Identification of Dynamic Changes in Proteins Associated with the Cellular Cytoskeleton after Exposure to Okadaic Acid

Abstract
:1. Introduction
2. Results and Discussion
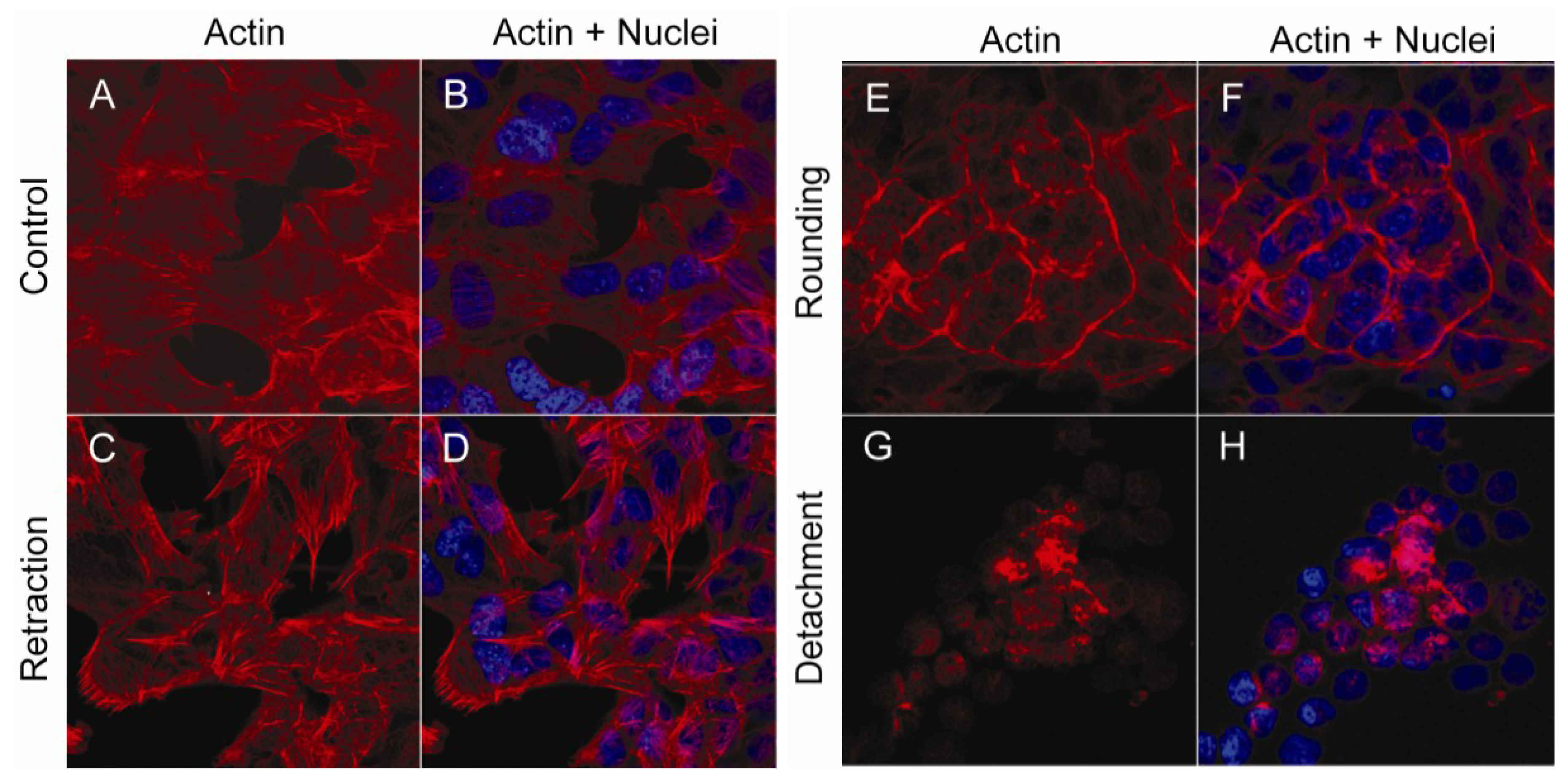
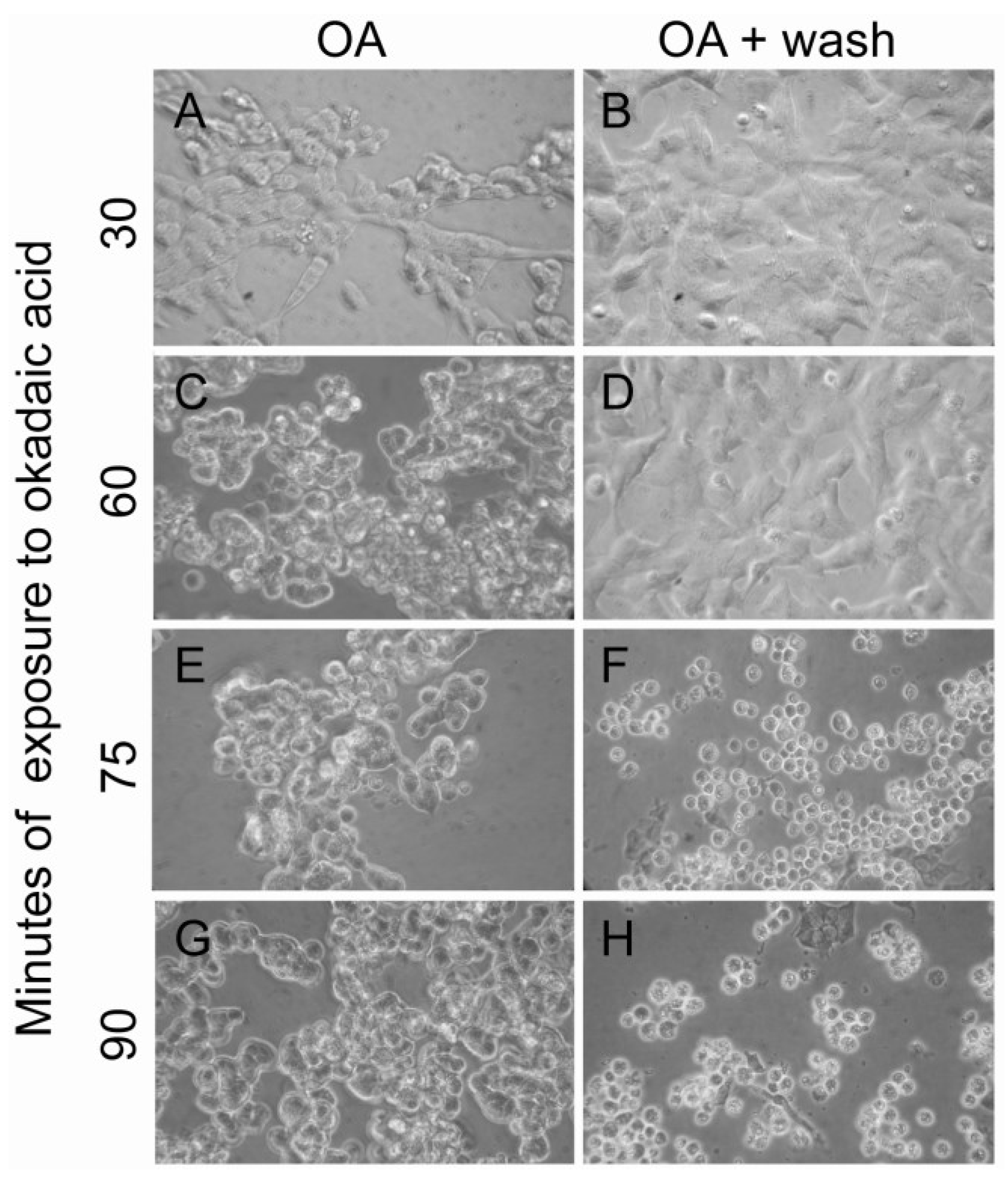
2.1. Actin and Morphological Re-Organization in SH-SY5Y Cells Exposed to Okadaic Acid

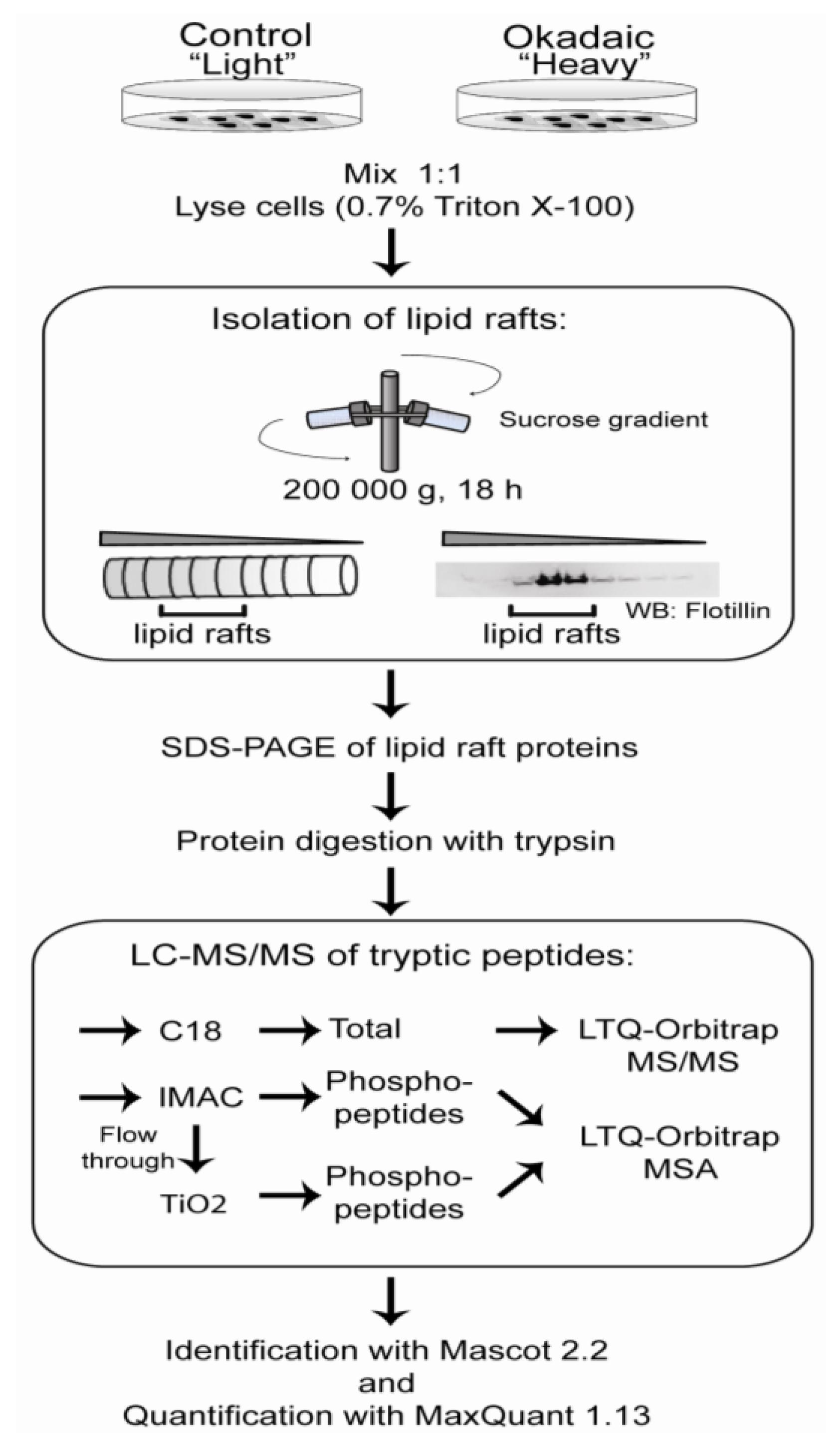
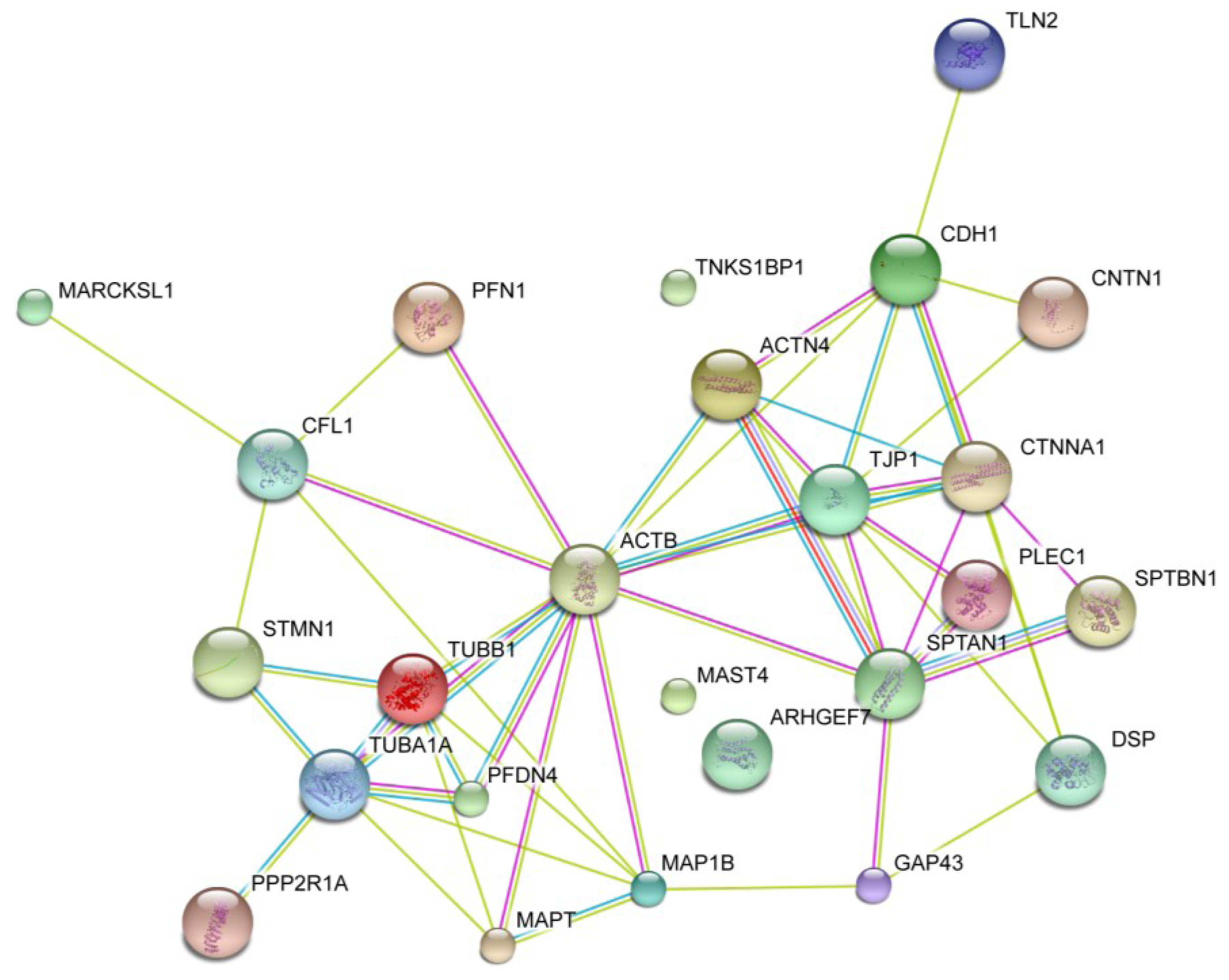
2.2. Enrichment and Identification of Okadaic Acid-Regulated Proteins Associated with the Cytoskeleton


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession a | Protein | Entry name | Phosphopeptide sequence b | P-site | Reference c | Fold change d |
|---|---|---|---|---|---|---|
| Up-regulated | ||||||
| Q9NQP4 | Prefoldin subunit 4 | PFDN4 | _FGS(ph)NINLEADES | S125 | [31] | novel |
| A6NG51 | Putative uncharacterized protein SPTAN1 | SPTAN1 | _S(ph)LQQLAEER | S1217 | [32] | novel |
| P49006 | MARCKS-related protein | MARCKSL1 | _GEVPPKET(ph)PK | S38 | [31] | 1.9 |
| P16949 | Stathmin | STMN1 | _ESVPEFPLS(ph)PPK | S38 | [33] | 1.6 |
| Q15149 | Plectin-1 | PLEC1 | _AQLEPVAS(ph)PAK | S1435 | [32] | 1.6 |
| C9JDD6 | Microtubule-associated protein tau | MAPT | _SPVVSGDT(ph)SPR | T739 | [34] | 1.6 |
| P46821 | Microtubule-associated protein 1B | MAP1B | _VQSLEGEKLS(ph)PK | S1779 | [32] | 1.4 |
| _ASVSPM(ox)DEPVPDSE S(ph)PIEK | S1389 | [35] | 1.4 | |||
| _SDIS(ph)PLTPR_ | S1785 | [32] | 1.5 | |||
| P23528 | Cofilin-1 | CFL1 | _(ac)AS(ph)GVAVSDGVIK | S3 | [32] | 1.3 |
| Q9C0C2 | Tankyrase 1 binding protein 1, 182 kDa | TNKS1BP1 | _VSGAGFS(ph)PSSK | S1138 | [32] | 1.3 |
| _NRS(ph)AEEGELAESK | S1666 | [32] | 1.3 | |||
| Down-regulated | ||||||
| Q01082 | Spectrin beta chain | SPTBN1 | _RPPSPEPS(ph)TK | S2106 | 52.1 | |
| O15021 | Microtubule-associated Ser/Thr-protein kinase 4 | MAST4 | _DCPTLCKQ(de)TDNR Q (de)T(ph)DK | T2357 | 30.0 | |
| Q9Y4G6 | Talin-2 | TLN2 | _LDEGT(ph)PPEPK | T1843 | [32] | 1.5 |
| Q14155 | Rho guanine nucleotide exchange factor 7 | ARHGEF7 | _M(ox)S(ph)GFIYQGK | S518 | [36] | 1.3 |
| Accession a | Protein | Entry name | Fold change b | # Peptides |
|---|---|---|---|---|
| Up-regulated | ||||
| P30153 | Ser/Thr-protein phosphatase 2A regulatory subunit a | PPP2R1A | 23 | 3 |
| A8K0Y4 | Putative uncharacterized protein GAP43 | GAP43 | 2.2 | 9 |
| Q12860 | Contactin-1 | CNTN1 | 1.9 | 5 |
