing interferon gamma in the epidermis. J. Exp. Med. 1997, 186, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Kotimaa, J.P.; van Werkhoven, M.B.; O’Flynn, J.; Klar-Mohamad, N.; van Groningen, J.; Schilders, G.; Rutjes, H.; Daha, M.R.; Seelen, M.A.; van Kooten, C. Functional assessment of mouse complement pathway activities and quantification of C3b/C3c/iC3b in an experimental model of mouse renal ischaemia/reperfusion injury. J. Immunol. Methods 2015, 419, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarfe, L.; Rak-Raszewska, A.; Geraci, S.; Darssan, D.; Sharkey, J.; Huang, J.; Burton, N.C.; Mason, D.; Ranjzad, P.; Kenny, S.; et al. Measures of kidney function by minimally invasive techniques correlate with histological glomerular damage in SCID mice with adriamycin-induced nephropathy. Sci. Rep. 2015, 5, 13601. [Google Scholar] [CrossRef] [Green Version]
- Sanz, I.; Lee, F.E. B cells as therapeutic targets in SLE. Nat. Rev. Rheumatol. 2010, 6, 326–337. [Google Scholar] [CrossRef]
- Aringer, M.; Smolen, J.S. The role of tumor necrosis factor-alpha in systemic lupus erythematosus. Arthritis Res. Ther. 2008, 10, 202. [Google Scholar] [CrossRef] [Green Version]
- Miossec, C.; Dutilleul, V.; Fassy, F.; Diu-Hercend, A. Evidence for CPP32 activation in the absence of apoptosis during T lymphocyte stimulation. J. Biol. Chem. 1997, 272, 13459–13462. [Google Scholar] [CrossRef] [Green Version]
- Dimitrova, P.; Ivanovska, N.; Belenska, L.; Milanova, V.; Schwaeble, W.; Stover, C. Abrogated RANKL expression in properdin-deficient mice is associated with better outcome from collagen-antibody-induced arthritis. Arthritis Res. Ther. 2012, 14, R173. [Google Scholar] [CrossRef] [Green Version]
- Machida, T.; Sakamoto, N.; Ishida, Y.; Takahashi, M.; Fujita, T.; Sekine, H. Essential Roles for Mannose-Binding Lectin-Associated Serine Protease-1/3 in the Development of Lupus-Like Glomerulonephritis in MRL/lpr Mice. Front. Immunol. 2018, 9, 1191. [Google Scholar] [CrossRef] [Green Version]
- Lesher, A.M.; Nilsson, B.; Song, W.C. Properdin in complement activation and tissue injury. Mol. Immunol. 2013, 56, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Sekine, H.; Kinser, T.T.; Qiao, F.; Martinez, E.; Paulling, E.; Ruiz, P.; Gilkeson, G.S.; Tomlinson, S. The benefit of targeted and selective inhibition of the alternative complement pathway for modulating autoimmunity and renal disease in MRL/lpr mice. Arthritis Rheum. 2011, 63, 1076–1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulik, L.; Hewitt, F.B.; Willis, V.C.; Rodriguez, R.; Tomlinson, S.; Holers, V.M. A new mouse anti-mouse complement receptor type 2 and 1 (CR2/CR1) monoclonal antibody as a tool to study receptor involvement in chronic models of immune responses and disease. Mol. Immunol. 2015, 63, 479–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulik, L.; Laskowski, J.; Renner, B.; Woolaver, R.; Zhang, L.; Lyubchenko, T.; You, Z.; Thurman, J.M.; Holers, V.M. Targeting the Immune Complex-Bound Complement C3d Ligand as a Novel Therapy for Lupus. J. Immunol. 2019, 203, 3136–3147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bröker, K.; Figge, J.; Magnusen, A.F.; Manz, R.A.; Köhl, J.; Karsten, C.M. A Novel Role for C5a in B-1 Cell Homeostasis. Front. Immunol. 2018, 9, 258. [Google Scholar] [CrossRef] [Green Version]
