Inhibition of Hepatitis C Virus Replication and Viral Helicase by Ethyl Acetate Extract of the Marine Feather Star Alloeocomatella polycladia

 , add
Show full author list
, add
Show full author list
Abstract
:1. Introduction
2. Results and Discussion
2.1. Primary Screening of Marine Organism Extracts on HCV NS3 Helicase Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Helicase Activity | ||||||
|---|---|---|---|---|---|---|
| Sample | (% of control) | Specimen | Phylum | Extract | Collection Site | |
| OK-99-2 | 78 | Agelas sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-3 | 73 | Plakortis sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-4 | 60 | Dysidea arenaria | Porifera | EtOAc | Shimoji Island | |
| OK-99-5 | 96 | Theonella cupola | Porifera | EtOAc | Shimoji Island | |
| OK-99-6 | 52 | Theonella conica | Porifera | EtOAc | Shimoji Island | |
| OK-99-7 | 85 | Epipolasis kushimotoensis | Porifera | EtOAc | Shimoji Island | |
| OK-99-9 | 51 | Hyrtios sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-10 | 75 | Theonella sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-12 | 53 | Isis hippuris | Cnidaria | EtOAc | Shimoji Island | |
| OK-99-13 | 68 | Acanthella sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-15 | 64 | Phyllospongia sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-17 | 59 | Petrosia sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-18 | 80 | Fasciospongia rimosa | Porifera | EtOAc | Shimoji Island | |
| OK-99-20 | 77 | Echinoclathria sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-21 | 68 | Strongylophora sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-23 | 74 | Dysidea herbacea | Porifera | EtOAc | Shimoji Island | |
| OK-99-26 | 55 | Dysidea cf. arenaria | Porifera | EtOAc | Shimoji Island | |
| OK-99-28 | 123 | Plakortis sp. | Porifera | EtOAc | Shimoji Island | |
| OK-99-31 | 118 | Spongia sp. | Porifera | EtOAc | Okinawa Island | |
| OK-99-34 | 119 | Theonella swinhoei | Porifera | EtOAc | Okinawa Island | |
| OK-99-35 | 108 | Petrosia sp. | Porifera | EtOAc | Okinawa Island | |
| OK-99-36 | 90 | Acanthella sp. | Porifera | EtOAc | Okinawa Island | |
| OK-99-37 | 102 | Luffariella sp. | Porifera | EtOAc | Okinawa Island | |
| OK-99-41 | 62 | Dysidea cf. arenaria | Porifera | EtOAc | Okinawa Island | |
| OK-99-43 | 85 | Xestospongia sp. | Porifera | EtOAc | Okinawa Island | |
| OK-99-44 | 61 | Dysidea arenaria | Porifera | EtOAc | Okinawa Island | |
| OK-99-47 | 108 | Dysidea cf. arenaria | Porifera | EtOAc | Okinawa Island | |
| OK-99-49 | 90 | Petrosia sp. | Porifera | EtOAc | Chibishi | |
| OK-99-51 | 69 | Isis hippuris | Cnidaria | EtOAc | Chibishi | |
