A Sulfated-Polysaccharide Fraction from Seaweed Gracilaria birdiae Prevents Naproxen-Induced Gastrointestinal Damage in Rats


Abstract
:1. Introduction
2. Results and Discussion


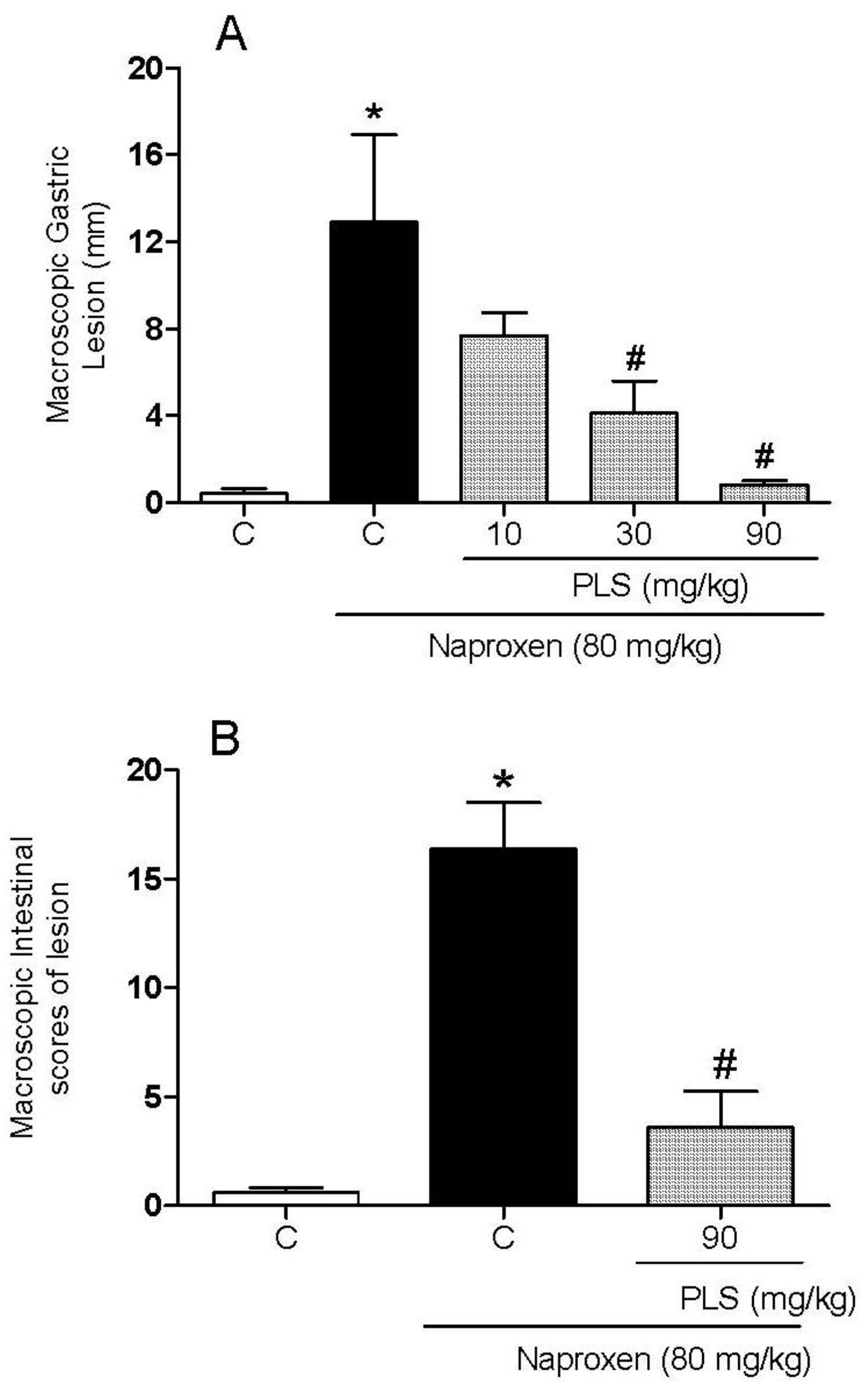{kind=link}
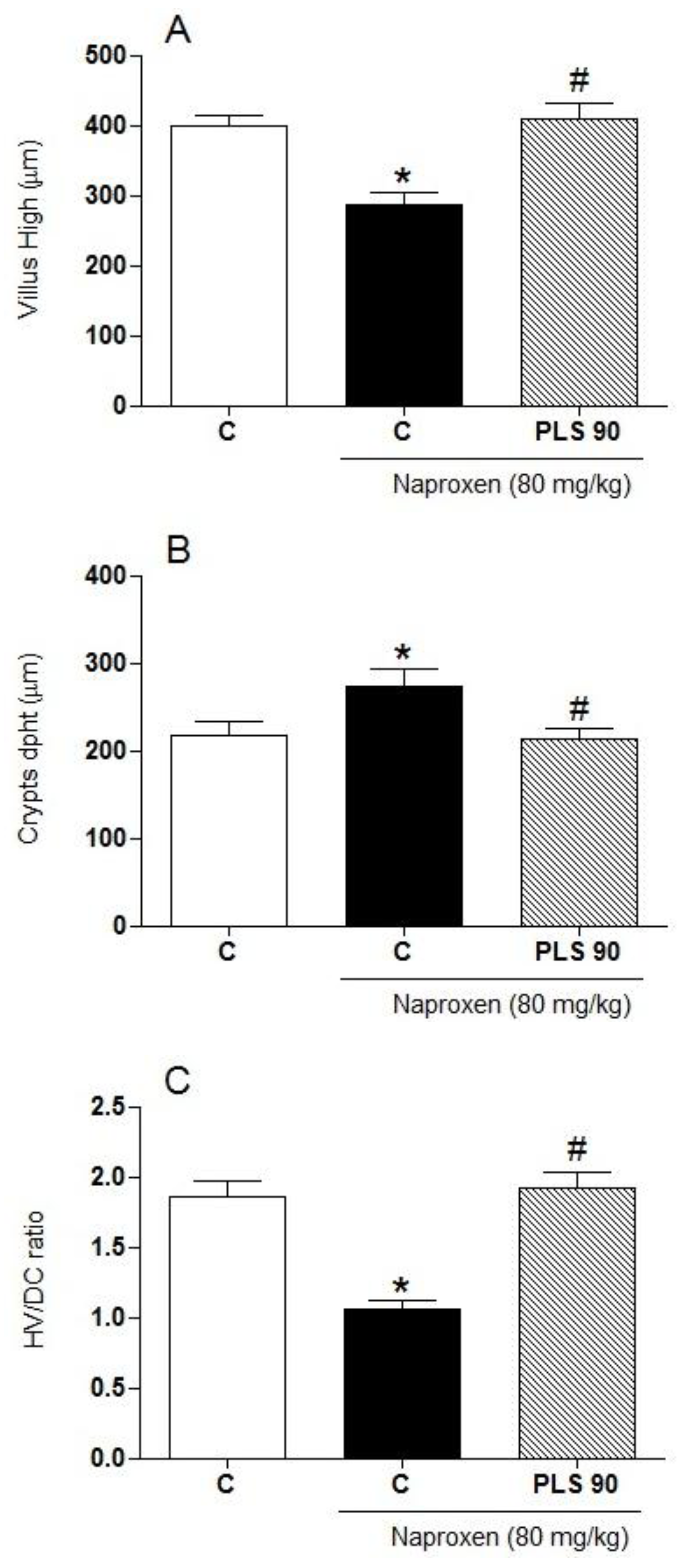{kind=link}
{kind=link}
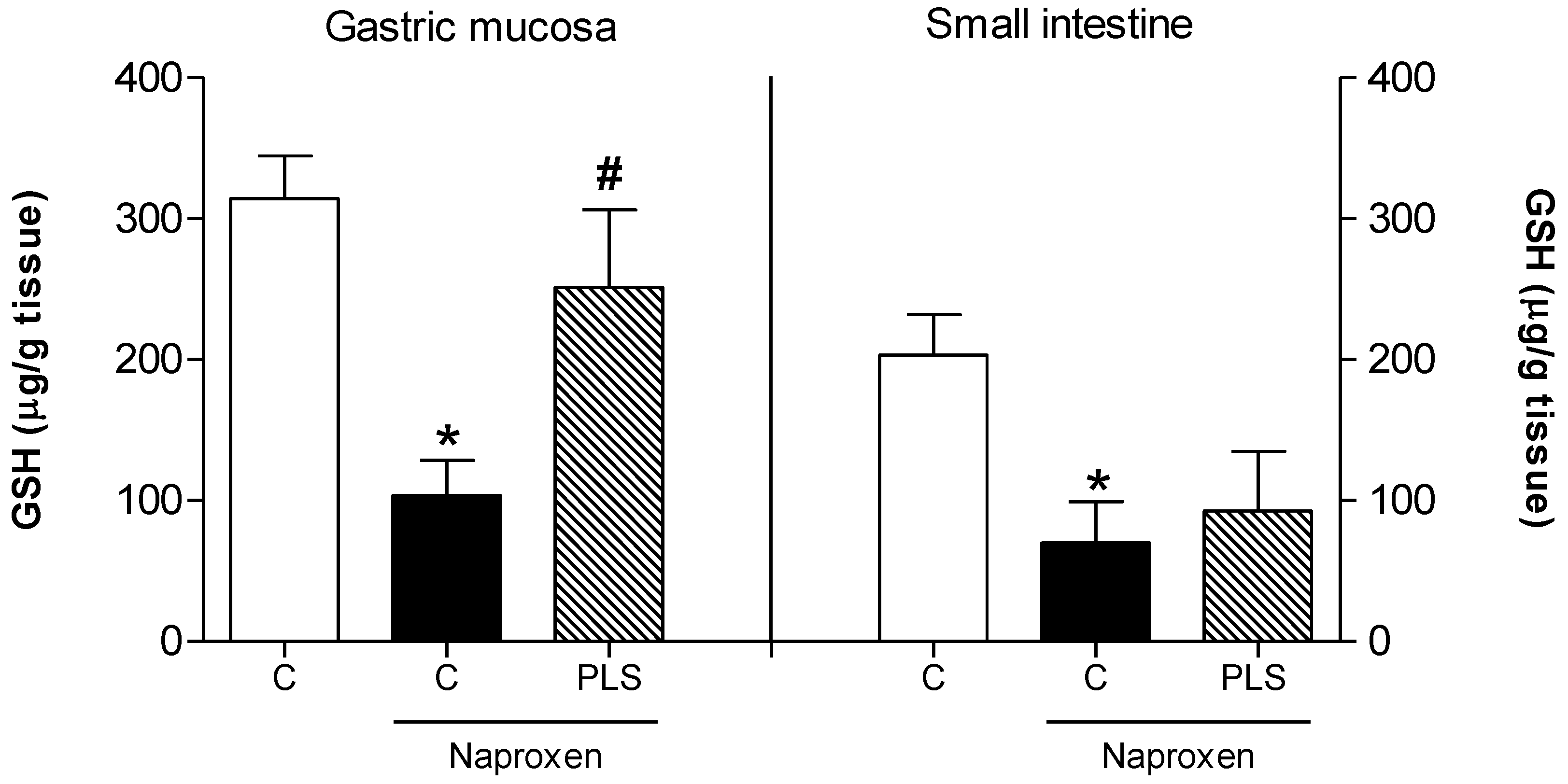{kind=link}
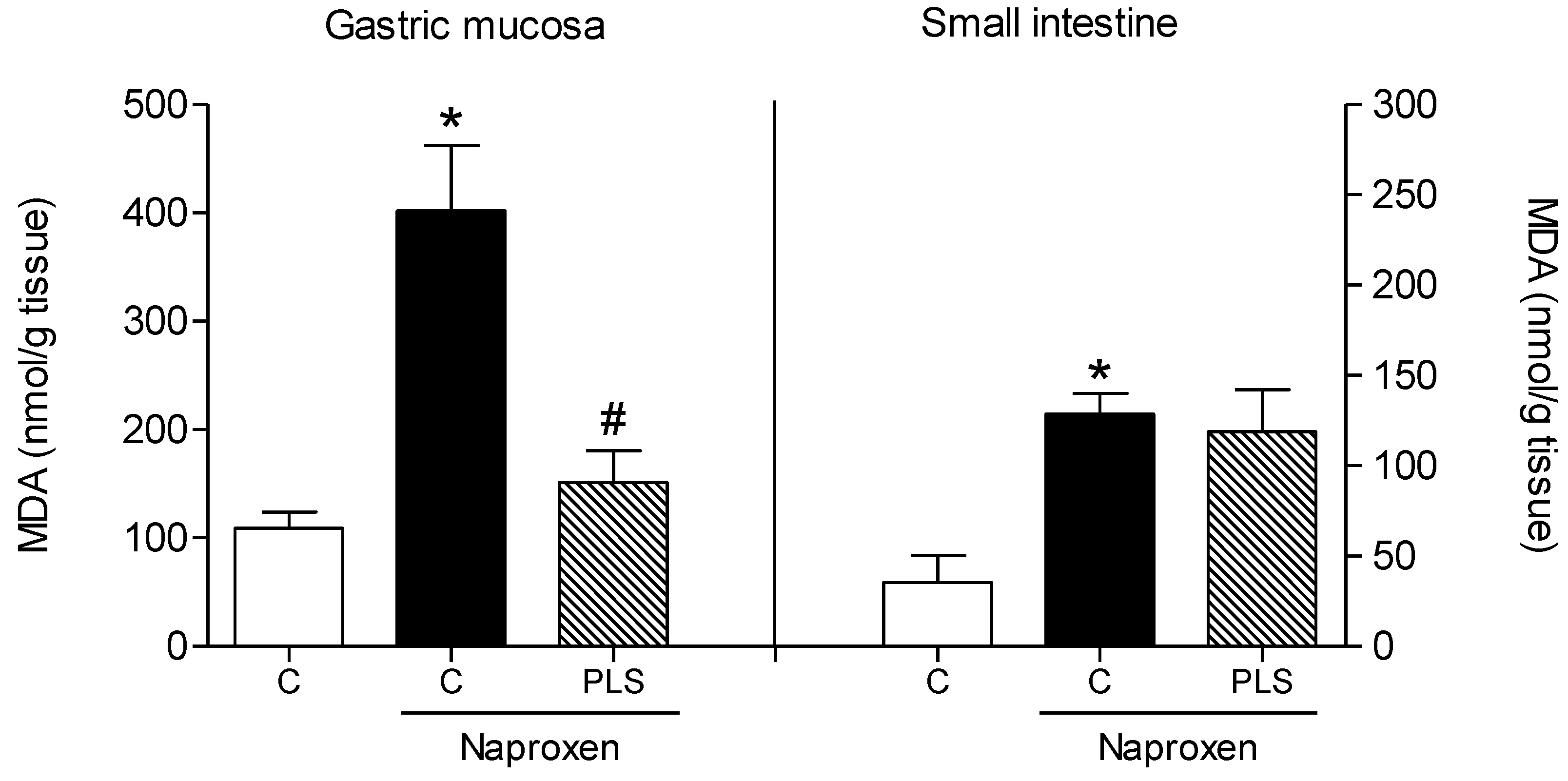{kind=link}
| Experimental group (n = 5) | Hemorrhagic damage (score, 0–4) | Edema (score, 0–4) | Epithelial cell loss (score, 0–3) | Inflammatory cells (score, 0–3) | Total (score, 0–14) |
|---|---|---|---|---|---|
| Control | 0 | 0 (0–1) | 0 | 0 (0–1) | 0 (0–3) |
| Naproxen | 0 | 3 (1–3) a | 3 (3–3) a | 2 (2–3) a | 7 (5–9) a |
| Naproxen + PLS (90 mg/kg) | 0 | 0 (0–1) b | 0 (0–2) b | 0 (0–1) b | 0 (0–3) b |




3. Experimental Section
3.1. Extraction of the Sulfated Polysaccharide Fraction
3.2. Chemical Characterization of the Sulfated Polysaccharide Fraction
3.3. Animals
3.4. Effect of PLS on Naproxen-Induced Gastric Damage
3.5. Effect of PLS on Naproxen-Induced Small Intestine Damage
| Scores | Criteria |
|---|---|
| 0 | No damage. |
| 1 | Focal hyperemia; no ulcers. |
| 2 | Ulceration without hyperemia or bowel wall thickening. |
| 3 | Ulceration with inflammation at 1 site. |
| 4 | ≥2 Sites of ulceration and inflammation. |
| 5 | ≥2 Major sites of ulceration and inflammation or 1 site of ulceration extending >1 cm along length of intestinal. |
| 6–10 | If damage covered >2 cm along length of intestinal, score is increased by 1 for each additional cm of involvement. |
3.6. Histological Evaluation of Gastric Damage
3.7. Intestinal Morphometric Analysis
3.8. Glutathione Levels
3.9. Malondialdehyde Concentration
