A Shark Liver Gene-Derived Active Peptide Expressed in the Silkworm, Bombyx mori: Preliminary Studies for Oral Administration of the Recombinant Protein

Abstract
:
1. Introduction
2. Results
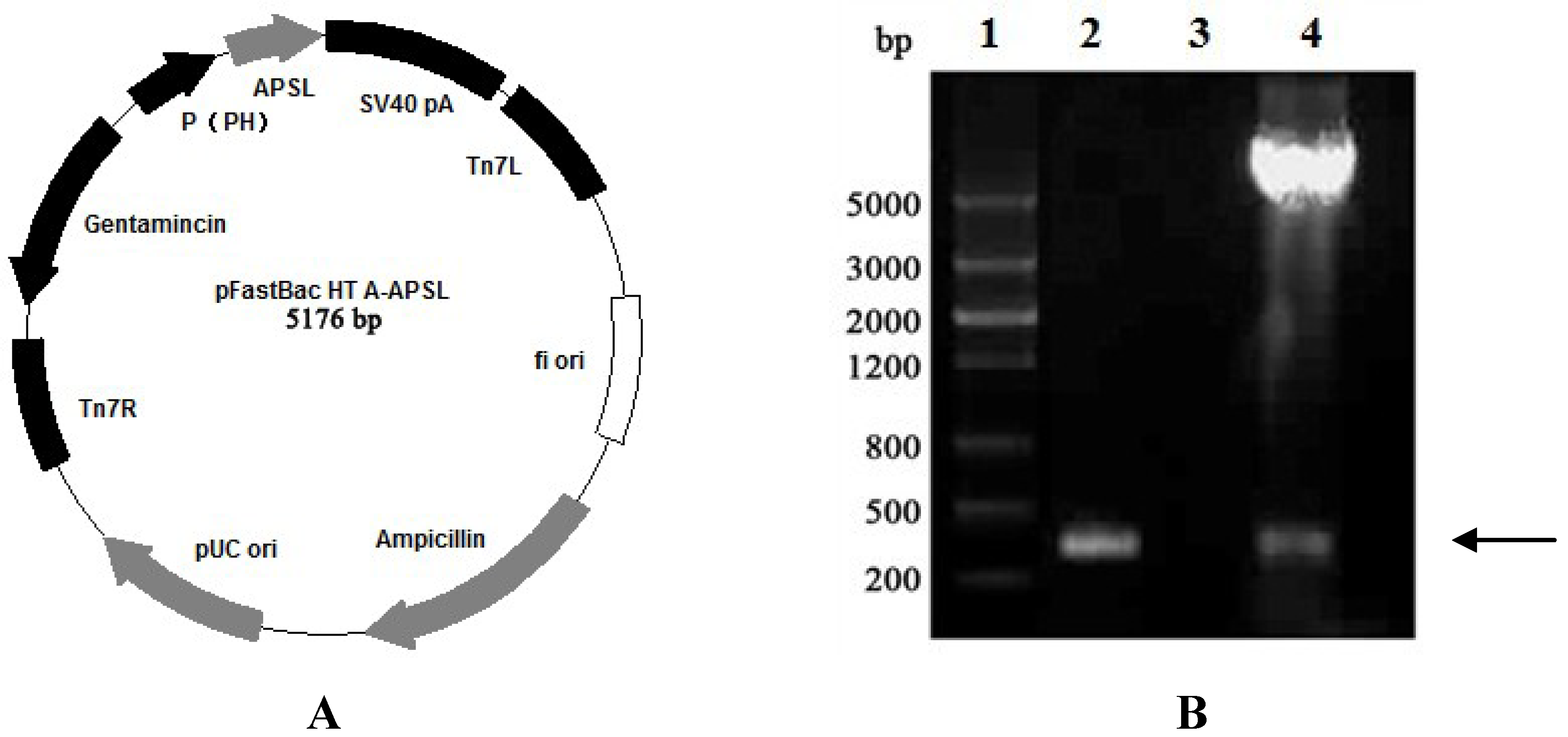
2.1. Construction of Recombinant Plasmid pFastBac HTA-APSL

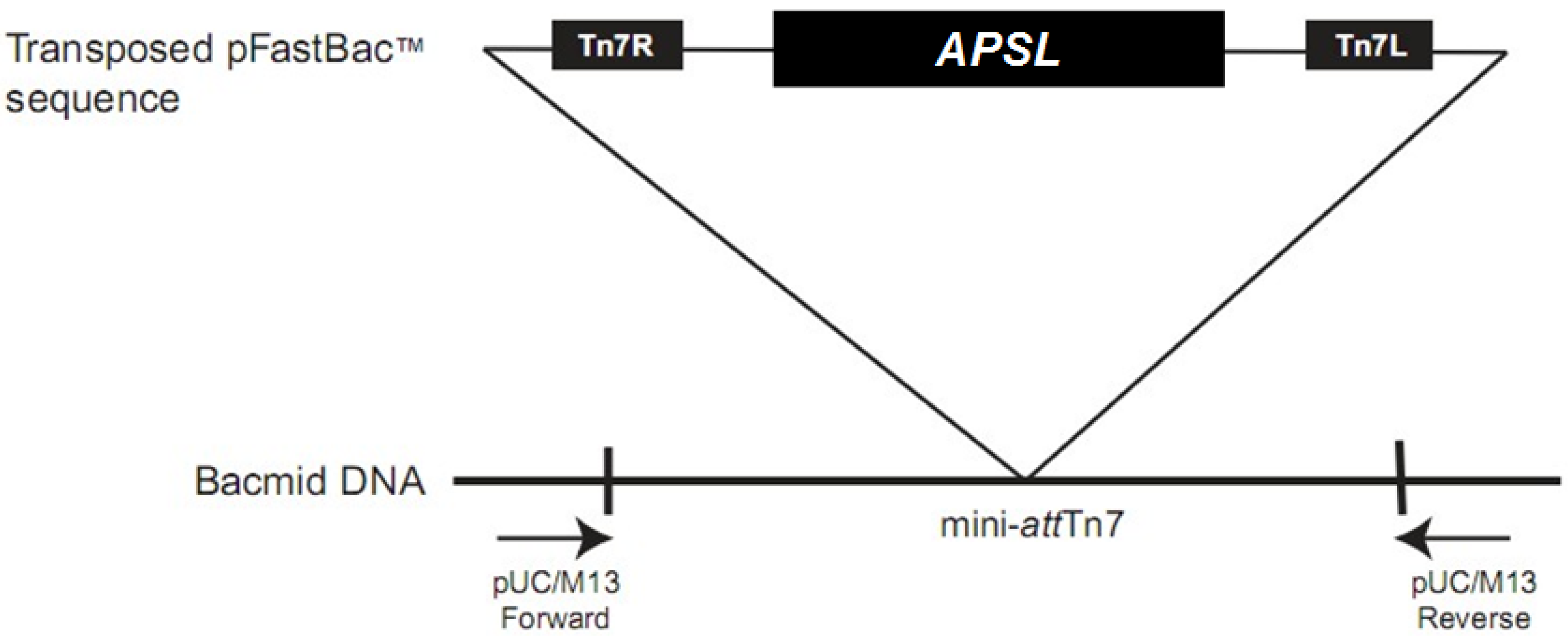
2.2. Construction and Isolation of Recombinant Bacmid


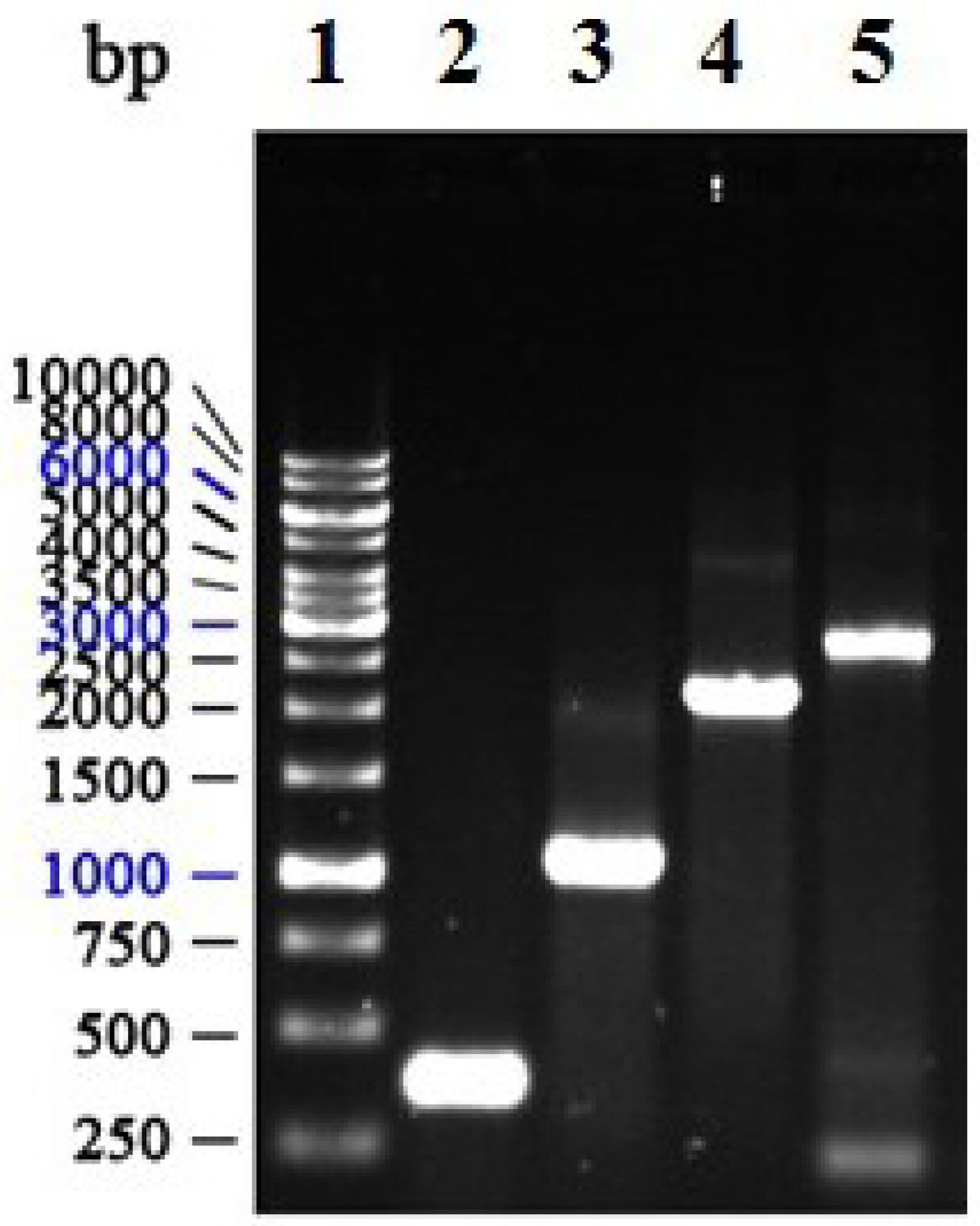