| O43707 | Alpha-actinin-4 | ACTN4 | 1.6 | 2 |
| Down-regulated | ||||
| P07737 | Profilin-1 | PFN1 | 1.3 | 5 |
| P35221 | Catenin alpha-1 | CTNNA1 | 1.3 | 4 |
2.3. Identification of Cytoskeleton-Associated Phosphoproteins Regulated by Okadaic Acid in SH-SY5Y Cells

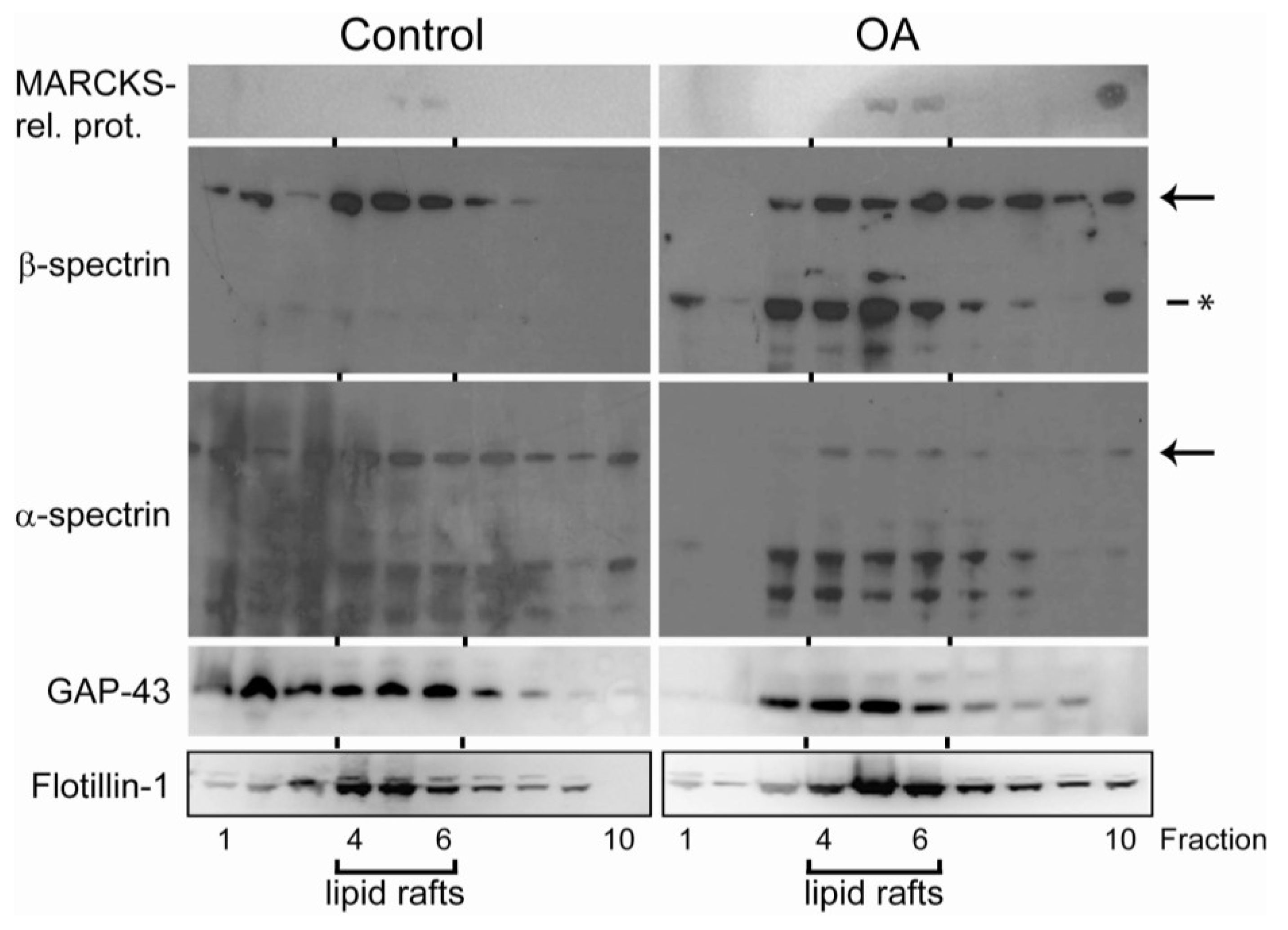
2.4. Translocation of Cytoskeleton-Associated Proteins after Exposure to Okadaic Acid


2.5. Localization of Okadaic Acid-Regulated Phosphoproteins
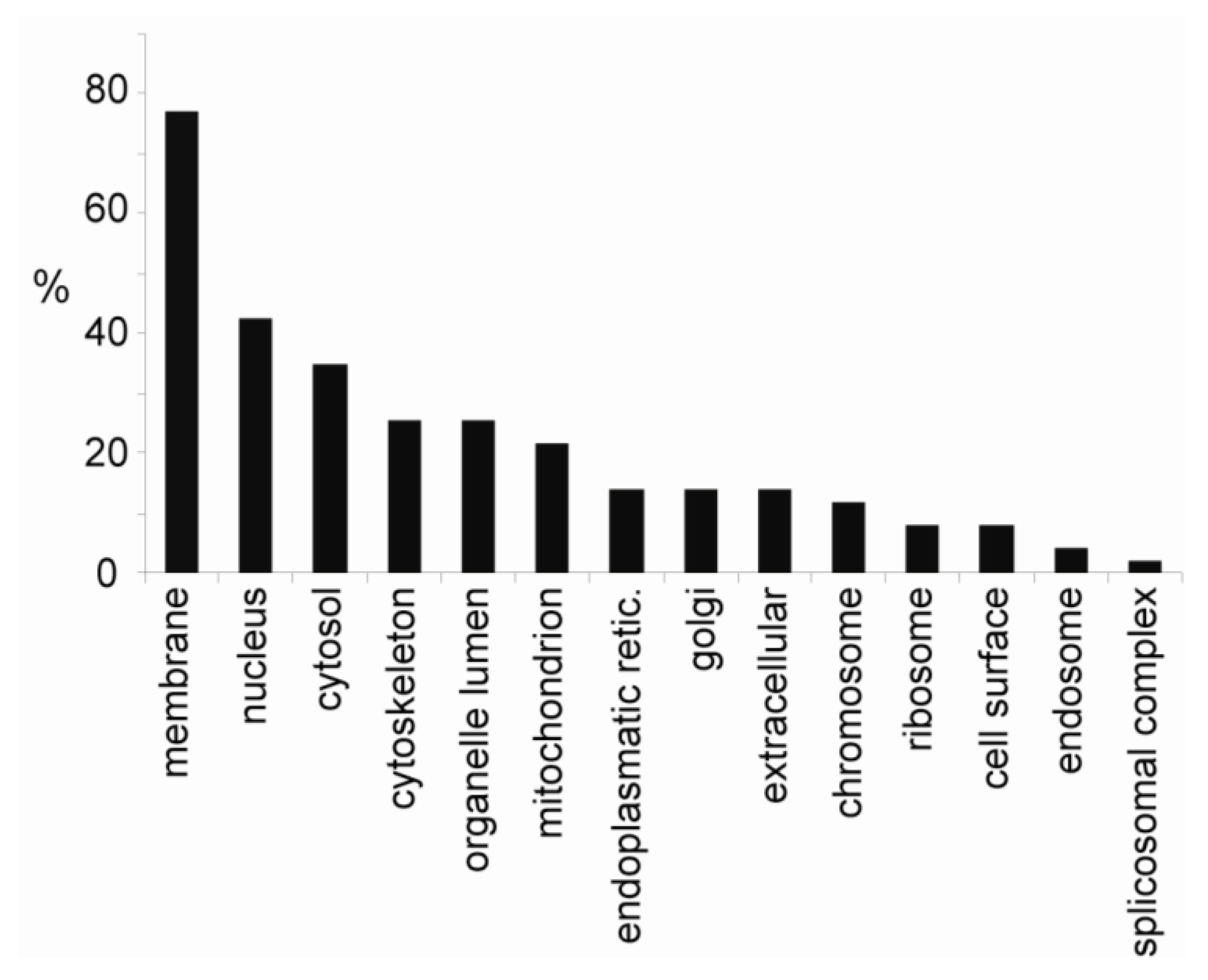
2.6. Okadaic Acid Induces a Coordinated Alternation in Proteins Associated with Cell Cytoskeleton and Cell Adhesion

2.7. Okadaic Acid-Induced Phosphorylation and Relocalization of Cytoskeletal-Associated Proteins May Be General Events of Both Apoptosis-Induction and Cellular Proliferation

3. Experimental Section
3.1. Chemicals and Antibodies
3.2. Cell Culture
3.3. Actin Labeling
3.4. Evaluation of Apoptosis
3.5. Lipid Raft Isolation
3.6. Removal of Lipid and Protein Separation
3.7. Sample Preparation for Mass Spectrometry
3.8. Peptide Separation and Mass Spectrometry Analysis
3.9. Data Processing and Identification
3.10. Western Blotting
3.11. Immunolabeling
3.12. Cell Death Rescue
4. Conclusion
Acknowledgements
Conflict of Interest
References
- Bialojan, C.; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Specificity and kinetics. Biochem. J. 1988, 256, 283–290. [Google Scholar]
- Yoshizawa, S.; Matsushima, R.; Watanabe, M.F.; Harada, K.; Ichihara, A.; Carmichael, W.W.; Fujiki, H. Inhibition of protein phosphatases by microcystins and nodularin associated with hepatotoxicity. J. Cancer Res. Clin. Oncol. 1990, 116, 609–614. [Google Scholar] [CrossRef]
- Dawson, J.F.; Holmes, C.F. Molecular mechanisms underlying inhibition of protein phosphatases by marine toxins. Front. Biosci. 1999, 4, 646–658. [Google Scholar] [CrossRef]
- Fujiki, H.; Suganuma, M. Tumor promotion by inhibitors of protein phosphatases 1 and 2A: The okadaic acid class of compounds. Adv. Cancer Res. 1993, 61, 143–194. [Google Scholar] [CrossRef]
- Suganuma, M.; Fujiki, H.; Suguri, H.; Yoshizawa, S.; Hirota, M.; Nakayasu, M.; Ojika, M.; Wakamatsu, K.; Yamada, K.; Sugimura, T. Okadaic acid: An additional non-phorbol-12-tetradecanoate-13-acetate-type tumor promoter. Proc. Natl. Acad. Sci. USA 1988, 85, 1768–1771. [Google Scholar] [CrossRef]
