Evaluation of Anti-Nociceptive and Anti-Inflammatory Activities of a Heterofucan from Dictyota menstrualis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
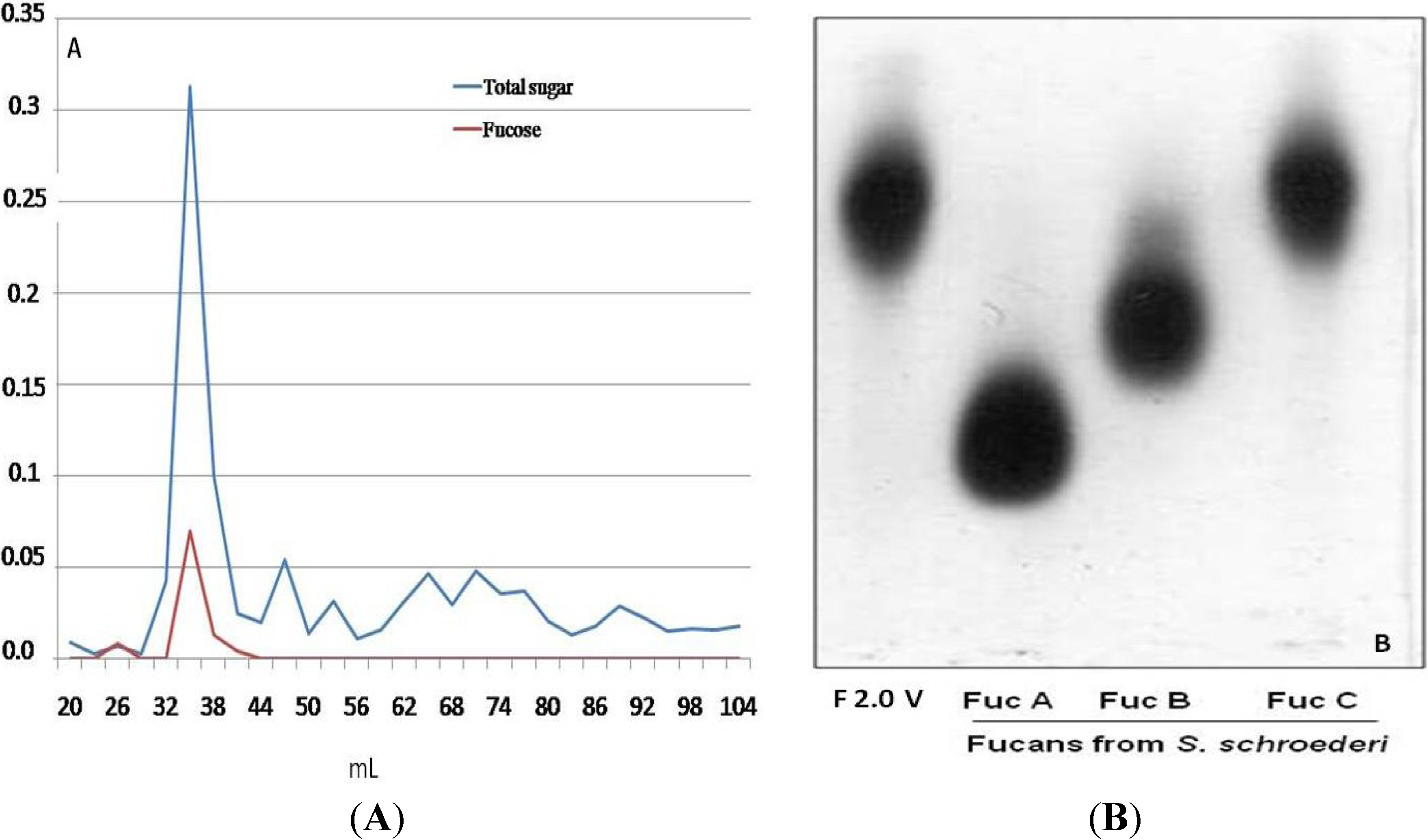
2.1. Obtaining Fucan F2.0v from D. menstrualis

2.2. Infrared Analysis of F2.0v

2.3. Anticoagulant Activity
2.4. Anti-Inflammatory Activity
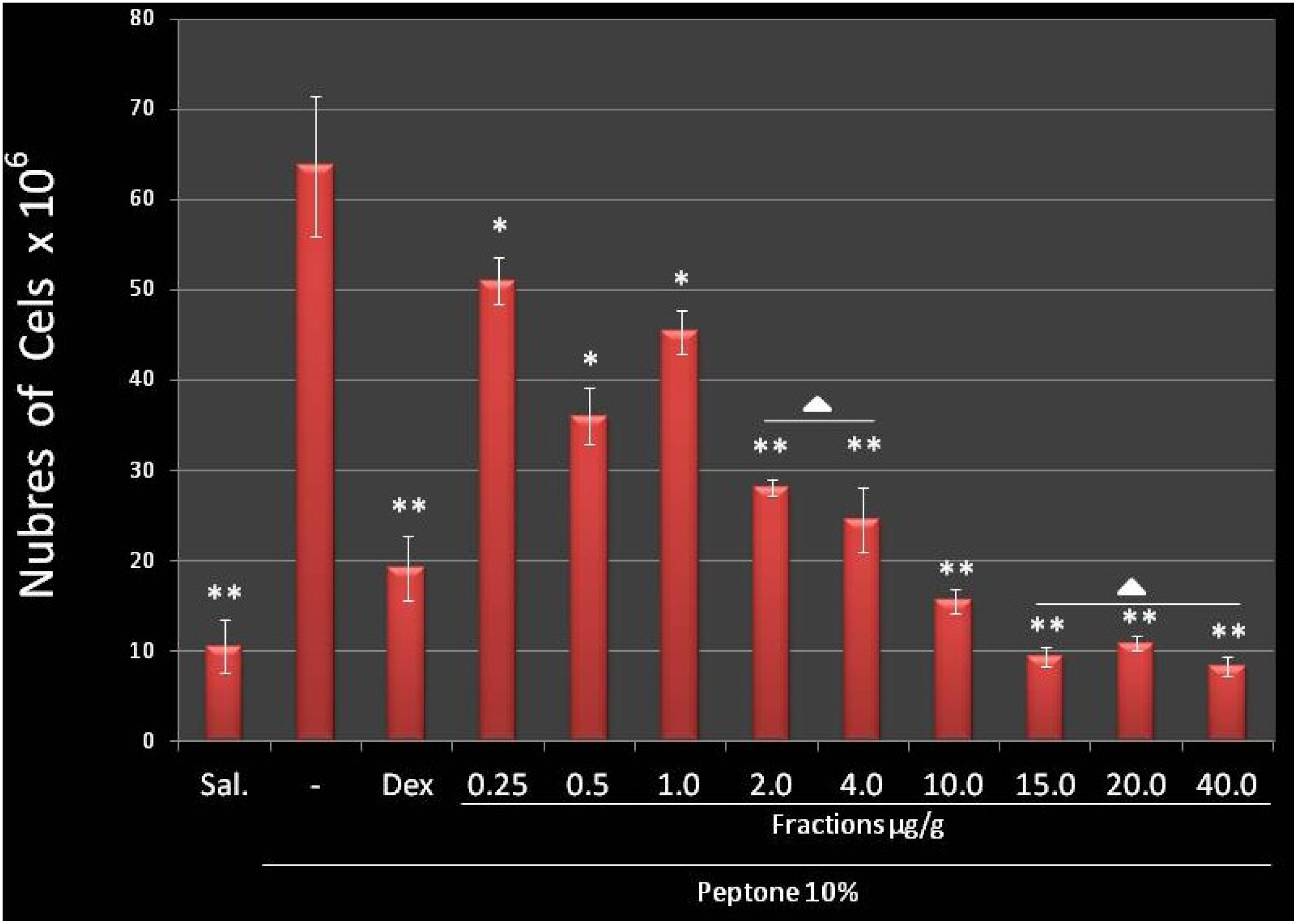
2.4.1. Fuc C Inhibits Leukocyte Migration into the Peritoneal Cavity
2.4.2. F2.0v Does Not Influence the Production of Pro-Inflammatory Cytokines

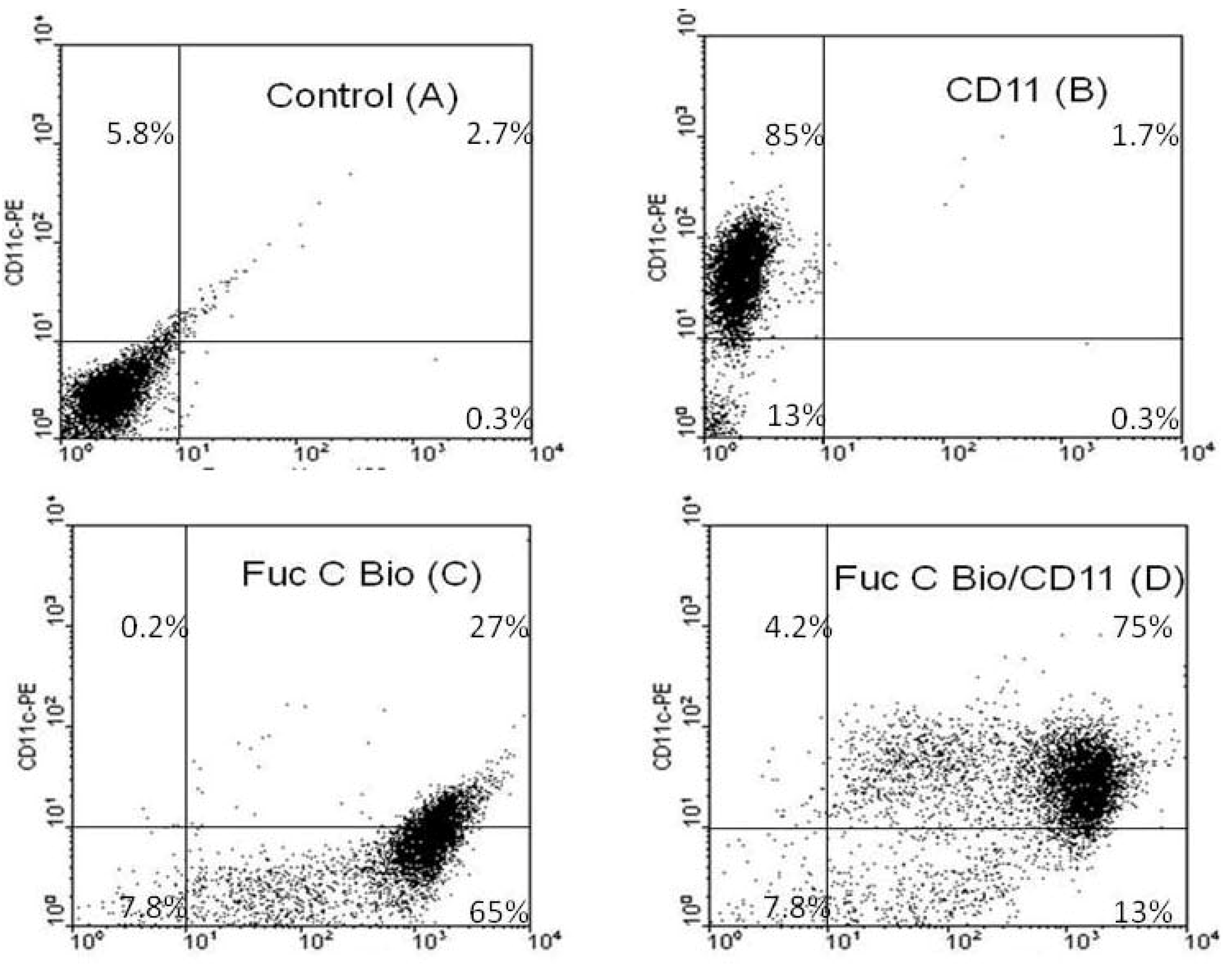
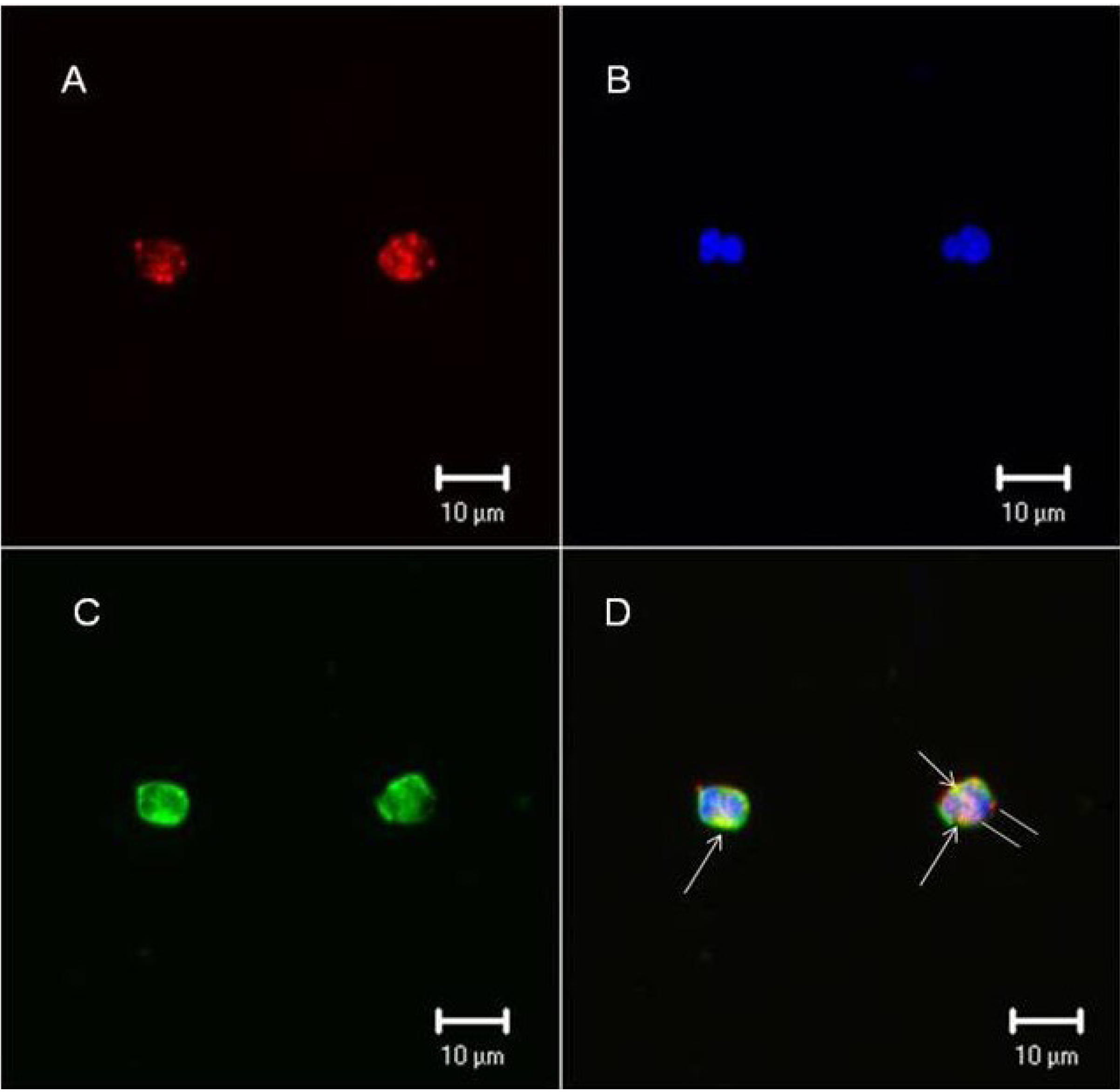
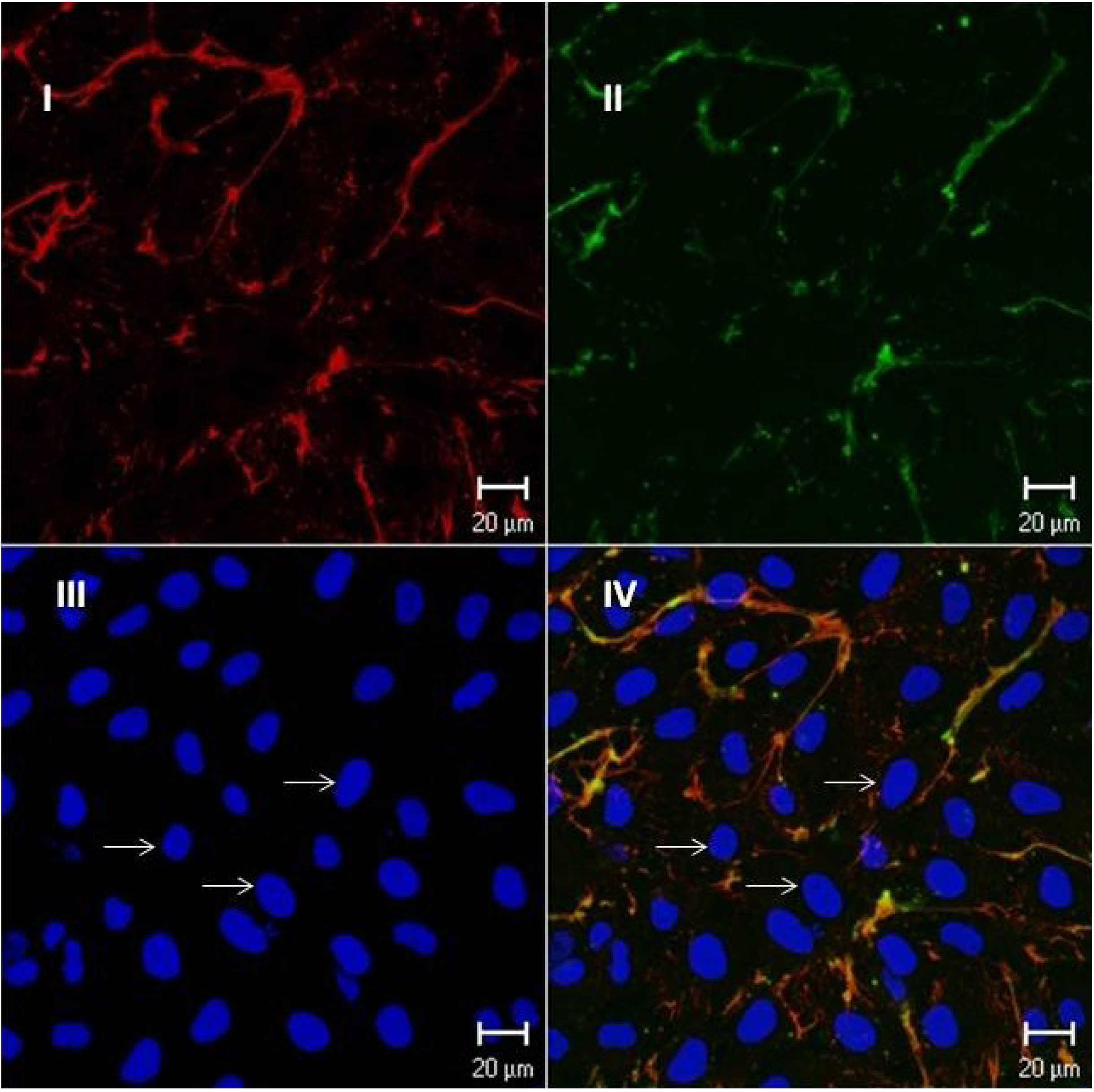
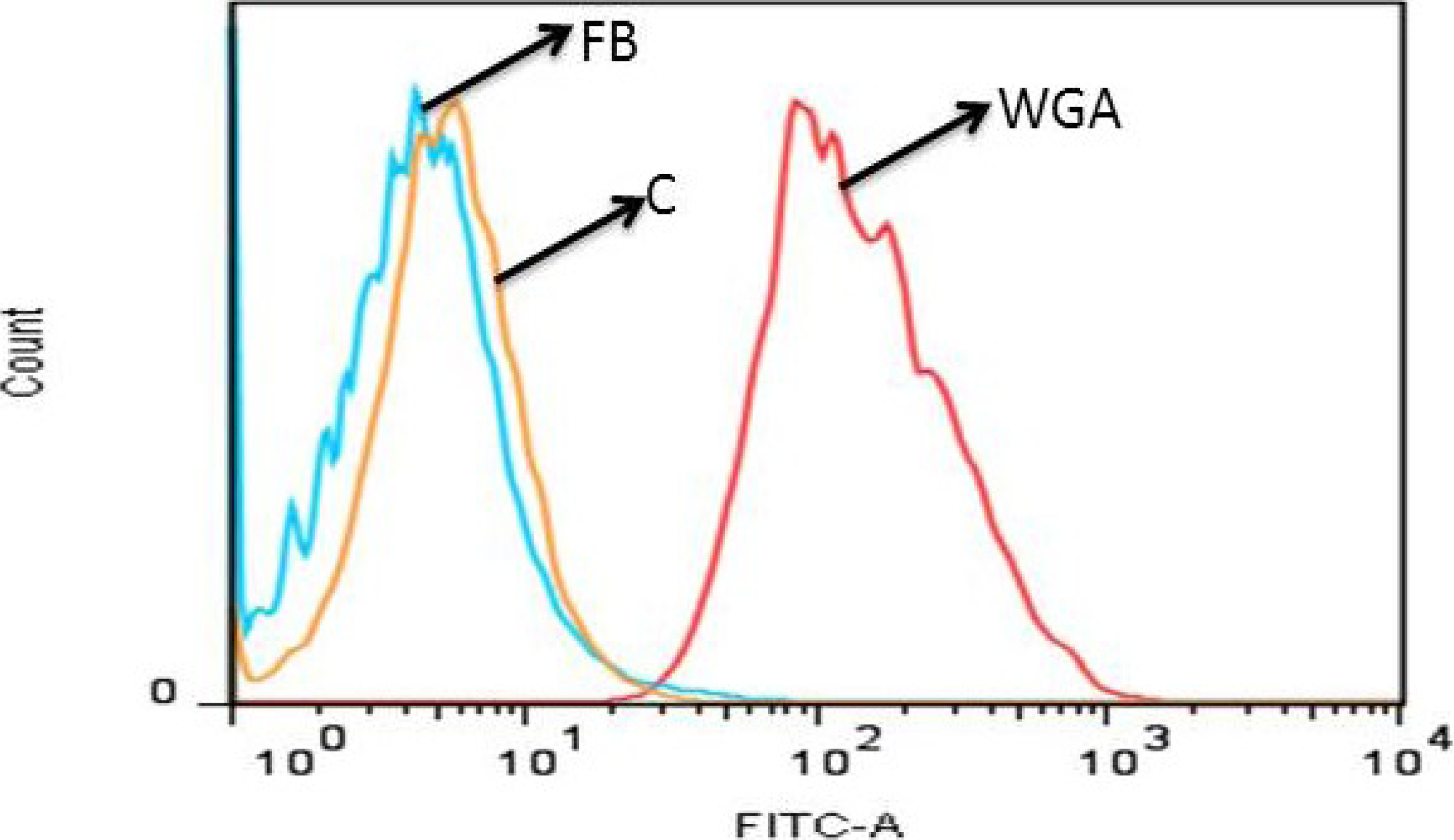
2.4.3. Fuc C Binds to Murine Leukocyte Surface




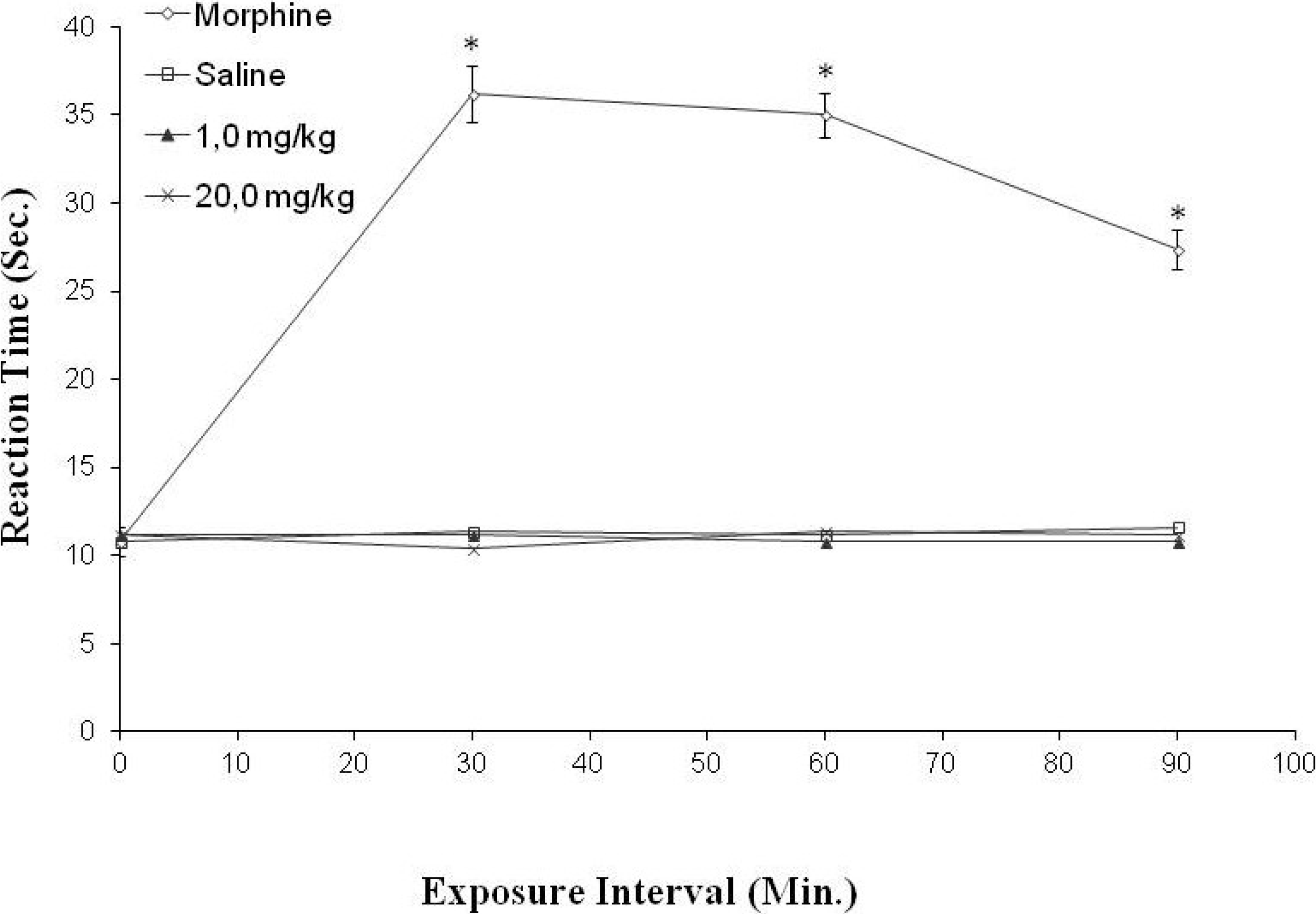
2.5. Anti-Nociceptive Activity


3. Experimental Section
3.1. F2.0v Purifying
3.2. Agarose Gel Electrophoresis
3.3. Chemical Analysis and Monosaccharide Composition
3.4. Fourier Transformed Infrared Spectroscopy (FTIR)
3.5. Anticoagulant Activity
3.6. Animals and Cells
3.7. Migration of Leukocytes into the Peritoneal Cavity
3.8. Quantification of Cytokines
3.9. Biotinylation of F2.0v
3.9.1. Flow Cytometric Analysis
3.9.2. Immunocytochemistry
3.9.3. Acetic Acid-Induced Writhing Test
3.9.4. Hot Plate Test
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Le Bars, D.; Gozariu, M.; Cardden, S.W. Animal models of nocicepcion. Phamacol. Rev. 2001, 53, 597–652. [Google Scholar]
