Ovatoxin-a and Palytoxin Accumulation in Seafood in Relation to Ostreopsis cf. ovata Blooms on the French Mediterranean Coast

 ,
,  ,
,
Abstract
:1. Introduction

2. Results and Discussion
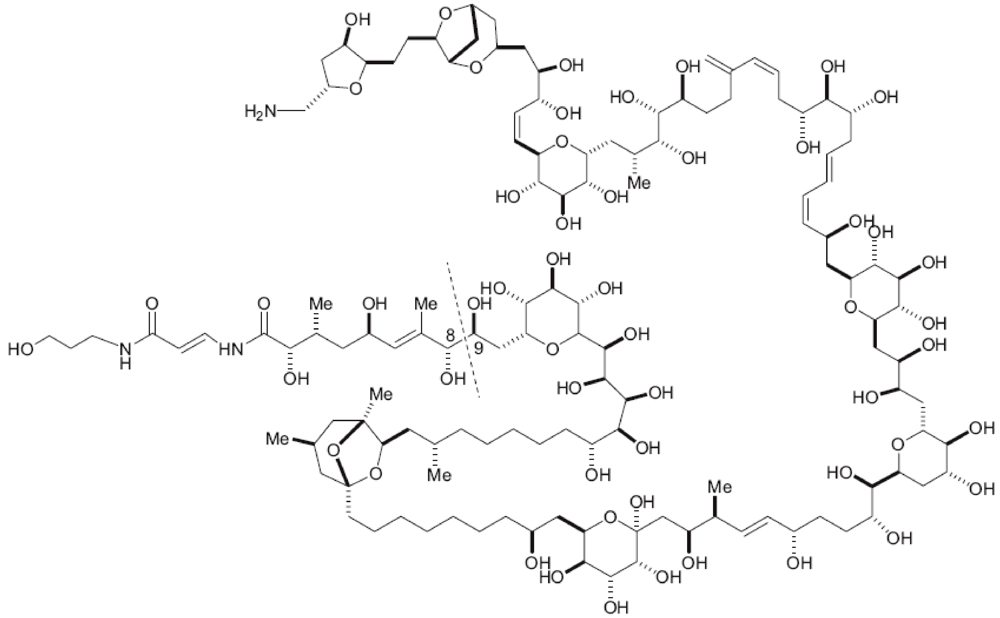
2.1. Optimization of the PLTX Quantification Method by LC-MS/MS
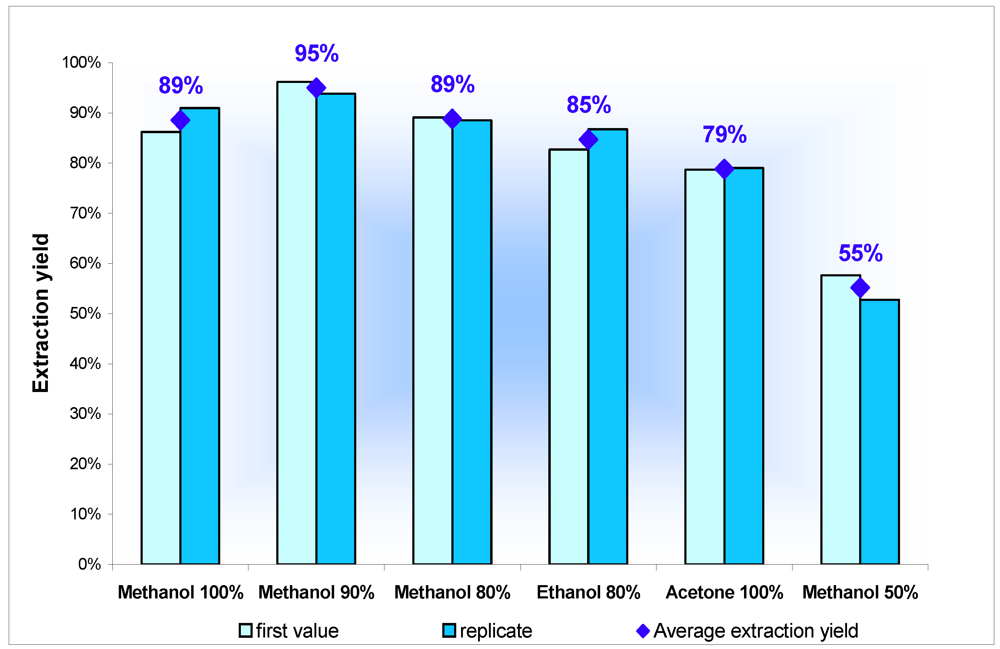
2.1.1. Selection of an Extraction Solvent for PLTX