3.10. Myeloperoxidase Activity
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Qi, H.; Zhao, T.; Zhang, Q.; Li, Z.; Zhao, Z. Antioxidant activity of different molecular weight sulfated polysaccharides from Ulva pertusa Kjellm (Chlorophyta). J. Appl. Phycol. 2005, 17, 527–534. [Google Scholar] [CrossRef]
- Yang, Y.F.; Fei, X.G.; Song, J.M.; Hu, H.Y.; Wang, G.C.; Chung, I.K. Growth of Gracilaria lemaneiformis under different cultivation conditions and its effects on nutrient removal in Chinese coastal waters. Aquaculture 2006, 254, 248–255. [Google Scholar] [CrossRef]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar]
- Almeida-Lima, J.; Costa, L.S.; Silva, N.B.; Melo-Silveira, R.F.; Silva, F.V.; Felipe, M.B.M.C.; Medeiros, S.R.B.M.; Leite, E.L.; Rocha, H.A.O. Evaluating the possible genotoxic, mutagenic and tumor cell proliferation-inhibition effects of a non-anticoagulant, but antithrombotic algal heterofucan. J. Appl. Toxicol. 2010, 30, 708–715. [Google Scholar] [CrossRef]
- 5. Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Câmara, R.B.G.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida-Lima, J.; Farias, E.H.C. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’incecco, A.; Picooli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar]
- Chaves, L.S.; Nicolau, L.A.D.; Silva, R.O.; Barros, F.C.N.; Freitas, A.L.P.; Aragao, K.S.; Ribeiro, R.A.; Souza, M.H.L.P.; Barbosa, A.L.R.; Medeiros, J.V.R. Anti-inflammatory and anti-nociceptive effects in mice of a sulfated polysaccharide fraction extracted from the marine red algae Gracilaria caudata. Immunopharmacol. Immunotoxicol. 2012, in press. [Google Scholar]