- Michel, G.; Tonon, T.; Scornet, D.; Cock, J.M.; Kloareg, B. The cell wall polysaccharide metabolism of the brown alga Ectocarpus siliculosus. Insights into the evolution of extracellular matrix polysaccharides in Eukaryotes. New Phytol. 2010, 188, 82–97. [Google Scholar] [CrossRef]
- Camara, R.B.G.; Costa, L.S.; Fidelis, G.P.; Nobre, L.T.D.B.; Dantas-Santos, N.; Cordeiro, S.L.; Costa, M.S.S.P.; Alves, L.G.; Rocha, H.A.O. Heterofucans from the brown seaweed Canistrocarpus cervicornis with anticoagulant and antioxidant activities. Mar. Drugs 2011, 9, 124–138. [Google Scholar] [CrossRef]
- Magalhães, K.D.; Costa, L.S.; Fidelis, G.P.; Oliveira, R.M.; Nobre, L.T.; Dantas-Santos, N.; Camara, R.B.; Albuquerque, I.R.; Cordeiro, S.L.; Sabry, D.A.; et al. Anticoagulant, antioxidant and antitumor activities of heterofucans from the seaweed Dictyopteris delicatula. Int. J. Mol. Sci. 2011, 12, 3352–3365. [Google Scholar] [CrossRef]
- Costa, L.S.; Fidelis, G.P.; Telles, C.B.S.; Dantas-Santos, N.; Camara, R.B.G.; Cordeiro, S.L.; Costa, M.S.S.P.; Almeida-Lima, J.; Melo-Silveira, R.F.; Albuquerque, I.R.L.; et al. Antioxidant and antiproliferative activities of heterofucans from the seaweed Sargassum filipendula. Mar. Drugs 2011, 9, 952–966. [Google Scholar] [CrossRef]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef]
- Thompson, K.D.; Dragar, C. Antiviral activity of Undaria pinnatifida against herpes simplex virus. Phytother. Res. 2004, 18, 551–555. [Google Scholar] [CrossRef]