2.3. Construction, Isolation and Analysis of Recombinant Bacmid


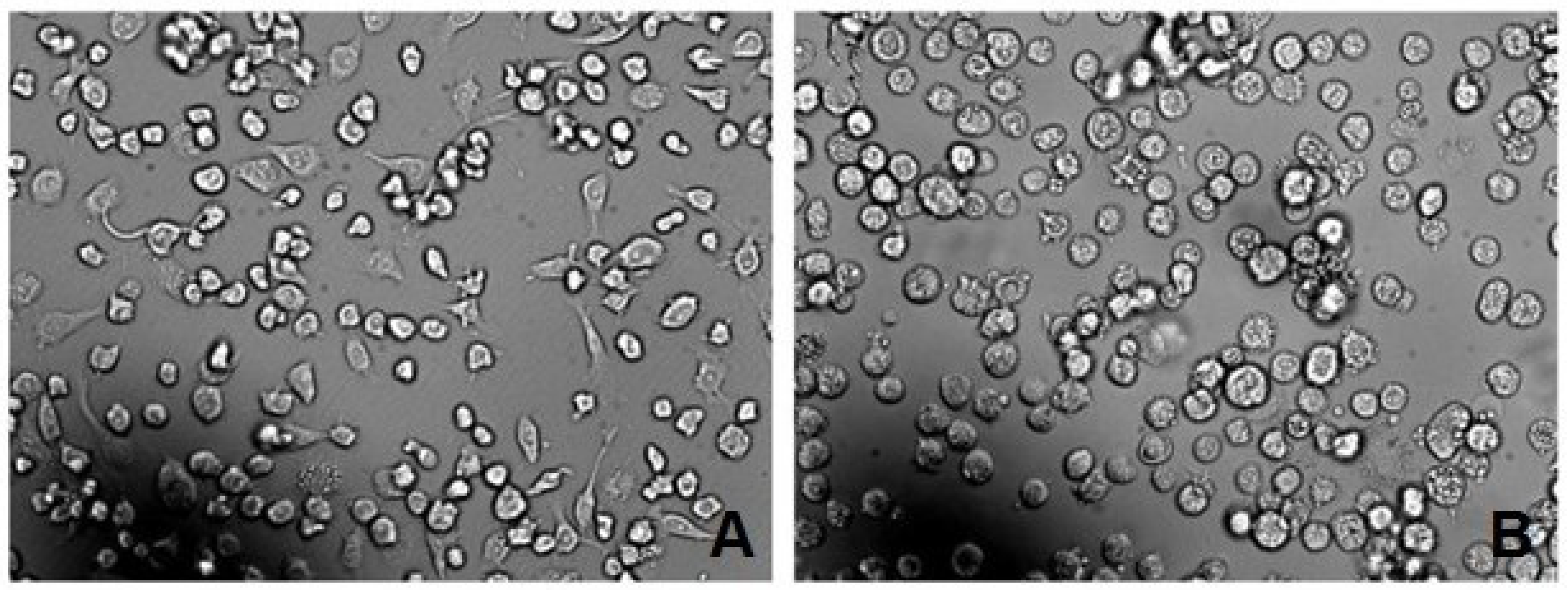
2.4. Expression, Collection and rAPSL Assay of BmN Cells

2.5. Expression, Collection and rAPSL Assay of Silkworm Larvae and Pupae

2.6. Bioactivity Analysis of Silkworm Pupae-Expressed rAPSL

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | FBG (mmol/L) | ||||
|---|---|---|---|---|---|