- Knutsen, S.H.; Myslabodski, D.E.; Larsen, B.; Usov, A.I. A modified system of nomenclature for red algal galactans. Bot. Mar. 1994, 37, 163–170. [Google Scholar]
- Rees, D.A. Biogenesis of 3,6-anhydro-l-galactose. Biochem. J. 1961, 81, 347–352. [Google Scholar]
- Ye, H.; Wang, K.; Zhou, C.; Liu, J.; Zeng, X. Purification, antitumor and antioxidant activities in vitro of polysaccharides from the brown seaweed Sargassum pallidum. Food Chem. 2008, 111, 428–432. [Google Scholar] [CrossRef]
- 11. Souza, B.W.S.; Cerqueira, M.A.; Bourbon, A.I.; Pinheiro, A.C.; Martins, J.T.; Teixeira, J.A.; Coimbra, M.A.; Vicente, A.A. Chemical characterization and antioxidant activity of sulfated polysaccharide from the red seaweed Gracilaria birdiae. Food Hydrocoll. 2012, 27, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.O.; Santos, G.M.P.; Nicolau, L.A.D.; Lucetti, L.T.; Santana, A.P.M.; Chaves, L.S.C.; Barros, C.N.; Freitas, A.L.P.; Souza, M.H.L.P.; Medeiros, J.V.R. Sulfated-polysaccharide fraction from red algae Gracilaria caudata protects mice gut against ethanol-induced gastric damage. Mar. Drugs 2011, 9, 2188–2200. [Google Scholar] [CrossRef]
- Fries, J.F. NSAID gastropathy: Epidemiology. J. Musculoskelet. Med. 1991, 8, 21–28. [Google Scholar]
- Villegas, I.; La Casa, C.; De la Lastra, C.A.; Motilva, V.; Herrerías, J.M.; Martín, M.J. Mucosal damage induced by preferential COX-1 and COX-2 inhibitors: Role of prostaglandins and inflammatory response. Life Sci. 2004, 74, 873–884. [Google Scholar] [CrossRef]
- Beck, W.S.; Schneider, H.T.; Dietzel, K.; Nuernberg, B.; Brune, K. Gastrointestinal ulcerations induced by anti-inflammatory drugs in rats. Arch. Toxicol. 1990, 64, 210–217. [Google Scholar] [CrossRef]