- Ponce, N.M.A.; Pujol, C.A.; Damonte, E.B.; Flores, M.L.; Stortz, C.A. Fucoidans from the brown seaweed Adenocystis utricularis: Extraction methods, antiviral activity and structural studies. Carbohydr. Res. 2003, 338, 153–165. [Google Scholar] [CrossRef]
- Paiva, A.A.; Castro, A.J.; Nascimento, M.S.; Will, L.S.; Santos, N.D.; Araújo, R.M.; Xavier, C.A.; Rocha, F.A.; Leite, E.L. Antioxidant and anti-inflammatory effect of polysaccharides from Lobophora variegata on zymosan-induced arthritis in rats. Int. Immunopharmacol. 2011, 11, 1241–1250. [Google Scholar] [CrossRef]
- Sokolova, R.V.; Ermakova, S.P.; Awada, S.M.; Zvyagintseva, T.N.; Kanaan, H.M. Composition, structural characteristics, and antitumor properties of polysaccharides from the brown algae Dictyopteris polypodioides and Sargassum sp. Chem. Nat. Comp. 2011, 47, 329–334. [Google Scholar] [CrossRef]
- Rocha, H.A.O.; Moraes, F.A.; Trindade, E.S.; Franco, C.R.C.; Torquato, R.J.S.; Veiga, S.S.; Valente, A.P.; Mourão, P.A.; Leite, E.L.; Nader, H.B.; et al. Structural and haemostatic activities of a sulfated galactofucan from the brown alga Spatoglossum schroederi. An ideal antithrombotic agent? J. Biol. Chem. 2005, 280, 41278–41288. [Google Scholar]
- Almeida-Lima, J.; Costa, L.S.; Silva, N.B.; Melo-Silveira, R.F.; Silva, F.V.; Felipe, M.B.M.C.; Medeiros, S.R.B.M.; Leite, E.L.; Rocha, H.A.O. Evaluating the possible genotoxic, mutagenic and tumor cell proliferation-inhibition effects of a non-anticoagulant, but antithrombotic algal heterofucan. J. Appl. Toxicol. 2010, 30, 708–715. [Google Scholar] [CrossRef]