2.1.2. Specificity of the LC-MS/MS Analytical Procedure

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seafood | Found Quantity (µg/g) | Added Quantity (µg/g) | Recovery |
|---|---|---|---|
| Manila clam | 0.08 | 0.1 | 81% |
| 0.18 | 0.2 | 87% | |
| 0.32 | 0.4 | 77% | |
| Mussels | 0.19 | 0.2 | 95% |
| 0.45 | 0.4 | 112% | |
| 0.68 | 0.8 | 86% | |
| Sea urchin | 0.92 | 0.8 | 115% |
| 1.37 | 1.6 | 85% | |
| 2.4 | 3.2 | 75% | |
| Wedge clam | 0.99 | 1.1 | 90 |
| 1.90 | 22 | 86% | |
| 4.20 | 4.5 | 93% |

2.1.3. Detection and Quantification Limits (LOD and LOQ)
2.2. First Report of the PLTX Group in Marine Organisms in the French Mediterranean

2.3. Genetic Characterization of Ostreopsis Strains

2.4. Culture of Ostreopsis cf. ovata Strains

2.5. In Situ Monitoring of Bioaccumulation of the PLTX Group in Filter-Feeding Shellfish during the Development Ostreopsis cf. ovata in Summer 2009

2.6. Monitoring of Contamination and Detoxification of Mussels and Sea Urchins

3. Conclusions
4. Materials and Methods
4.1. Materials
4.1.1. Reagents and Reference Material
4.1.2. Seafood Samples
4.1.3. Microalgae Samples
4.2. Methods
4.2.1. Cultures of Ostreopsis
4.2.2. Taxonomic Identification by Molecular Analysis
4.2.2.1. DNA Amplification and Sequencing
4.2.2.2. Sequences Alignment and Phylogenetic Analysis
4.2.3. Extraction Procedure for Palytoxin Group Toxins
4.2.3.1. Seafood
4.2.3.2. Ostreopsis Samples
4.2.4. Liquid Chromatography-Multiple Tandem Mass Spectrometry Analysis (LC-MS/MS)
Acknowledgements
References
- Moore, R.E.; Scheuer, P.J. Palytoxin: A new marine toxin from a coelenterate. Science 1971, 172, 495–498. [Google Scholar]
- Moore, R.E.; Bartolini, G. Structure of palytoxin. J. Am. Chem. Soc. 1981, 103, 2491–2494. [Google Scholar]
- Uemura, D. Further studies on palytoxin. II. Structure on palytoxin. Tetrahedron Lett. 1981, 22, 2781–2784. [Google Scholar] [CrossRef]
- Moore, R.E. Structure of palytoxin. Fortschr. Chem. Org. Naturst. 1985, 48, 81–202. [Google Scholar]
- Munday, R. Palytoxin toxicology: Animals studies. Toxicon 2011, 57, 470–477. [Google Scholar]
- Rossini, G.P.; Bigiani, A. Palytoxin action on the Na+, K+-ATPase and the disruption of ion equilibria in biological systems. Toxicon 2011, 57, 429–439. [Google Scholar]
- Usami, M.; Satake, M.; Ishida, S.; Inoue, A.; Yukiko, K.; Yasumoto, T. Palytoxin analogs from the dinoflagellate Ostreopsis siamensis. J. Am. Chem. Soc. 1995, 117, 5389–5390. [Google Scholar]
- Ukena, T.; Satake, M.; Usami, M.; Oshima, Y.; Naoki, H.; Fujita, T.; Kan, Y.; Yasumoto, T. Structure elucidation of Ostreocin D, a palytoxin analog isolated from the dinoflagellate Ostreopsis siamensis. Biosci. Biotechnol. Biochem. 2001, 65, 2585–2588. [Google Scholar]
- Aligizaki, K.; Katikou, P.; Milandri, A.; Diogene, J. Occurence of palytoxin-group toxins in seafood and future strategies to complement the present state of the art. Toxicon 2011, 57, 390–399. [Google Scholar]
- Alcala, A.C.; Alcala, L.C.; Garth, J.S.; Yasumura, D.; Yasumoto, T. Humain fatality due to ingestion of the crac Demanda reynaudii that container a palytoxin-like toxin. Toxicon 1988, 26, 105–107. [Google Scholar]