- Tenembaum, J. The epidemiology of nonsteroidal anti-inflammatory drugs. Can. J. Gastroenterol. 1999, 13, 119–122. [Google Scholar]
- Kim, J.H.; Kim, Y.S.; Song, G.G.; Park, J.J.; Chang, H.I. Protective effect of astaxanthin on naproxen-induced gastric antral ulceration in rats. Eur. J. Pharmacol. 2005, 514, 53–59. [Google Scholar] [CrossRef]
- Parks, D.A. Oxygen radicals: Mediators of gastrointestinal pathophysiology. Gut 1989, 30, 293–298. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Naito, Y.; Ueda, S.; Oyamada, H.; Takemura, T.; Yoshida, N.; Sugino, S.; Kondo, M. Role of oxygen-derived free radicals in the pathogenesis of gastric mucosal lesions in rats. J. Clin. Gastroenterol. 1990, 12, 65–71. [Google Scholar] [CrossRef]
- Maciel, J.S.; Chaves, L.S.; Souza, B.W.S.; Teixeira, D.I.A.; Freitas, A.L.P.; Feitosa, J.P.A.; De Paula, R.C.M. Structural characterization of cold extracted fraction of soluble sulfated polysaccharide from red seaweed Gracilaria birdiae. Carbohydr. Polym. 2008, 71, 559–565. [Google Scholar] [CrossRef]
- Regula, J.; Butruk, E.; Dekkers, C.P.; de Boer, S.Y.; Raps, D.; Simon, L.; Terjung, A.; Thomas, K.B.; Luhmann, R.; Fischer, R. Prevention of NSAID-associated gastrointestinal lesions: A comparison study pantoprazole versus omeprazole. Am. J. Gastroenterol. 2006, 101, 1747–1755. [Google Scholar] [CrossRef]
- Nishida, T.; Tsujii, M.; Tsujii, S. Are COX-2 inhibitors truly able to prevent NSAIDs-associated ulcers? Nippon Rinsho 2004, 62, 561–565. [Google Scholar]
- Wallace, J.L. NSAID gastroenterophaty: Past, present and future. Can. J. Gastroenterol. 1996, 10, 451–459. [Google Scholar]