- Rocha, H.A.O.; Franco, C.R.C.; Trindade, E.S.; Veiga, S.S.; Leite, E.L.; Dietrich, C.P.; Nader, H.B. Fucan inhibits Chinese hamster ovary cell (CHO) adhesion to fibronectin by binding to the extracellular matrix. Planta Med. 2005, 71, 628–633. [Google Scholar] [CrossRef]
- Almeida-Lima, J.; Dantas-Santos, N.; Gomes, D.L.; Cordeiro, S.L.; Sabry, D.A.; Costa, L.S.; Freitas, M.L.; Silva, N.B.; Moura, C.E.B.; Lemos, T.M.A.M.; et al. Evaluation of acute and subchronic toxicity of a non-anticoagulant, but antithrombotic algal heterofucan from the Spatoglossum schröederi in wistar rats. Rev. Bras. Farmacogn. 2011, 21, 674–679. [Google Scholar] [CrossRef]
- Farias, W.R.; Lima, P.C.; Rodrigues, N.V.; Siqueira, R.C.; Amorim, R.M.; Pereira, M.G.; Assreuy, A.M. A novel antinociceptive sulphated polysaccharide of the brown marine alga Spatoglossum schroederi. Nat. Prod. Commun. 2011, 6, 863–866. [Google Scholar]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A; Câmara, R.B.G.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida-Lima, J.; Farias, E.H.C.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Albuquerque, I.R.L.; Queiroz, K.C.S.; Alves, L.G.; Santos, E.A.; Leite, E.L.; Rocha, H.A.O. Heterofucans from Dictyota menstrualis have anticoagulant activity. Braz. J. Med. Biol. Res. 2004, 37, 167–171. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars, and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Dische, Z. Color Reactions of 6-Deoxi and 3-6-Dideoxioses. In Methods in Carbohydrate Chemistry; Whistler, R.L., Wolfrom, M.L., Eds.; Academic Press, Inc.: New York, NY, USA, 1962; Volume 1, pp. 501–503. [Google Scholar]