| OK-99-52 | 78 | Petrosia sp. | Porifera | EtOAc | Kuro Island | |
| OK-99-55 | 65 | Acanthella sp. | Porifera | EtOAc | Kuro Island | |
| OK-99-57 | 84 | Theonella swinhoei | Porifera | EtOAc | Kuro Island | |
| OK-99-63 | 117 | Epipolasis kushimotoensis | Porifera | EtOAc | Kuro Island | |
| OK-99-64 | 98 | Xestospongia sp. | Porifera | EtOAc | Kuro Island | |
| SG1-1-2 | 77 | Comanthus gisleni | Echinodermata | MeOH | Kume Island | |
| SG1-2-2 | 112 | Stephanometra indica | Echinodermata | MeOH | Kume Island | |
| SG1-5-2 | 33 | Comantella sp. cf. maculata | Echinodermata | MeOH | Kume Island | |
| SG1-9-2 | 57 | Phanogenia gracilis | Echinodermata | MeOH | Kume Island | |
| SG1-12-2 | 39 | Comanthus parvicirrus | Echinodermata | MeOH | Kume Island | |
| SG1-14-2 | 117 | Comaster schlegelii | Echinodermata | MeOH | Kume Island | |
| SG1-15-2 | 26 | Colobometridae sp. | Echinodermata | MeOH | Kume Island | |
| SG1-16-2 | 66 | Cenometra bella | Echinodermata | MeOH | Kume Island | |
| SG1-19-2 | 78 | Comaster nobilis | Echinodermata | MeOH | Kume Island | |
| SG1-21-2 | 32 | Oxycomanthus sp. | Echinodermata | MeOH | Kume Island | |
| SG1-23-1 | -3 | Alloeocomatella polycladia | Echinodermata | EtOAc | Kume Island | |
| SG1-24-1 | 24 | Comanthus sp. | Echinodermata | EtOAc | Kume Island | |
| SG1-26-2 | 51 | Oxycomanthus benetti | Echinodermata | MeOH | Kume Island | |
| SG1-28-2 | 38 | Lamprometra palmata | Echinodermata | MeOH | Kume Island | |
| SG1-30-1 | 25 | Colobometra perspinosa | Echinodermata | EtOAc | Kume Island | |
| SG1-31-1 | 26 | Comanthus sp. | Echinodermata | EtOAc | Kume Island | |
| SG1-33-1 | 32 | Basilometra boschmai | Echinodermata | EtOAc | Kume Island | |
| SG3-1 | 82 | Stereonephthya sp. | Cnidaria | EtOAc | Tokashiki Island | |
| SG3-4 | 73 | Dysidea cf. arenaria | Porifera | EtOAc | Tokashiki Island | |
| SG3-6 | 74 | Stylotella sp. | Porifera | EtOAc | Tokashiki Island | |
| SG3-10 | 139 | Epipolasis sp. | Porifera | EtOAc | Tokashiki Island | |
| SG3-11 | 97 | Nephthea sp. | Cnidaria | EtOAc | Tokashiki Island | |
| SG3-21 | 106 | Myrmekioderma sp. | Porifera | EtOAc | Tokashiki Island | |
| SG3-25 | 111 | Pseudoceratina purpurea | Porifera | EtOAc | Tokashiki Island | |
| SG3-26 | 95 | Leucetta sp. | Porifera | EtOAc | Tokashiki Island | |
| SG3-28 | 65 | Lyngbya sp. | Cyanobacteria | EtOAc | Tokashiki Island | |
| SG3-29 | 61 | Dysidea sp. | Porifera | EtOAc | Tokashiki Island |



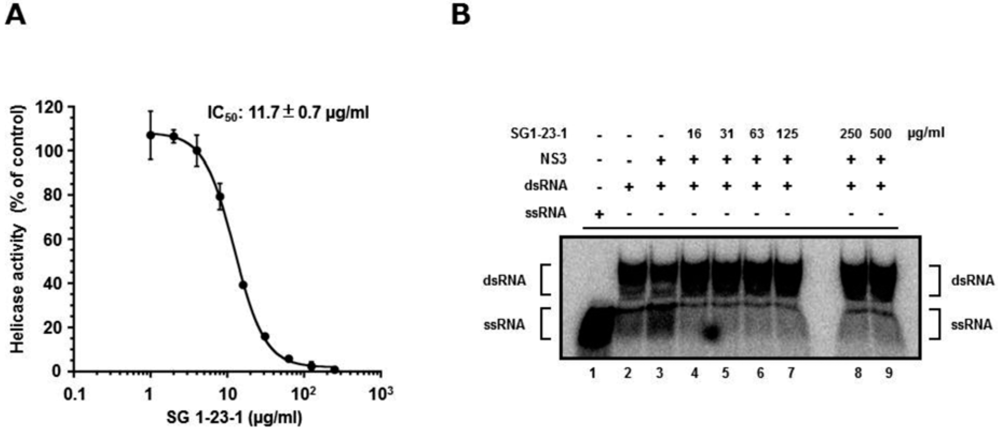
2.2. Effect of SG1-23-1 on HCV NS3 ATPase and RNA Binding Activities

2.3. Effect of SG1-23-1 on HCV RNA Replication in HCV 1b Replicon Cells