- Boe, R.; Gjertsen, B.T.; Vintermyr, O.K.; Houge, G.; Lanotte, M.; Doskeland, S.O. The protein phosphatase inhibitor okadaic acid induces morphological changes typical of apoptosis in mammalian cells. Exp. Cell Res. 1991, 195, 237–246. [Google Scholar] [CrossRef]
- Gehringer, M.M. Microcystin-LR and okadaic acid-induced cellular effects: A dualistic response. FEBS Lett. 2004, 557, 1–8. [Google Scholar] [CrossRef]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef]
- Fladmark, K.E.; Serres, M.H.; Larsen, N.L.; Yasumoto, T.; Aune, T.; Doskeland, S.O. Sensitive detection of apoptogenic toxins in suspension cultures of rat and salmon hepatocytes. Toxicon 1998, 36, 1101–1114. [Google Scholar] [CrossRef]
- Fladmark, K.E.; Brustugun, O.T.; Hovland, R.; Boe, R.; Gjertsen, B.T.; Zhivotovsky, B.; Doskeland, S.O. Ultrarapid caspase-3 dependent apoptosis induction by serine/threonine phosphatase inhibitors. Cell Death Differ. 1999, 6, 1099–1108. [Google Scholar]
- Hanana, H.; Talarmin, H.; Pennec, J.P.; Droguet, M.; Morel, J.; Dorange, G. Effect of okadaic acid on cultured clam heart cells: Involvement of MAPkinase pathways. Biol. Open 2012, 1, 1192–1199. [Google Scholar]
- Sontag, J.M.; Sontag, E. Regulation of cell adhesion by PP2A and SV40 small tumor antigen: An important link to cell transformation. Cell Mol. Life Sci. 2006, 63, 2979–2991. [Google Scholar] [CrossRef]
- Yatsunami, J.; Fujiki, H.; Suganuma, M.; Yoshizawa, S.; Eriksson, J.E.; Olson, M.O.; Goldman, R.D. Vimentin is hyperphosphorylated in primary human fibroblasts treated with okadaic acid. Biochem. Biophys. Res. Commun. 1991, 177, 1165–1170. [Google Scholar] [CrossRef]
- Kim, B.; van Golen, C.M.; Feldman, E.L. Degradation and dephosphorylation of focal adhesion kinase during okadaic acid-induced apoptosis in human neuroblastoma cells. Neoplasia 2003, 5, 405–416. [Google Scholar]
- Leira, F.; Vieites, J.M.; Vieytes, M.R.; Botana, L.M. Apoptotic events induced by the phosphatase inhibitor okadaic acid in normal human lung fibroblasts. Toxicol. In Vitro 2001, 15, 199–208. [Google Scholar] [CrossRef]
- Fiorentini, C.; Matarrese, P.; Fattorossi, A.; Donelli, G. Okadaic acid induces changes in the organization of F-actin in intestinal cells. Toxicon 1996, 34, 937–945. [Google Scholar] [CrossRef]
- Vale, C.; Botana, L.M. Marine toxins and the cytoskeleton: Okadaic acid and dinophysistoxins. FEBS J. 2008, 275, 6060–6066. [Google Scholar] [CrossRef]