- Rocha, H.A.O.; Bezerra, L.C.; Albuquerque, I.R.L.; Costa, L.S.; Guerra, C.M.; Abreu, L.D.; Nader, H.B.; Leite, E.L. A xylogalactofucan from the brown seaweed Spatoglossum schroederi stimulates the synthesis of an antithrombotic heparan sulfate from endothelial. Planta Med. 2005, 71, 379–381. [Google Scholar] [CrossRef]
- Laurie-Eve, R.; Sylvie, L.T.; Martin, B. Structural characterization of laminaran and galactofucan extracted from the brown seaweed Saccharina longicruris. Phytochemistry 2010, 71, 1586–1595. [Google Scholar] [CrossRef]
- Ermakova, S.; Sokolova, R.; Kim, S.; Um, B.; Isakov, V.; Zvyagintseva, T. Fucoidans from brown seaweeds Sargassum hornery, Eclonia cava, Costaria costata: Structural characteristics and anticancer activity. Appl. Biochem. Biotechnol. 2011, 164, 841–850. [Google Scholar] [CrossRef]
- Barroso, E.M.; Costa, L.S.; Medeiros, V.P.; Cordeiro, S.L.; Costa, M.S.P.; Franco, C.R.; Nader, H.B.; Leite, E.L.; Rocha, H.A.O. A non-anticoagulant heterofucan has antithrombotic activity in vivo. Planta Med. 2008, 74, 708–712. [Google Scholar]
- Wang, X.; Wang, J.; Zhang, J.; Zhao, B.; Yao, J.; Wang, Y. Structure-antioxidant relashionships of sulfated galactomannan from guar gum. Int. J. Biol. Macromol. 2010, 46, 59–66. [Google Scholar] [CrossRef]
- Shumbula, P.M.; Moloto, M.J.; Tshikhundo, T.R.; Fernandes, M. Dichloro(bis[diphenylthiourea]) cadmium complex as a precursor for HDA-capped CdS nanoparticles and their solubility in water. S. Afr. J. Sci. 2010, 106, 1–7. [Google Scholar]