| Replicon Cell Line | Virus Strain | EC50 a | EC90 b | CC50 c | |
|---|---|---|---|---|---|
| (Genotype 1b) | (µg/mL) | (µg/mL) | (µg/mL) | ||
| Subgenome | |||||
| Huh7 Lunet/Con1 LUN Sb #26 | Con1 | 22.9 ± 0.4 | 48.1 ± 1.5 | >50 | |
| Huh7 rep Feo | N | 44.2 ± 1.5 | >50 | >50 | |
| Hu7#94/ORN3-5B#24 | O | 19.9 ± 1.8 | 48.8 ± 0.3 | >50 | |
| Full genome | |||||
| OR6 | O | 39.5 ± 0.8 | >50 | >50 | |
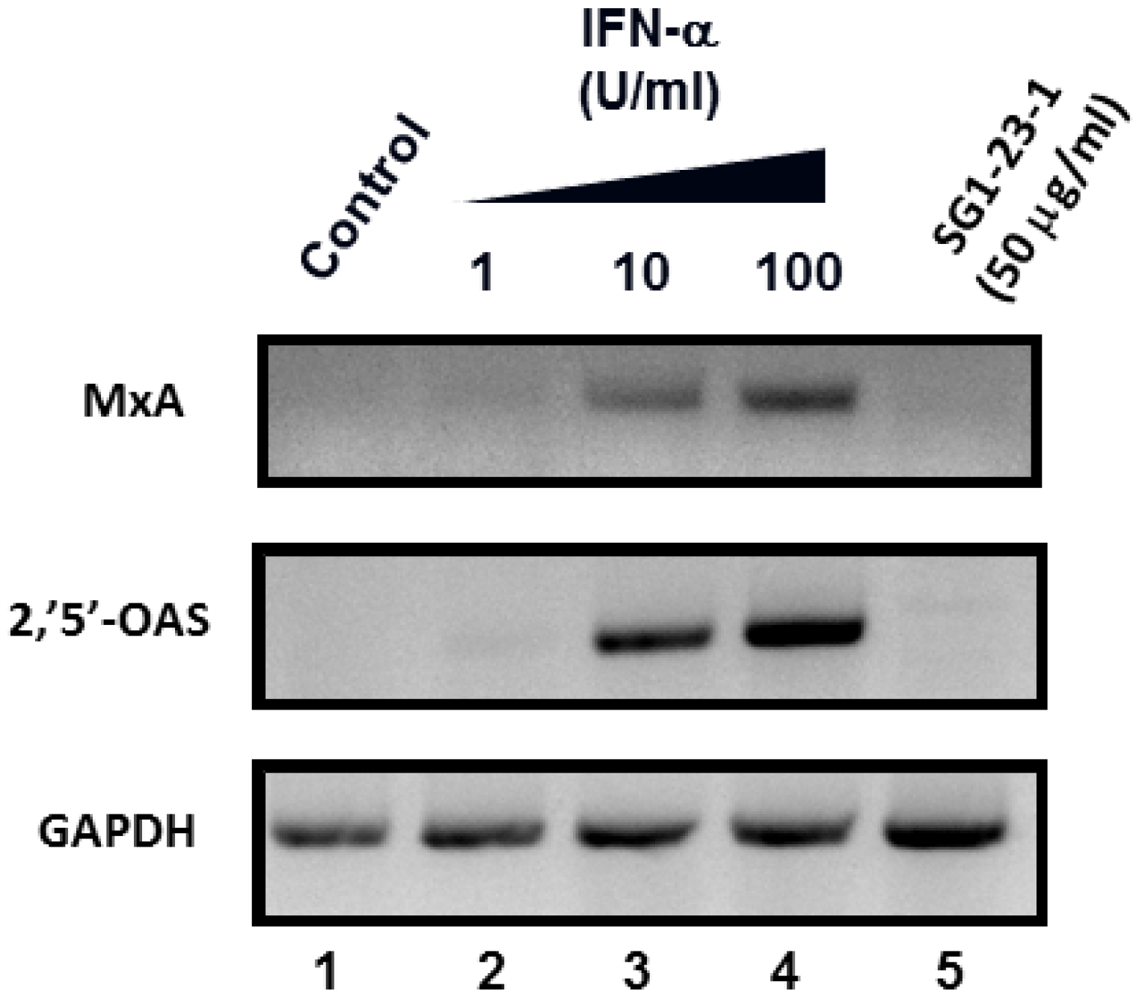
2.4. Effect of SG1-23-1 on the Interferon (IFN) Signaling Pathway

3. Experimental Section
3.1. Preparation of Extracts from Marine Organisms
3.2. High-Throughput Screening of NS3 Helicase Inhibitors
3.3. ATPase Assay
3.4. RNA Helicase Assay
3.5. RNA Binding Assay
3.6. Cell Lines
3.7. Determination of Luciferase Activity in HCV Replicon Cells
3.8. Determination of Cytotoxicity in HCV Replicon Cells
3.9. Effects on Activities of Luciferase and Internal Ribosome Entry Site (IRES)
3.10. Western Blotting
3.11. Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
4. Conclusions
Acknowledgments
References
- Baldo, V.; Baldovin, T.; Trivello, R.; Floreani, A. Epidemiology of HCV infection. Curr. Pharm. Des. 2008, 14, 1646–1654. [Google Scholar]
- Seeff, L.B. Natural history of chronic hepatitis C. Hepatology 2002, 36, S35–S46. [Google Scholar]
- Moriishi, K.; Matsuura, Y. Host factors involved in the replication of hepatitis C virus. Rev. Med. Virol. 2007, 17, 343–354. [Google Scholar]
- Tsukiyama-Kohara, K.; Iizuka, N.; Kohara, M.; Nomoto, A. Internal ribosome entry site within hepatitis C virus RNA. J. Virol. 1992, 66, 1476–1483. [Google Scholar]
- Bartenschlager, R.; Ahlborn-Laake, L.; Mous, J.; Jacobsen, H. Nonstructural protein 3 of the hepatitis C virus encodes a serine-type proteinase required for cleavage at the NS3/4 and NS4/5 junctions. J. Virol. 1993, 67, 3835–3844. [Google Scholar]
- Kim, D.W.; Gwack, Y.; Han, J.H.; Choe, J. C-terminal domain of the hepatitis C virus NS3 protein contains an RNA helicase activity. Biochem. Biophys. Res. Commun. 1995, 215, 160–166. [Google Scholar] [CrossRef]
- Failla, C.; Tomei, L.; de Francesco, R. Both NS3 and NS4A are required for proteolytic processing of hepatitis C virus nonstructural proteins. J. Virol. 1994, 68, 3753–3760. [Google Scholar]
- Belon, C.A.; Frick, D.N. Helicase inhibitors as specifically targeted antiviral therapy for hepatitis C. Future Virol. 2009, 4, 277–293. [Google Scholar]
- Frick, D.N. The hepatitis C virus NS3 protein: a model RNA helicase and potential drug target. Curr. Issues Mol. Biol. 2007, 9, 1–20. [Google Scholar]
- Kwong, A.D.; Rao, B.G.; Jeang, K.T. Viral and cellular RNA helicases as antiviral targets. Nat. Rev. Drug Discov. 2005, 4, 845–853. [Google Scholar]