- Serres, M.; Grangeasse, C.; Haftek, M.; Durocher, Y.; Duclos, B.; Schmitt, D. Hyperphosphorylation of beta-catenin on serine-threonine residues and loss of cell-cell contacts induced by calyculin A and okadaic acid in human epidermal cells. Exp. Cell Res. 1997, 231, 163–172. [Google Scholar]
- Romashko, A.A.; Young, M.R. Protein phosphatase-2A maintains focal adhesion complexes in keratinocytes and the loss of this regulation in squamous cell carcinomas. Clin. Exp. Metastasis 2004, 21, 371–379. [Google Scholar] [CrossRef]
- Pasdar, M.; Li, Z.; Chan, H. Desmosome assembly and disassembly are regulated by reversible protein phosphorylation in cultured epithelial cells. Cell Motil. Cytoskelet. 1995, 30, 108–121. [Google Scholar] [CrossRef]
- Haystead, T.A.; Sim, A.T.; Carling, D.; Honnor, R.C.; Tsukitani, Y.; Cohen, P.; Hardie, D.G. Effects of the tumour promoter okadaic acid on intracellular protein phosphorylation and metabolism. Nature 1989, 337, 78–81. [Google Scholar] [CrossRef]
- Levitan, I.; Gooch, K.J. Lipid rafts in membrane-cytoskeleton interactions and control of cellular biomechanics: Actions of oxLDL. Antioxid. Redox Signal. 2007, 9, 1519–1534. [Google Scholar]
- Viola, A.; Gupta, N. Tether and trap: Regulation of membrane-raft dynamics by actin-binding proteins. Nat. Rev. Immunol. 2007, 7, 889–896. [Google Scholar] [CrossRef]
- Opsahl, J.A.; Hjornevik, L.V.; Bull, V.H.; Fismen, L.; Froyset, A.K.; Gromyko, D.; Solstad, T.; Fladmark, K.E. Increased interaction between DJ-1 and the Mi-2/nucleosome remodelling and deacetylase complex during cellular stress. Proteomics 2010, 10, 1494–1504. [Google Scholar] [CrossRef]
- Arias, C.; Sharma, N.; Davies, P.; Shafit-Zagardo, B. Okadaic acid induces early changes in microtubule-associated protein 2 and tau phosphorylation prior to neurodegeneration in cultured cortical neurons. J. Neurochem. 1993, 61, 673–682. [Google Scholar]
- Nuydens, R.; de Jong, M.; van den Kieboom, G.; Heers, C.; Dispersyn, G.; Cornelissen, F.; Nuyens, R.; Borgers, M.; Geerts, H. Okadaic acid-induced apoptosis in neuronal cells: Evidence for an abortive mitotic attempt. J. Neurochem. 1998, 70, 1124–1133. [Google Scholar]
- Kahns, S.; Lykkebo, S.; Jakobsen, L.D.; Nielsen, M.S.; Jensen, P.H. Caspase-mediated parkin cleavage in apoptotic cell death. J. Biol. Chem. 2002, 277, 15303–15308. [Google Scholar]