- Xu, X.; Li, S.; Jia, F.; Liu, P. Synthesis and antimicrobial activity of nano-fumed sílica derivative with N,N-dimethyl-N-hexadecylamine. Life Sci. J. 2006, 3, 59–62. [Google Scholar]
- Brito, A.S.; Arimateia, D.S.; Souza, L.R.; Lima, M.A.; Santos, V.O.; Medeiros, V.P.; Ferreira, P.A.; Silva, R.A.; Ferreira, C.V.; Justo, G.Z.; et al. Anti-inflammatory properties of a heparin-like glycosaminoglycan with reduced anticoagulant activity isolated from a marine shrimp. Bioorg. Med. Chem. 2008, 16, 9588–9595. [Google Scholar] [CrossRef]
- Lima, G.M.; Quintans-Júnior, L.J.; Thomazzi, S.M.; Almeida, E.M.S.A.; Melo, M.S.; Serafi, M.R.; Cavalcanti, S.C.H.; Gelain, D.P.; Santos, J.P.A.; Blank, A.F.; et al. Phytochemical screening, antinociceptive and anti-infl ammatory activities of Chrysopogon zizanioides essential oil. Rev. Bras. Farmacogn. 2012, 22, 443–450. [Google Scholar] [CrossRef]
- Anastase-Ravion, S.; Carreno, M.P.; Blondin, C.; Ravion, O.; Champion, J.; Chaubet, F.; Haeffner-Cavaillon, N.; Letourneur, D. Heparin-like polymers modulate proinflammtory cytokine production by lipopolysaccharide-stimulated human monocytes. J. Biomed. Mater. Res. 2002, 60, 375–383. [Google Scholar] [CrossRef]
- Coura, C.O.; de Araújo, I.W.; Vanderlei, E.S.; Rodrigues, J.A.; Quinderé, A.L.; Fontes, B.P.; de Queiroz, I.N.; de Menezes, D.B.; Bezerra, M.M.; e Silva, A.A.; et al. Antinociceptive and anti-inflammatory activities of sulphated polysaccharides from the red seaweed Gracilaria cornea. Basic Clin. Pharmacol. Toxicol. 2012, 110, 335–341. [Google Scholar]