- Yasumoto, T.; Yasumura, D.; Ohizumi, Y.; Takahashi, M.; Alcala, A.C.; Alcala, L.C. Palytoxin in two species of xanthid crab from the Philippines. Agric. Biol. Chem. 1986, 50, 163–167. [Google Scholar]
- Yasumoto, T. Fish poisoning due to toxins of microalgal origins in the Pacific. Toxicon 1998, 36, 1515–1518. [Google Scholar]
- Mebs, D. Occurrence and sequestration of toxins in food chains. Toxicon 1998, 36, 1519–1522. [Google Scholar]
- Onuma, Y.; Satake, M.; Ukena, T.; Roux, J.; Chanteau, S.; Rasolofonirina, N.; Ratsimaloto, M.; Naoki, H.; Yasumoto, T. Identification of putative palytoxin as the cause of clupeotoxism. Toxicon 1999, 37, 55–65. [Google Scholar] [CrossRef]
- Riobo, P.; Paz, B.; Franco, J.M. Analisis of palytoxin-like in Ostreopsis cultures by liquid chromatography with precolumn derivatization and fluorescente detection. Anal. Chim. Acta 2006, 556, 217–223. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L. A 4-decade-long (and still ongoing) hunt for palytoxins chemical architecture. Toxicon 2011, 57, 362–367. [Google Scholar]
- Riobo, P.; Franco, J.M. Palytoxins: Biological and chemical determination. Toxicon 2011, 57, 368–375. [Google Scholar]
- Rhodes, L.; Towers, N.; Briggs, L.; Munday, R.; Adamson, J. Uptake of palytoxin-like compounds by shellfish fed Ostreopsis siamensis (Dinophyceae). N. Z. J. Mar. Freshw. Res. 2002, 36, 631–636. [Google Scholar]
- Taniyama, S.; Arakawa, O.; Terada, M.; Nishio, S.; Takatani, T.; Mahmud, Y.; Noguchi, T. Ostreopsis sp., a possible origin of palytoxin (PLTX) in parrotfish Scarus ovifrons. Toxicon 2003, 42, 29–33. [Google Scholar]
- Lenoir, S.; Ten-Hage, L.; Turquet, J.; Quod, J.P.; Bernard, C.; Hennion, M.C. First evidence of palytoxin analogues from Ostreopsis mascarensis (Dinophyceae) benthic bloom in southern Indian Ocean. J. Phycol. 2004, 40, 1042–1051. [Google Scholar]
- Vila, M.; Garcés, E.; Masó, M. Potentially toxic epiphytic dinoflagellate assemblages on macroalgae in the NW Mediterranean. Aquat. Microb. Ecol. 2001, 26, 51–60. [Google Scholar]
- Sansoni, G.; Borghini, B.; Camici, G.; Casotti, M.; Righini, P.; Rustighi, C. Fioriture algali di Ostreopsis ovata (Gonyaulacales: Dinophyceae): Un problema emergente. Biol. Ambient. 2003, 17, 17–23. [Google Scholar]
- Aligizaki, K.; Katikou, P.; Nikolaidis, G.; Panou, A. First episode of shellfish contamination by palytoxin-like compounds from Ostreopsis species (Aegean Sea, Greece). Toxicon 2008, 51, 418–427. [Google Scholar]
- Penna, A.; Vila, M.; Fraga, S.; Giacobbe, M.G.; Andreoni, F.; Riobo, P.; Vernesi, C. Characterization of Ostreopsis ovata and Coolia (Dinophyceae) isolates in the Western Mediterranean Sea based on morphological, toxicity and Internal Transcribed Spacer 5.8S rDNA sequences. J. Phycol. 2005, 41, 212–225. [Google Scholar] [CrossRef]
- Turki, S. Distribution of toxic dinoflagellates along the leaves of seagrass Posidonia oceanica and Cymodocea nodosa from the Gulf of Tunis. Cahiers Biol. Mar. 2005, 38, 291–295. [Google Scholar]