- Kameda, N.; Higuchi, K.; Shiba, M. A prospective, single-blind trial comparing wireless capsule endoscopy and double-balloon enteroscopy in patients with obscure gastrointestinal bleeding. Gastroenterology 2008, 43, 434–440. [Google Scholar] [CrossRef]
- Pemberton, R.E.; Strand, L.J. A review of upper-gastrointestinal effects of the newer nonsteroidal antiinflammatory agents. Dig. Dis. Sci. 1979, 24, 53–64. [Google Scholar] [CrossRef]
- Higuchi, K.; Umegaki, E.; Watanabe, T.; Yoda, Y.; Morita, E.; Murano, M.; Tokioka, S.; Arakawa, T. Present status and strategy of NSAIDs-induced small bowel injury. J. Gastroenterol. 2009, 44, 879–888. [Google Scholar] [CrossRef]
- Takeuchi, K.; Miyazawa, T.; Tanaka, A.; Kato, S.; Kunikat, T. Pathogenic importance of intestinal hypermotility in NSAID-induced small intestinal damage in rats. Digestion 2002, 66, 30–41. [Google Scholar] [CrossRef]
- Reuter, B.K.; Davies, N.M.; Wallace, J.L. Nonsteroidal anti-inflammatory drug enteropathy in rats: Role of permeability, bacteria and enterohepatic circulation. Gastroenterology 1997, 112, 109–117. [Google Scholar] [CrossRef]
- Chen, D.; Wu, X.Z.; Wen, Z.Y. Sulfated polysaccharides and immune response: Promoter or inhibitor? Panminerva Med. 2008, 50, 177–183. [Google Scholar]
- Suzuki, M.; Mori, M.; Miura, S.; Suematsu, M.; Fukumura, D.; Kimura, H.; Ishii, H. Omeprazole attenuates oxygen-derived free radical production from human neutrophils. Free Radic. Biol. Med. 1996, 21, 727–731. [Google Scholar] [CrossRef]
- Zhang, X.; Tajima, K.; Kageyama, K.; Kyoi, T. Irsogladine maleate suppresses indomethacin-induced elevation of proinflammatory cytokines and gastric injury in rats. World J. Gastroenterol. 2008, 14, 4784–4790. [Google Scholar] [CrossRef]