- Valdiglesias, V.; Laffon, B.; Pasaro, E.; Mendez, J. Evaluation of okadaic acid-induced genotoxicity in human cells using the micronucleus test and gammaH2AX analysis. J. Toxicol. Environ. Health A 2011, 74, 980–992. [Google Scholar] [CrossRef]
- Zheng, Y.Z.; Foster, L.J. Contributions of quantitative proteomics to understanding membrane microdomains. J. Lipid Res. 2009. [Google Scholar] [CrossRef]
- Chichili, G.R.; Rodgers, W. Cytoskeleton-membrane interactions in membrane raft structure. Cell Mol. Life Sci. 2009, 66, 2319–2328. [Google Scholar] [CrossRef]
- Dephoure, N.; Zhou, C.; Villen, J.; Beausoleil, S.A.; Bakalarski, C.E.; Elledge, S.J.; Gygi, S.P. A quantitative atlas of mitotic phosphorylation. Proc. Natl. Acad. Sci. USA 2008, 105, 10762–10767. [Google Scholar]
- Olsen, J.V.; Blagoev, B.; Gnad, F.; Macek, B.; Kumar, C.; Mortensen, P.; Mann, M. Global, in vivo, and site-specific phosphorylation dynamics in signaling networks. Cell 2006, 127, 635–648. [Google Scholar] [CrossRef]
- Marklund, U.; Larsson, N.; Gradin, H.M.; Brattsand, G.; Gullberg, M. Oncoprotein 18 is a phosphorylation-responsive regulator of microtubule dynamics. EMBO J. 1996, 15, 5290–5298. [Google Scholar]
- Sato, S.; Cerny, R.L.; Buescher, J.L.; Ikezu, T. Tau-tubulin kinase 1 (TTBK1), a neuron-specific tau kinase candidate, is involved in tau phosphorylation and aggregation. J. Neurochem. 2006, 98, 1573–1584. [Google Scholar] [CrossRef]
- Gauci, S.; Helbig, A.O.; Slijper, M.; Krijgsveld, J.; Heck, A.J.; Mohammed, S. Lys-N and trypsin cover complementary parts of the phosphoproteome in a refined SCX-based approach. Anal. Chem. 2009, 81, 4493–4501. [Google Scholar] [CrossRef]
- Mayya, V.; Lundgren, D.H.; Hwang, S.I.; Rezaul, K.; Wu, L.; Eng, J.K.; Rodionov, V.; Han, D.K. Quantitative phosphoproteomic analysis of T cell receptor signaling reveals system-wide modulation of protein-protein interactions. Sci. Signal. 2009, 2, 46. [Google Scholar]
- Mann, M. Functional and quantitative proteomics using SILAC. Nat. Rev. Mol. Cell Biol. 2006, 7, 952–958. [Google Scholar] [CrossRef]
- Bamburg, J.R.; Wiggan, O.P. ADF/cofilin and actin dynamics in disease. Trends Cell Biol. 2002, 12, 598–605. [Google Scholar] [CrossRef]
- Feng, Q.; Baird, D.; Yoo, S.; Antonyak, M.; Cerione, R.A. Phosphorylation of the cool-1/beta-Pix protein serves as a regulatory signal for the migration and invasive activity of Src-transformed cells. J. Biol. Chem. 2010, 285, 18806–18816. [Google Scholar]