| First week | Second week | Third week | Forth week | Fifth week | |
| Control | 4.72 ± 0.92 | 4.81 ± 0.86 | 4.83 ± 1.07 | 4.83 ± 1.38 | 4.78 ± 1.16 |
| Model | 28.79 ± 3.44 | 27.28 ± 2.39 | 26.32 ± 3.07 | 25.6 ± 1.41 ▲▲▲ | All Died |
| Pupae | 26.48 ± 3.38 | 25.20 ± 2.43 | 23.39 ± 2.51 | 21.75 ± 2.01 * | 20.22 ± 1.61 ** |
| rAPSL | 28.57 ± 2.81 | 25.15 ± 3.23 | 21.33 ± 3.10 ** | 19.09 ± 2.80 ** | 15.00 ± 2.72 *** |

3. Discussion
4. Experimental
4.1. Materials
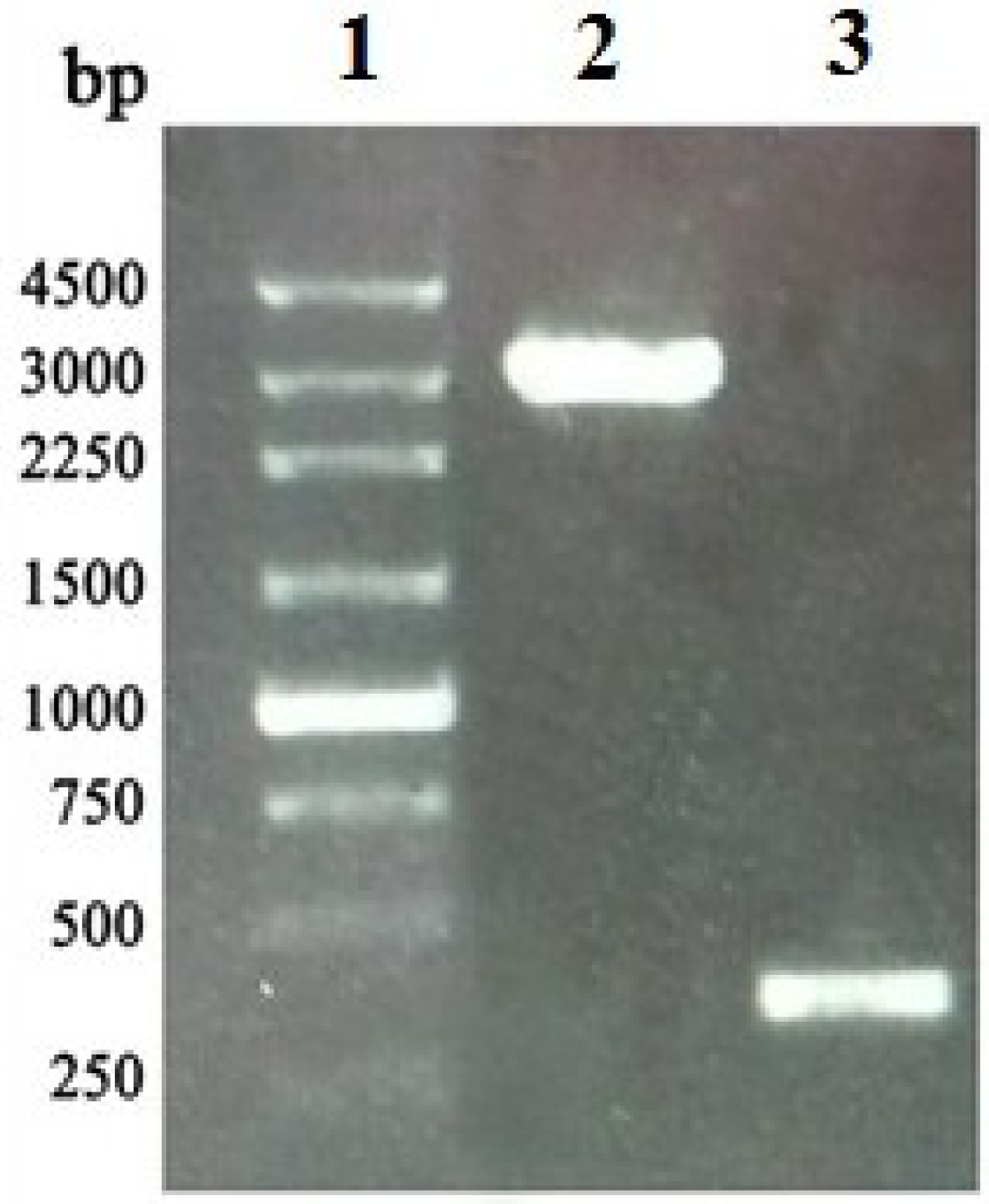
4.2. Amplification, Cloning and Identification of the APSL Gene
4.3. Construction of the Recombinant Plasmid pFastBac HTA-APSL
4.4. Construction, Isolation and Analysis of the Recombinant Bacmid
4.5. Expression of the APSL Gene in BmN Cells
4.6. Expression of the APSL Gene in Silkworm Larvae and Pupae
4.7. Protective Effects against Streptozotocin-Induced Diabetes Mellitus in Mice
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Harris, M.; Zimmet, P. Classification of diabetes mellitus and other categories of glucose intolerance. In International Textbook of Diabetes Mellitus, 2nd ed; John Wiley and Sons: Chichester, UK, 1997; pp. 9–23. [Google Scholar]
- International Diabetes Federation, IDF Diabetes Atlas, 5th ed; International Diabetes Federation: Brussels, Belgium, 2011.