- Manns, M.P.; Wedemeyer, H.; Cornberg, M. Treating viral hepatitis C: efficacy, side effects, and complications. Gut 2006, 55, 1350–1359. [Google Scholar]
- McHutchison, J.G.; Everson, G.T.; Gordon, S.C.; Jacobson, I.M.; Sulkowski, M.; Kauffman, R.; McNair, L.; Alam, J.; Muir, A.J.; Team, P.S. Telaprevir with peginterferon and ribavirin for chronic HCV genotype 1 infection. N. Engl. J. Med. 2009, 360, 1827–1838. [Google Scholar]
- Zeuzem, S.; Hultcrantz, R.; Bourliere, M.; Goeser, T.; Marcellin, P.; Sanchez-Tapias, J.; Sarrazin, C.; Harvey, J.; Brass, C.; Albrecht, J. Peginterferon alfa-2b plus ribavirin for treatment of chronic hepatitis C in previously untreated patients infected with HCV genotypes 2 or 3. J. Hepatol. 2004, 40, 993–999. [Google Scholar]
- Asselah, T.; Marcellin, P. New direct-acting antivirals’ combination for the treatment of chronic hepatitis C. Liver Int. 2011, 31 Suppl 1, 68–77. [Google Scholar] [CrossRef]
- Jazwinski, A.B.; Muir, A.J. Direct-acting antiviral medications for chronic hepatitis C virus infection. Gastroenterol. Hepatol. (NY) 2011, 7, 154–162. [Google Scholar]
- Lange, C.M.; Sarrazin, C.; Zeuzem, S. Review article: specifically targeted anti-viral therapy for hepatitis C—A new era in therapy. Aliment. Pharmacol. Ther. 2010, 32, 14–28. [Google Scholar]
- Hofmann, W.P.; Zeuzem, S. A new standard of care for the treatment of chronic HCV infection. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 257–264. [Google Scholar]
- Kwong, A.D.; Kauffman, R.S.; Hurter, P.; Mueller, P. Discovery and development of telaprevir: An NS3-4A protease inhibitor for treating genotype 1 chronic hepatitis C virus. Nat. Biotechnol. 2011, 29, 993–1003. [Google Scholar]
- Kieffer, T.L.; Kwong, A.D.; Picchio, G.R. Viral resistance to specifically targeted antiviral therapies for hepatitis C (STAT-Cs). J. Antimicrob. Chemother. 2010, 65, 202–212. [Google Scholar]
- Thompson, A.J.; McHutchison, J.G. Antiviral resistance and specifically targeted therapy for HCV (STAT-C). J. Viral. Hepat. 2009, 16, 377–387. [Google Scholar]
- Belon, C.A.; High, Y.D.; Lin, T.I.; Pauwels, F.; Frick, D.N. Mechanism and specificity of a symmetrical benzimidazolephenylcarboxamide helicase inhibitor. Biochemistry 2010, 49, 1822–1832. [Google Scholar]
- Maga, G.; Gemma, S.; Fattorusso, C.; Locatelli, G.A.; Butini, S.; Persico, M.; Kukreja, G.; Romano, M.P.; Chiasserini, L.; Savini, L.; et al. Specific targeting of hepatitis C virus NS3 RNA helicase. Discovery of the potent and selective competitive nucleotide-mimicking inhibitor QU663. Biochemistry 2005, 44, 9637–9644. [Google Scholar]