- Franco, S.J.; Huttenlocher, A. Regulating cell migration: Calpains make the cut. J. Cell Sci. 2005, 118, 3829–3838. [Google Scholar] [CrossRef]
- Simons, C.T.; Staes, A.; Rommelaere, H.; Ampe, C.; Lewis, S.A.; Cowan, N.J. Selective contribution of eukaryotic prefoldin subunits to actin and tubulin binding. J. Biol. Chem. 2004, 279, 4196–4203. [Google Scholar]
- Steinbock, F.A.; Wiche, G. Plectin: A cytolinker by design. Biol. Chem. 1999, 380, 151–158. [Google Scholar]
- Bordeleau, F.; Galarneau, L.; Gilbert, S.; Loranger, A.; Marceau, N. Keratin 8/18 modulation of protein kinase C-mediated integrin-dependent adhesion and migration of liver epithelial cells. Mol. Biol. Cell 2010, 21, 1698–1713. [Google Scholar] [CrossRef]
- Finlayson, A.E.; Freeman, K.W. A cell motility screen reveals role for MARCKS-related protein in adherens junction formation and tumorigenesis. PLoS One 2009, 4, e7833. [Google Scholar] [CrossRef]
- Berrou, E.; Bryckaert, M. Recruitment of protein phosphatase 2A to dorsal ruffles by platelet-derived growth factor in smooth muscle cells: Dephosphorylation of Hsp27. Exp. Cell Res. 2009, 315, 836–848. [Google Scholar] [CrossRef]
- Ito, A.; Koma, Y.; Sohda, M.; Watabe, K.; Nagano, T.; Misumi, Y.; Nojima, H.; Kitamura, Y. Localization of the PP2A B56gamma regulatory subunit at the Golgi complex: Possible role in vesicle transport and migration. Am. J. Pathol. 2003, 162, 479–489. [Google Scholar] [CrossRef]
- Denny, J.B. Molecular mechanisms, biological actions, and neuropharmacology of the growth-associated protein GAP-43. Curr. Neuropharmacol. 2006, 4, 293–304. [Google Scholar] [CrossRef]
- Honda, K.; Yamada, T.; Endo, R.; Ino, Y.; Gotoh, M.; Tsuda, H.; Yamada, Y.; Chiba, H.; Hirohashi, S. Actinin-4, a novel actin-bundling protein associated with cell motility and cancer invasion. J. Cell Biol. 1998, 140, 1383–1393. [Google Scholar] [CrossRef]
- Shao, H.; Wang, J.H.; Pollak, M.R.; Wells, A. Alpha-actinin-4 is essential for maintaining the spreading, motility and contractility of fibroblasts. PLoS One 2010, 5, e13921. [Google Scholar]
- Su, J.L.; Yang, C.Y.; Shih, J.Y.; Wei, L.H.; Hsieh, C.Y.; Jeng, Y.M.; Wang, M.Y.; Yang, P.C.; Kuo, M.L. Knockdown of contactin-1 expression suppresses invasion and metastasis of lung adenocarcinoma. Cancer Res. 2006, 66, 2553–2561. [Google Scholar] [CrossRef]