- Zhang, Y.Z.; Chen, Z.; Lv, Z.B.; Nie, Z.M.; Zhang, X.Y.; Wu, X.F. Can 29 kDa rhGM-CSF expressed by silkworm pupae bioreactor bring into effect as active cytokine through orally administration? Eur. J. Pharm. Sci. 2006, 28, 212–223. [Google Scholar]
- Ryu, K.S.; Lee, H.S.; Chung, S.H.; Kang, P.D. An activity of lowering blood-glucose levels according to preparative condition of silkworm powder. Korean J. Seric. Sci. 1997, 39, 79–85. [Google Scholar]
- Ryu, K.S.; Lee, H.S.; Kim, S.Y. Pharmacodynamic study of silkworm powder in mice administered to maltose, sucrose and lactose. Korean J. Seric. Sci. 1999, 41, 9–13. [Google Scholar]
- Han, J.; Inoue, S.; Isoda, H. Effects of silkworm powder on glucose absorption by human intestinal epithelial cell line Caco-2. J. Nat. Med. 2007, 61, 387–390. [Google Scholar] [CrossRef] [Green Version]
- Maeda, S.; Kawai, T.; Obinata, M.; et al. Production of human alpha-interferon in silkworm using baculovirus vector. Nature 1985, 315, 592–594. [Google Scholar]
- Miao, Y.G.; Zhang, Y.S.; Nakagaki, K.; Zhao, T.F.; Zhao, A.C.; Meng, Y.; Nakagaki, M.; Park, E.Y.; Maenaka, K. Expression of spider flagelliform silk protein in Bombyx mori cell line by a novel Bac-to-Bac/BmNPV baculovirus expression system. Appl. Microbiol. Biot. 2006, 71, 192–199. [Google Scholar] [CrossRef]
- Carbonell, L.F.; Miller, L.K. Baculovirus interaction with nontarget organisms: A virus-borne reporter gene is not expressed in two mammalian cell lines. Appl. Environ. Microbiol. 1987, 53, 1412–1417. [Google Scholar]
- Herrington, D.A.; Losonsdy, G.A.; Smith, G.; Vovovitz, F.; Cochran, M.; Jackson, K.; Hoffman, S.L.; Gordon, D.M.; Levine, M.M.; Edelman, R. Safety and immunogenicity in volunteers of a recombinant Plasmodium falciparum circumsporozoite protein malaria vaccine produced in Lepidopteran cells. Vaccine 1992, 10, 841–846. [Google Scholar] [CrossRef]
- Yu, W.; Chen, J.; Zhao, X.L.; Lv, Z.B.; Nie, Z.M.; Zhang, Y.Z. Expression of polyhedrin-hEGF fusion protein in cultured cells and larvae of Bombyx mori. Afr. J. Biotechnol. 2006, 11, 1034–1040. [Google Scholar]
- Zhang, W.; Lv, Z.; Nie, Z.; Chen, G.; Chen, J.; Sheng, Q.; Yu, W.; Jin, Y.; Wu, X.; Zhang, Y. Bioavailability of orally administered rhGM-CSF: A single-dose, randomized, open-label, two-period crossover trial. PLoS One 2009, 4, e5353. [Google Scholar]
- Yue, W.F.; Miao, Y.G.; Li, X.H.; Wu, X.F.; Zhao, A.C.; Nakagaki, M. Cloning and expression of manganese superoxide dismutase of the silkworm, Bombyx mori by Bac-to-Bac/BmNPV Baculovirus expression system. Appl. Microbiol. Biotechnol. 2006, 73, 181–186. [Google Scholar] [CrossRef]
- Kato, T.; Kajikawa, M.; Maenaka, K.; Park, E.Y. Silkworm expression system as a platform technology in life science. Appl. Microbiol. Biotechnol. 2010, 85, 459–470. [Google Scholar] [CrossRef]
- Lv, Z.B.; Ou, Y.; Li, Q.; Zhang, W.P.; Ye, B.P.; Wu, W.T. Expression, purification and bioactivities analysis of recombinant active peptide from shark liver. Mar. Drugs 2009, 7, 258–267. [Google Scholar] [CrossRef]
- Huang, F.J.; Wu, W.T. Antidiabetic effect of a new peptide from squalus mitsukurii liver (S-8300) in Alloxan-induced diabetes. Clin. Exp. Pharmacol. Physiol. 2005, 32, 521–525. [Google Scholar] [CrossRef]