- Naito, Y.; Yoshikawa, T.; Matsuyama, K.; Yagi, N.; Arai, M.; Nakamura, Y.; Nishimura, S.; Yoshida, N.; Kondo, M. Effects of oxygen radical scavengers on the quality of gastric ulcer healing in rats. J. Clin. Gastroenterol. 1995, 21, 82–86. [Google Scholar] [CrossRef]
- Wallace, J.L. Pathogenesis of NSAID-induced gastroduedenal mucosal injury. Best Pract. Res. Clin. Gastroenterol. 2001, 15, 691–703. [Google Scholar] [CrossRef]
- Odabasoglu, F.; Cakir, A.; Suleyman, H.; Aslan, A.; Bayir, Y.; Halici, M.; Kazaz, C. Gastroprotective and antioxidant effects of usnic acid on indomethacine-induced gastric ulcer in rats. J. Ethnopharmacol. 2006, 103, 59–65. [Google Scholar] [CrossRef]
- Basiviredy, J.; Jacob, M.; Ramamoorthy, P.; Pulimood, A.B.; Balasubramanian, K.A. Indomethacin-induced free radical-mediated changes in the intestinal brush border membranes. Biochem. Pharmacol. 2003, 65, 683–695. [Google Scholar]
- Medeiros, J.V.; Gadelha, G.G.; Lima, S.J.; Garcia, J.A.; Soares, P.M.; Santos, A.A.; Brito, G.A.; Ribeiro, R.A.; Souza, M.H. Role of the NO/cGMP/KATP pathway in the protective effects of sildenafil against ethanol-induced gastric damage in rats. Br. J. Pharmacol. 2008, 153, 721–727. [Google Scholar] [CrossRef]
- Das, D.; Bandyopadhyay, D.; Bhattacharjee, M.; Banerjee, R.K. Hydroxyl radical is the major cousative factor in stress-induced gastric ulceration. Free Radic. Biol. Med. 1997, 23, 8–18. [Google Scholar] [CrossRef]
- Bradley, P.P.; Christensen, R.D.; Rothstein, G. Cellular and extracellular myeloperoxidase in pyogenic inflammation. Blood 1982, 60, 618–622. [Google Scholar]
- Cuzzocrea, S.; Riley, D.P.; Caputi, A.P.; Salvemini, D. Antioxidant therapy: A new pharmacological approach in shock, inflammation and ischemia/reperfusion injury. Pharmacol. Rev. 2001, 53, 135–159. [Google Scholar]