- Babakov, V.N.; Petukhova, O.A.; Turoverova, L.V.; Kropacheva, I.V.; Tentler, D.G.; Bolshakova, A.V.; Podolskaya, E.P.; Magnusson, K.E.; Pinaev, G.P. RelA/NF-kappaB transcription factor associates with alpha-actinin-4. Exp. Cell Res. 2008, 314, 1030–1038. [Google Scholar] [CrossRef]
- Shao, H.; Wu, C.; Wells, A. Phosphorylation of alpha-actinin 4 upon epidermal growth factor exposure regulates its interaction with actin. J. Biol. Chem. 2010, 285, 2591–2600. [Google Scholar] [CrossRef]
- Ji, H.; Wang, J.; Nika, H.; Hawke, D.; Keezer, S.; Ge, Q.; Fang, B.; Fang, X.; Fang, D.; Litchfield, D.W.; et al. EGF-induced ERK activation promotes CK2-mediated disassociation of alpha-CATENIN from beta-Catenin and transactivation of beta-Catenin. Mol. Cell 2009, 36, 547–559. [Google Scholar] [CrossRef]
- Condeelis, J. How is actin polymerization nucleated in vivo? Trends Cell Biol. 2001, 11, 288–293. [Google Scholar] [CrossRef]
- Garland, P.; Quraishe, S.; French, P.; O’Connor, V. Expression of the MAST family of serine/threonine kinases. Brain Res. 2008, 1195, 12–19. [Google Scholar] [CrossRef]
- Seimiya, H.; Smith, S. The telomeric poly(ADP-ribose) polymerase, tankyrase 1, contains multiple binding sites for telomeric repeat binding factor 1 (TRF1) and a novel acceptor, 182-kDa tankyrase-binding protein (TAB182). J. Biol. Chem. 2002, 277, 14116–14126. [Google Scholar] [CrossRef]
- Li, D.W.; Xiang, H.; Mao, Y.W.; Wang, J.; Fass, U.; Zhang, X.Y.; Xu, C. Caspase-3 is actively involved in okadaic acid-induced lens epithelial cell apoptosis. Exp. Cell Res. 2001, 266, 279–291. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Fernandez-Tajes, J.; Pasaro, E.; Mendez, J.; Laffon, B. Identification of differentially expressed genes in SHSY5Y cells exposed to okadaic acid by suppression subtractive hybridization. BMC Genomics 2012, 13, 46. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.Y.; Lin, L.; Gao, Y.; Hong, H.S.; Wang, D.Z. Quantitative proteomic analysis of okadaic acid treated mouse small intestines reveals differentially expressed proteins involved in diarrhetic shellfish poisoning. J. Proteomics 2012, 75, 2038–2052. [Google Scholar] [CrossRef]