- Assreuy, A.M.S.; Gomes, D.M.; Silva, M.S.J.; Torres, V.M.; Siqueira, R.C.L.; Pires, A.F.; Criddle, D.N.; Alencar, N.M.N.; Cavada, B.S.; Sampaio, A.H.; et al. Biological effects of a sulfated-polysaccharide isolated from the marine red algae Champia feldmannii. Biol. Pharm. Bull. 2008, 31, 691–695. [Google Scholar]
- Ribeiro, R.A.; Vale, M.L.; Thomazzi, S.M.; Paschoalato, A.B.; Poole, S.; Ferreira, S.H.; Cunha, F.Q. Involvement of resident macrophages and mast cells in the writhing nociceptive response induced by zymosan and acetic acid in mice. Eur. J. Pharmacol. 2000, 387, 111–118. [Google Scholar] [CrossRef]
- Dietrich, C.P.; Dietrich, S.M.C. Electrophoretic behaviour of acidic mucopolysaccarides in diamine buffers. Anal. Biochem. 1976, 70, 645–647. [Google Scholar] [CrossRef]
- Dodgson, K.S.; Price, R.G. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar]
- Spector, J. Refinemente of the comassie blue method of protein quantification. A simple and liner spectrofotometric assay of 0.5 to 50 μg of protein. Anal. Biochem. 1978, 86, 142–146. [Google Scholar] [CrossRef]
- Costa, M.S.S.P.; Costa, L.S.; Cordeiro, S.L.; Almeida-Lima, J.; Dantas-Santos, N.; Magalhães, K.D.; Sabry, D.A.; Albuquerque, I.R.L.; Pereira, M.R.; Leite, E.L.; Rocha, H.A.O. Evaluating the possible anticoagulant and antioxidante effects of sulfated polysaccharides from the tropical green alga Caulerpa cupressoides var. flabella. J. Appl. Phycol. 2012, 24, 1159–1167. [Google Scholar] [CrossRef]