- Zhao, B. Natural antioxidants protect neurons in Alzheimer’s disease and Parkinson’s disease. Neurochem. Res. 2009, 34, 630–638. [Google Scholar] [CrossRef]
- Xing, R.; Yu, H.; Liu, S.; Zhang, W.; Zhang, Q.; Li, Z.; Li, P. Antioxidant activity of differently regioselective chitosan sulfates in vitro. Bioorg. Med. Chem. 2005, 13, 1387–1392. [Google Scholar] [CrossRef]
- Farias, W.R.L.; Valente, A.P.; Pereira, M.S.; Mourão, P.A.S. Structure and anticoagulant activity of sulfated galactans. Isolation of a unique sulfated galactan from the red alga Botryocladia occidentalis and comparison of its anticoagulant action with that of sulfated galactans from invertebrates. J. Biol. Chem. 2000, 275, 29299–29307. [Google Scholar]
- Dubois, M.; Gillis, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar]
- Lloyd, A.G.; Dodgson, K.S.; Price, R.G.; Rose, F.A.I. Infrared studies on sulphate esters. I. Polysaccharide sulphates. Biochim. Biophys. Acta 1961, 46, 108–115. [Google Scholar] [CrossRef]
- Stevenson, T.T.; Furneaux, R.H. Chemical methods for the analysis of sulphated galactans from red algae. Carbohydr. Res. 1991, 210, 277–298. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 24, 1992–2005. [Google Scholar]
- Mihara, M.; Uchiyama, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Invest. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef]
- Morris, G.P.; Beck, P.L.; Herridge, M.S.; Depew, W.T.; Szewczuk, M.R.; Wallace, J.L. Hapten-induced model of chronic inflammation and ulceration in the rat colon. Gastroenterology 1989, 96, 795–803. [Google Scholar]