- Monti, M.; Minocci, M.; Beran, A.; Ivesa, L. First record of Ostreopsis cf. ovata on macroalgae in the Northern Adriatic Sea. Mar. Pollut. Bull. 2007, 54, 598–601. [Google Scholar] [CrossRef]
- Mangialajo, L.; Bertolotto, R.; Cattaneo-Vietti, R.; Chiantore, M.; Grillo, C.; Lemee, R.; Melchiorre, N.; Moretto, P.; Povero, P.; Ruggieri, N. The toxic benthic dinoflagellate Ostreopsis ovata: Quantification of proliferation along the coastline of Genoa, Italy. Mar. Pollut. Bull. 2008, 56, 1209–1214. [Google Scholar]
- Mangialajo, L.; Ganzin, N.; Accoroni, S.; Asnaghi, V.; Blanfuné, A.; Cabrini, M.; Cattaneo-Vietti, R.; Chavanon, F.; Chiantore, M.; Cohu, S.; et al. Trends in Ostreopsis proliferation along the Northern Mediterranean coasts. Toxicon 2011, 57, 408–420. [Google Scholar] [CrossRef] [Green Version]
- Cohu, S.; Thibaut, T.; Mangialajo, L.; Labat, J.P.; Passafiume, O.; Blanfuné, A.; Simon, N.; Cottalorda, J.M.; Lemée, R. Occurence of the toxic dinoflagellate Ostreopsis cf. ovata in relation with environmental factors in Monaco (NW Mediterrean). Mar. Pollut. Bull. 2011, 62, 2681–2691. [Google Scholar]
- Brescianini, C.; Grillo, C.; Melchiorre, N.; Bertolotto, R.; Ferrari, A.; Vivaldi, B.; Icardi, G.; Gramaccioni, L.; Funari, E.; Scardala, S. Ostreopsis ovata algal blooms affecting human health in Genova, Italy, 2005 and 2006. Eurosurveillance 2006, 11, article 3. [Google Scholar]
- Hoffmann, K.; Hermanns-Clausen, M.; Buhl, C.; Büchler, M.; Schemmer, P.; Mebs, D.; Kauferstein, S. A case of palytoxin poisonning due to contact with zoanthid corals through a skin injury. Toxicon 2008, 51, 1535–1537. [Google Scholar]
- Tichadou, L.; Glaizal, M.; Armengaud, A.; Grossel, H.; Lemée, R.; Kantin, R.; Lasalle, J.L.; Drouet, G.; Rambaud, L.; Malfait, P.; et al. Health impact of unicellular algae of the Ostreopsis genus blooms in the Mediterranean Sea: Experience of the French Mediterranean Coast Surveillance Network from 2006 to 2009. Clin. Toxicol. 2010, 48, 839–844. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku District. Bull. Jpn. Soc. Sci. Fish. 1978, 44, 1249–1255. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Rossi, R.; Soprano, V.; Capozzo, D.; Serpe, L. Palytoxin in seafood by liquid chromatography tandem mass spectrometry: Investigation of extraction efficiency and matrix effect. Anal. Bioanal. Chem. 2011, 401, 1043–1050. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative palytoxin and its new analogue, ovatoxin-a, in Ostreopsis ovata collected along the ligurian coasts during the 2006 toxic outbreak. Rapid Commun. Mass Spectrom. 2008, 19, 111–120. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Guerrini, F.; Pistocchi, R. Complex palytoxin-like profile of Ostreopsis ovata. Identification of four new ovatoxins by high-resolution liquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 2735–2744. [Google Scholar]