- Eddy, N.B.; Leimbach, P. Synthetic analgesics; dithienylbutenyland dithienylbutylamines. J. Pharmacol. Exp. Ther. 1953, 107, 385–389. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Albuquerque, I.R.L.; Cordeiro, S.L.; Gomes, D.L.; Dreyfuss, J.L.; Filgueira, L.G.A.; Leite, E.L.; Nader, H.B.; Rocha, H.A.O. Evaluation of Anti-Nociceptive and Anti-Inflammatory Activities of a Heterofucan from Dictyota menstrualis. Mar. Drugs 2013, 11, 2722-2740. https://doi.org/10.3390/md11082722
Albuquerque IRL, Cordeiro SL, Gomes DL, Dreyfuss JL, Filgueira LGA, Leite EL, Nader HB, Rocha HAO. Evaluation of Anti-Nociceptive and Anti-Inflammatory Activities of a Heterofucan from Dictyota menstrualis. Marine Drugs. 2013; 11(8):2722-2740. https://doi.org/10.3390/md11082722
Chicago/Turabian StyleAlbuquerque, Ivan Rui Lopes, Sara Lima Cordeiro, Dayanne Lopes Gomes, Juliana Luporini Dreyfuss, Luciana Guimarães Alves Filgueira, Edda Lisboa Leite, Helena Bonciani Nader, and Hugo Alexandre Oliveira Rocha. 2013. "Evaluation of Anti-Nociceptive and Anti-Inflammatory Activities of a Heterofucan from Dictyota menstrualis" Marine Drugs 11, no. 8: 2722-2740. https://doi.org/10.3390/md11082722
APA StyleAlbuquerque, I. R. L., Cordeiro, S. L., Gomes, D. L., Dreyfuss, J. L., Filgueira, L. G. A., Leite, E. L., Nader, H. B., & Rocha, H. A. O. (2013). Evaluation of Anti-Nociceptive and Anti-Inflammatory Activities of a Heterofucan from Dictyota menstrualis. Marine Drugs, 11(8), 2722-2740. https://doi.org/10.3390/md11082722