- Chin, Y.W.; Balunas, M.J.; Chai, H.B.; Kinghorn, A.D. Drug discovery from natural sources. AAPS J. 2006, 8, E239–E253. [Google Scholar]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar]
- Li, J.W.; Vederas, J.C. Drug discovery and natural products: End of an era or an endless frontier? Science 2009, 325, 161–165. [Google Scholar] [CrossRef]
- Ahmed-Belkacem, A.; Ahnou, N.; Barbotte, L.; Wychowski, C.; Pallier, C.; Brillet, R.; Pohl, R.T.; Pawlotsky, J.M. Silibinin and related compounds are direct inhibitors of hepatitis C virus RNA-dependent RNA polymerase. Gastroenterology 2010, 138, 1112–1122. [Google Scholar]
- Wagoner, J.; Negash, A.; Kane, O.J.; Martinez, L.E.; Nahmias, Y.; Bourne, N.; Owen, D.M.; Grove, J.; Brimacombe, C.; McKeating, J.A.; et al. Multiple effects of silymarin on the hepatitis C virus lifecycle. Hepatology 2010, 51, 1912–1921. [Google Scholar] [CrossRef]
- Ciesek, S.; von Hahn, T.; Colpitts, C.C.; Schang, L.M.; Friesland, M.; Steinmann, J.; Manns, M.P.; Ott, M.; Wedemeyer, H.; Meuleman, P.; et al. The green tea polyphenol, epigallocatechin-3-gallate, inhibits hepatitis C virus entry. Hepatology 2011, 54, 1947–1955. [Google Scholar] [CrossRef]
- Takeshita, M.; Ishida, Y.; Akamatsu, E.; Ohmori, Y.; Sudoh, M.; Uto, H.; Tsubouchi, H.; Kataoka, H. Proanthocyanidin from blueberry leaves suppresses expression of subgenomic hepatitis C virus RNA. J. Biol. Chem. 2009, 284, 21165–21176. [Google Scholar]
- Donia, M.; Hamann, M.T. Marine natural products and their potential applications as anti-infective agents. Lancet Infect. Dis. 2003, 3, 338–348. [Google Scholar]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar]
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar]
- Yokota, T.; Sakamoto, N.; Enomoto, N.; Tanabe, Y.; Miyagishi, M.; Maekawa, S.; Yi, L.; Kurosaki, M.; Taira, K.; Watanabe, M.; et al. Inhibition of intracellular hepatitis C virus replication by synthetic and vector-derived small interfering RNAs. EMBO Rep. 2003, 4, 602–608. [Google Scholar] [CrossRef]
- Frese, M.; Barth, K.; Kaul, A.; Lohmann, V.; Schwarzle, V.; Bartenschlager, R. Hepatitis C virus RNA replication is resistant to tumour necrosis factor-alpha. J. Virol. 2003, 84, 1253–1259. [Google Scholar]
- Ikeda, M.; Abe, K.; Dansako, H.; Nakamura, T.; Naka, K.; Kato, N. Efficient replication of a full-length hepatitis C virus genome, strain O, in cell culture, and development of a luciferase reporter system. Biochem. Biophys. Res. Commun. 2005, 329, 1350–1359. [Google Scholar] [CrossRef]
- Blight, K.J.; Kolykhalov, A.A.; Rice, C.M. Efficient initiation of HCV RNA replication in cell culture. Science 2000, 290, 1972–1974. [Google Scholar]