- Laine, L.; Weinstein, W.M. Histology of alcoholic hemorrhagic “gastritis”: A prospective evaluation. Gastroenterology 1988, 94, 1254–1262. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Silva, R.O.; Santana, A.P.M.; Carvalho, N.S.; Bezerra, T.S.; Oliveira, C.B.; Damasceno, S.R.B.; Chaves, L.S.; Freitas, A.L.P.; Soares, P.M.G.; Souza, M.H.L.P.; et al. A Sulfated-Polysaccharide Fraction from Seaweed Gracilaria birdiae Prevents Naproxen-Induced Gastrointestinal Damage in Rats. Mar. Drugs 2012, 10, 2618-2633. https://doi.org/10.3390/md10122618
Silva RO, Santana APM, Carvalho NS, Bezerra TS, Oliveira CB, Damasceno SRB, Chaves LS, Freitas ALP, Soares PMG, Souza MHLP, et al. A Sulfated-Polysaccharide Fraction from Seaweed Gracilaria birdiae Prevents Naproxen-Induced Gastrointestinal Damage in Rats. Marine Drugs. 2012; 10(12):2618-2633. https://doi.org/10.3390/md10122618
Chicago/Turabian StyleSilva, Renan O., Ana Paula M. Santana, Nathalia S. Carvalho, Talita S. Bezerra, Camila B. Oliveira, Samara R. B. Damasceno, Luciano S. Chaves, Ana Lúcia P. Freitas, Pedro M. G. Soares, Marcellus H. L. P. Souza, and et al. 2012. "A Sulfated-Polysaccharide Fraction from Seaweed Gracilaria birdiae Prevents Naproxen-Induced Gastrointestinal Damage in Rats" Marine Drugs 10, no. 12: 2618-2633. https://doi.org/10.3390/md10122618
APA StyleSilva, R. O., Santana, A. P. M., Carvalho, N. S., Bezerra, T. S., Oliveira, C. B., Damasceno, S. R. B., Chaves, L. S., Freitas, A. L. P., Soares, P. M. G., Souza, M. H. L. P., Barbosa, A. L. R., & Medeiros, J. -V. R. (2012). A Sulfated-Polysaccharide Fraction from Seaweed Gracilaria birdiae Prevents Naproxen-Induced Gastrointestinal Damage in Rats. Marine Drugs, 10(12), 2618-2633. https://doi.org/10.3390/md10122618