- Parkin, E.T.; Turner, A.J.; Hooper, N.M. Amyloid precursor protein, although partially detergent-insoluble in mouse cerebral cortex, behaves as an atypical lipid raft protein. Biochem. J. 1999, 344, 23–30. [Google Scholar] [CrossRef]
- Schroeder, M.J.; Shabanowitz, J.; Schwartz, J.C.; Hunt, D.F.; Coon, J.J. A neutral loss activation method for improved phosphopeptide sequence analysis by quadrupole ion trap mass spectrometry. Anal. Chem. 2004, 76, 3590–3598. [Google Scholar] [CrossRef]
- Stensballe, A.; Andersen, S.; Jensen, O.N. Characterization of phosphoproteins from electrophoretic gels by nanoscale Fe(III) affinity chromatography with off-line mass spectrometry analysis. Proteomics 2001, 1, 207–222. [Google Scholar] [CrossRef]
- Larsen, M.R.; Thingholm, T.E.; Jensen, O.N.; Roepstorff, P.; Jørgensen, T.J. Highly selective enrichment of phosphorylated peptides from peptide mixtures using titanium dioxide microcolumns. Mol. Cell. Proteomics 2005, 4, 873–886. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar]
- Swiss Institute of Bioinformatics. Available online: http://expacy.org/sprot/ (accessed on 12 April 2012).
- STRING—Known and Predicted Protein-Protein Interactions. Available online: http://string-db.org/ (accessed on 13 June 2012).
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Opsahl, J.A.; Ljostveit, S.; Solstad, T.; Risa, K.; Roepstorff, P.; Fladmark, K.E. Identification of Dynamic Changes in Proteins Associated with the Cellular Cytoskeleton after Exposure to Okadaic Acid. Mar. Drugs 2013, 11, 1763-1782. https://doi.org/10.3390/md11061763
Opsahl JA, Ljostveit S, Solstad T, Risa K, Roepstorff P, Fladmark KE. Identification of Dynamic Changes in Proteins Associated with the Cellular Cytoskeleton after Exposure to Okadaic Acid. Marine Drugs. 2013; 11(6):1763-1782. https://doi.org/10.3390/md11061763
Chicago/Turabian StyleOpsahl, Jill A., Sonja Ljostveit, Therese Solstad, Kristin Risa, Peter Roepstorff, and Kari E. Fladmark. 2013. "Identification of Dynamic Changes in Proteins Associated with the Cellular Cytoskeleton after Exposure to Okadaic Acid" Marine Drugs 11, no. 6: 1763-1782. https://doi.org/10.3390/md11061763
APA StyleOpsahl, J. A., Ljostveit, S., Solstad, T., Risa, K., Roepstorff, P., & Fladmark, K. E. (2013). Identification of Dynamic Changes in Proteins Associated with the Cellular Cytoskeleton after Exposure to Okadaic Acid. Marine Drugs, 11(6), 1763-1782. https://doi.org/10.3390/md11061763