- Huang, F.J.; Wu, W.T. Purification and characterization of a new peptide (S-8300) from shark liver. J. Food Biochem. 2010, 34, 962–970. [Google Scholar]
- Huang, F.J.; Wu, W.T. Antidiabetic effect of a new peptide from squalus mitsukurii liver (S-8300) in Streptozotocin-induced diabetes. J. Pharm. Pharmacol. 2005, 57, 1575–1580. [Google Scholar]
- Ou, Y.; Li, Q.; Lv, Z.B.; Wu, W.T.; Wang, Q.J. Purification and characterization of hepatocyte regeneration stimulatory factor from shark liver. J. Chin. Pharm. Sci. 2003, 12, 175–180. [Google Scholar]
- LaBrecque, D.R.; Steele, G.; Fogerty, S.; Wilson, M.; Barton, J. Purification and physical-chemical characterization of hepatic stimulator substance. Hepatology 1987, 7, 100–106. [Google Scholar] [CrossRef]
- Luckow, V.A.; Lee, S.C.; Barry, G.F.; Olins, P.O. Efficient generation of infectious recombinant baculovirues by site-specific transposon-mediated insertion of foreign genes into a baculovirus genome propagated in Escherichia coli. J. Virol. 1993, 67, 4566–4579. [Google Scholar]
- Smith, G.E.; Summers, M.D.; Fraser, M.J. Production of human beta interferon in insect cells infected with a baculovirus expression vector. Mol. Cell. Biol. 1983, 3, 2156–2165. [Google Scholar]
- Jönsson, B.; CODE-2 Advisory Board. Revealing the cost of Type II in Europe. Diabetologia 2002, 45, S5–S12. [Google Scholar] [CrossRef]
- American Diabetes Association. Direct and Indirect Costs of Diabetes in the United States. 2011. Available online: http://www.house.gov/degette/diabetes/docs/Costs.fs_Final.pdf (accessed on 1 November 2012).
- UN General Assembly. Political Declaration of the high-level Meeting of The General Assembly on the Prevention and Control of Non-communicable Diseases. 2011. Available online: http://www.un.org/ga/search/view_doc.asp?symbol=A/66/L.1 (accessed on 1 November 2012).
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, Y.; Chen, Y.; Chen, J.; Zhang, W.; Sheng, Q.; Chen, J.; Yu, W.; Nie, Z.; Zhang, Y.; Wu, W.; et al. A Shark Liver Gene-Derived Active Peptide Expressed in the Silkworm, Bombyx mori: Preliminary Studies for Oral Administration of the Recombinant Protein. Mar. Drugs 2013, 11, 1492-1505. https://doi.org/10.3390/md11051492
Liu Y, Chen Y, Chen J, Zhang W, Sheng Q, Chen J, Yu W, Nie Z, Zhang Y, Wu W, et al. A Shark Liver Gene-Derived Active Peptide Expressed in the Silkworm, Bombyx mori: Preliminary Studies for Oral Administration of the Recombinant Protein. Marine Drugs. 2013; 11(5):1492-1505. https://doi.org/10.3390/md11051492
Chicago/Turabian StyleLiu, Yunlong, Ying Chen, Jianqing Chen, Wenping Zhang, Qing Sheng, Jian Chen, Wei Yu, Zuoming Nie, Yaozhou Zhang, Wutong Wu, and et al. 2013. "A Shark Liver Gene-Derived Active Peptide Expressed in the Silkworm, Bombyx mori: Preliminary Studies for Oral Administration of the Recombinant Protein" Marine Drugs 11, no. 5: 1492-1505. https://doi.org/10.3390/md11051492
APA StyleLiu, Y., Chen, Y., Chen, J., Zhang, W., Sheng, Q., Chen, J., Yu, W., Nie, Z., Zhang, Y., Wu, W., Wang, L., Indran, I. R., Li, J., Qian, L., & Lv, Z. (2013). A Shark Liver Gene-Derived Active Peptide Expressed in the Silkworm, Bombyx mori: Preliminary Studies for Oral Administration of the Recombinant Protein. Marine Drugs, 11(5), 1492-1505. https://doi.org/10.3390/md11051492