- Guo, J.T.; Bichko, V.V.; Seeger, C. Effect of alpha interferon on the hepatitis C virus replicon. J. Virol. 2001, 75, 8516–8523. [Google Scholar]
- Tani, H.; Akimitsu, N.; Fujita, O.; Matsuda, Y.; Miyata, R.; Tsuneda, S.; Igarashi, M.; Sekiguchi, Y.; Noda, N. High-throughput screening assay of hepatitis C virus helicase inhibitors using fluorescence-quenching phenomenon. Biochem. Biophys. Res. Commun. 2009, 379, 1054–1059. [Google Scholar]
- Gallinari, P.; Brennan, D.; Nardi, C.; Brunetti, M.; Tomei, L.; Steinkuhler, C.; De Francesco, R. Multiple enzymatic activities associated with recombinant NS3 protein of hepatitis C virus. J. Virol. 1998, 72, 6758–6769. [Google Scholar]
- Huang, Y.; Liu, Z.R. The ATPase, RNA unwinding, and RNA binding activities of recombinant p68 RNA helicas. J. Biol. Chem. 2002, 277, 12810–12815. [Google Scholar]
- Jin, H.; Yamashita, A.; Maekawa, S.; Yang, P.; He, L.; Takayanagi, S.; Wakita, T.; Sakamoto, N.; Enomoto, N.; Ito, M. Griseofulvin, an oral antifungal agent, suppresses hepatitis C virus replication in vitro. Hepatol. Res. 2008, 38, 909–918. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yamashita, A.; Salam, K.A.; Furuta, A.; Matsuda, Y.; Fujita, O.; Tani, H.; Fujita, Y.; Fujimoto, Y.; Ikeda, M.; Kato, N.; et al. Inhibition of Hepatitis C Virus Replication and Viral Helicase by Ethyl Acetate Extract of the Marine Feather Star Alloeocomatella polycladia. Mar. Drugs 2012, 10, 744-761. https://doi.org/10.3390/md10040744
Yamashita A, Salam KA, Furuta A, Matsuda Y, Fujita O, Tani H, Fujita Y, Fujimoto Y, Ikeda M, Kato N, et al. Inhibition of Hepatitis C Virus Replication and Viral Helicase by Ethyl Acetate Extract of the Marine Feather Star Alloeocomatella polycladia. Marine Drugs. 2012; 10(4):744-761. https://doi.org/10.3390/md10040744
Chicago/Turabian StyleYamashita, Atsuya, Kazi Abdus Salam, Atsushi Furuta, Yasuyoshi Matsuda, Osamu Fujita, Hidenori Tani, Yoshihisa Fujita, Yuusuke Fujimoto, Masanori Ikeda, Nobuyuki Kato, and et al. 2012. "Inhibition of Hepatitis C Virus Replication and Viral Helicase by Ethyl Acetate Extract of the Marine Feather Star Alloeocomatella polycladia" Marine Drugs 10, no. 4: 744-761. https://doi.org/10.3390/md10040744
APA StyleYamashita, A., Salam, K. A., Furuta, A., Matsuda, Y., Fujita, O., Tani, H., Fujita, Y., Fujimoto, Y., Ikeda, M., Kato, N., Sakamoto, N., Maekawa, S., Enomoto, N., Nakakoshi, M., Tsubuki, M., Sekiguchi, Y., Tsuneda, S., Akimitsu, N., Noda, N., ... Moriishi, K. (2012). Inhibition of Hepatitis C Virus Replication and Viral Helicase by Ethyl Acetate Extract of the Marine Feather Star Alloeocomatella polycladia. Marine Drugs, 10(4), 744-761. https://doi.org/10.3390/md10040744