- Congrestri, R.; Penna, A.; Zingone, A. BENTOX-NET, a research and management initiative on Ostreopsis spp. and other benthic microalgal blooms on the Italian coast. Harmful Algae News 2007, No. 32, 11–12. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, G.S.; Tartaglione, L.; Grillo, C.; Melchiorre, N. The Genoa 2005 outbreak. Determination of putative palytoxin in mediterranean Ostreopsis ovata by a new liquid chromatography tandem mass spectrometry method. Anal. Chem. 2006, 78, 6153–6159. [Google Scholar]
- Illoul, H.; Hernandez, F.R.; Vila, M.; Adjas, N.; Younes, A.A.; Bournissa, M. The Genus Ostreopsis along the Algiers Coastal Waters (SW Mediterranean Sea) Associated with a Human Respiratory Intoxication Episode. The International Conference on Ostreopsis Development (ICOD), Villefranche-sur-Mer, France, 4–8 April 2011.
- Gallitelli, M.; Ungaro, N.; Addante, L.M.; Procacci, V.; Silver, N.G.; Sabbà, C. Respiratory illness as a reaction to tropical algal blooms occurring in a temperate climate. JAMA 2005, 293, 2599–2600. [Google Scholar]
- Pistocchi, R.; Pezzolesi, L.; Guerrini, F.; Vanucci, S.; Dell’Aversano, C.; Fattorusso, E. A review on the effects of environmental conditions on growth and toxin production of Ostreopsis ovat. Toxicon 2011, 57, 421–428. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific opinion on marine biotoxins in shellfish—Palytoxin group. EFSA J. 2009, 7, 1393:1–1393:38.
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrates Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Edgar, R.C. Muscle: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinforma. 2004, 5, 113. [Google Scholar]
- BioEdit, version 7.1.3, Ibis Biosciences: Carlsbad, CA, USA, 2011; biological sequences alignment editor for windows.
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Posada, D. jModeltest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar]
- MrBayes, version 3.1.2 ed. Available online: http://mrbayes.sourceforge.net (accessed on 23 December 2005). Bayesian Inference of Phylogeny.
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Amzil, Z.; Sibat, M.; Chomerat, N.; Grossel, H.; Marco-Miralles, F.; Lemee, R.; Nezan, E.; Sechet, V. Ovatoxin-a and Palytoxin Accumulation in Seafood in Relation to Ostreopsis cf. ovata Blooms on the French Mediterranean Coast. Mar. Drugs 2012, 10, 477-496. https://doi.org/10.3390/md10020477
Amzil Z, Sibat M, Chomerat N, Grossel H, Marco-Miralles F, Lemee R, Nezan E, Sechet V. Ovatoxin-a and Palytoxin Accumulation in Seafood in Relation to Ostreopsis cf. ovata Blooms on the French Mediterranean Coast. Marine Drugs. 2012; 10(2):477-496. https://doi.org/10.3390/md10020477
Chicago/Turabian StyleAmzil, Zouher, Manoella Sibat, Nicolas Chomerat, Hubert Grossel, Francoise Marco-Miralles, Rodolphe Lemee, Elisabeth Nezan, and Veronique Sechet. 2012. "Ovatoxin-a and Palytoxin Accumulation in Seafood in Relation to Ostreopsis cf. ovata Blooms on the French Mediterranean Coast" Marine Drugs 10, no. 2: 477-496. https://doi.org/10.3390/md10020477
APA StyleAmzil, Z., Sibat, M., Chomerat, N., Grossel, H., Marco-Miralles, F., Lemee, R., Nezan, E., & Sechet, V. (2012). Ovatoxin-a and Palytoxin Accumulation in Seafood in Relation to Ostreopsis cf. ovata Blooms on the French Mediterranean Coast. Marine Drugs, 10(2), 477-496. https://doi.org/10.3390/md10020477