- Kulik, L.; Laskowski, J.; Thurman, J.; Holers, V. Treatment of MRL/lpr mice with a mAb blocking binding of C3d to its receptors decreases anti-DNA autoantibodies and proteinuria: Support for targeting the CR2:C3d interaction as a therapeutic strategy in SLE. Lupus 2016, 3 (Suppl. S1), A1–A80. [Google Scholar]
- Jacob, A.; Hack, B.; Chiang, E.; Garcia, J.G.; Quigg, R.J.; Alexander, J.J. C5a alters blood-brain barrier integrity in experimental lupus. FASEB J. 2010, 24, 1682–1688. [Google Scholar] [CrossRef] [Green Version]
- Schwarting, A.; Relle, M.; Meineck, M.; Föhr, B.; Triantafyllias, K.; Weinmann, A.; Roth, W.; Weinmann-Menke, J. Renal tubular epithelial cell-derived BAFF expression mediates kidney damage and correlates with activity of proliferative lupus nephritis in mouse and men. Lupus 2018, 27, 243–256. [Google Scholar] [CrossRef]
- Ueda, Y.; Miwa, T.; Gullipalli, D.; Sato, S.; Ito, D.; Kim, H.; Palmer, M.; Song, W.-C. Blocking Properdin Prevents Complement-Mediated Hemolytic Uremic Syndrome and Systemic Thrombophilia. J. Am. Soc. Nephrol. 2018, 29, 1928–1937. [Google Scholar] [CrossRef] [Green Version]
- Michels, M.A.H.M.; Volokhina, E.B.; van de Kar, N.C.A.J.; Heuvel, L.P.W.J.V.D. The role of properdin in complement-mediated renal diseases: A new player in complement-inhibiting therapy? Pediatr. Nephrol. 2019, 34, 1349–1367. [Google Scholar] [CrossRef] [Green Version]
- Lieberman, L.A.; Mizui, M.; Nalbandian, A.; Bossé, R.; Crispín, J.C.; Tsokos, G.C. Complement receptor of the immunoglobulin superfamily reduces murine lupus nephritis and cutaneous disease. Clin. Immunol. 2015, 160, 286–291. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MRL/lpr PWT n = 8 | MRL/lpr PKO n = 7 | Unpaired t-Test (MRL/lpr PWT cp MRL/lpr PKO) | |
|---|---|---|---|
| Creatinine (mg/dL) (baseline (4 WT)): 0.29 ± 0.05 | 0.82 ± 0.08 | 0.49 ± 0.03 | p < 0.0001 |
| BAFF (pg/mL) (baseline (4 WT)): 31.17 ± 2.32) | 70.5 ± 3.42 | 55.71 ±3.73 | p < 0.0001 |
| TNFα (pg/mL) (baseline (4 WT)): 7.1 ± 1.8 | 43.05 ± 3.21 | 18.46 ± 2.77 | p < 0.0001 |
| MRL/lpr PWT | MRL/lpr PKO | |
|---|---|---|
| Classical Pathway (Purified IgM) | 48% | 66% |
| Lectin Pathway (Mannan from Saccharomyces cerevisiae) | 37% | 48% |
| Alternative Pathway (Salmonella enteritidis LPS) | 25% | 1% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alaridhee, H.; Alharbi, A.; Saeed, Z.; Thomas, R.C.; Stover, C.M. Complement Properdin Determines Disease Activity in MRL/lpr Mice. Medicina 2020, 56, 430. https://doi.org/10.3390/medicina56090430
Alaridhee H, Alharbi A, Saeed Z, Thomas RC, Stover CM. Complement Properdin Determines Disease Activity in MRL/lpr Mice. Medicina. 2020; 56(9):430. https://doi.org/10.3390/medicina56090430
Chicago/Turabian StyleAlaridhee, Hasanain, Azzah Alharbi, Zeayd Saeed, Róisín C. Thomas, and Cordula M. Stover. 2020. "Complement Properdin Determines Disease Activity in MRL/lpr Mice" Medicina 56, no. 9: 430. https://doi.org/10.3390/medicina56090430
APA StyleAlaridhee, H., Alharbi, A., Saeed, Z., Thomas, R. C., & Stover, C. M. (2020). Complement Properdin Determines Disease Activity in MRL/lpr Mice. Medicina, 56(9), 430. https://doi.org/10.3390/medicina56090430